










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkCuadernos de herpetología
versión On-line ISSN 1852-5768
Cuad. herpetol. vol.27 no.2 San Salvador de Jujuy set. 2013
TRABAJOS
Taxonomía y filogenia de un grupo de lagartos amenazados: el grupo de Liolaemus anomalus (Iguania: Liolaemidae)
Cristian Simón Abdala1, 2, Viviana Isabel Juárez Heredia2
1 CONICET - Instituto de Herpetología, Fundación Miguel Lillo. Miguel Lillo 251. 4000. Tucumán. Argentina.
2 Facultad de Ciencias Naturales e I.M.L., Universidad Nacional de Tucumán. Miguel Lillo 251. 4000. Tucumán. Argentina.
Recibido: 17/04/12
Revisado: 10/10/12
Aceptado: 27/01/13
RESUMEN
El género Liolaemus, es uno de los grupos naturales con mayor cantidad de especies dentro de Iguania, está formado por dos subgéneros y varios grupos monofiléticos. Uno de estos, es el grupo monofilético de Liolaemus anomalus, uno de los clados con mayor cantidad de arreglos taxonómicos propuestos. Este conjunto está representado por un escaso número de especies las cuales tienen baja densidad poblacional. Se distribuye desde el centro sur de Catamarca hasta el suroeste de La Pampa, Argentina. Las especies que integran este clado están asociadas a ambientes áridos y salobres, con escasa vegetación y expuestas a elevadas temperaturas, siendo lagartijas ovíparas e insectívoras y sin dimorfismo sexual. Son taxa muy difíciles de observar, varias especies y poblaciones están amenazadas debido principalmente a factores antrópicos.
En este trabajo, se realiza una revisión taxonómica y un análisis filogenético del grupo de Liolaemus anomalus. En la matriz morfológica utilizada para los análisis filogenéticos, se describen nuevos caracteres informativos para el clado y el género, que ayudan a clarificar las relaciones internas del grupo. Como apoyo a la metodología cladística, se realizaron análisis estadísticos (MANOVA), con el fin de efectuar un estudio detallado de los caracteres morfométricos. Por otro lado, se realiza una redescripción formal de Liolaemus anomalus, Liolaemus ditadai, Liolaemus lentus y Liolaemus pseudoanomalus, con base en nuevos ejemplares colectados en diversas campañas y utilizando caracteres no analizados en las descripciones originales. Además se describen tres nuevas especies para la ciencia, se aportan datos que actualizan el conocimiento de las diferentes especies que integran el grupo y se analizan las congruencias con las hipótesis filogenéticas presentadas por otros autores. Los resultados filogenéticos obtenidos aportan evidencia que justifican la descripción de estas tres nuevas especies y se propone una nueva hipótesis de parentesco para el grupo de Liolaemus anomalus.
PALABRAS CLAVE: Lagartijas; Liolaemus; Cladistica; Estadistica, Morfología; Nuevas Especies; Argentina.
ABSTRACT
The genus Liolaemus is one of the natural groups with the greatest number of species within Iguania, formed by two subgenera and several monophyletic groups. One of these is the monophyletic group of Liolaemus anomalus, a clade with perhaps the greatest number of proposed taxonomic arrangements. This group is represented by a small number of species with low population densities. It is distributed from south-central Catamarca to southwest La Pampa provinces, Argentina. Species included in this clade are associated to arid and brackish environments, with scarce vegetation, and exposed to high temperatures. They are oviparous, insectivorous and don´t exhibit sexual dimorphism. These species are very difficult to observe and some are being threatened mainly due to human factors. Here, we present a taxonomic review and phylogenetic analysis of the Liolaemus anomalus group. In the morphological matrix used in the phylogenetic analysis, we describe new informative characters for the clade and the genus, which help clarify the relationships within the group. Supporting the cladistic method, statistical analyses (MANOVA) were performed, in order to study in more detail the morphometric characters. In addition, a formal redescription of Liolaemus anomalus, Liolaemus ditadai, Liolaemus lentus and Liolaemus pseudoanomalus, is performed based on new specimens collected during last field trips, using characters not included in the original description. We also describe three new species, providing data which update our knowledge of the different species of the group. Congruence with previous phylogenetic hypotheses is discussed.
The phylogenetic results we obtained provide evidence which justify the description of three new species and the proposal of a new hypothesis of relationships for the group of Liolaemus anomalus.
KEYWORDS: Lizards; Liolaemus; Cladistics; Statistics; Morphology; New Species; Argentina.
INTRODUCCIÓN
El género Liolaemus, pertenece a la familia Liolaemidae (Frost y Etheridge, 1989) y es un grupo natural de lagartijas que presenta una amplia distribución latitudinal y altitudinal (Etheridge, 1995). Presenta una gran diversidad de formas que incluye especies de hábitos saxícolas, arborícolas, samófilos, con modalidad reproductiva ovípara o vivípara, así como insectívoros, herbívoros y omnívoros. La variedad de patrones de coloración, tamaños y formas vuelve aún más atractivo al género.
El conocimiento del género Liolaemus en los últimos 10 años se ha incrementado notablemente debido al desarrollo de estudios anatómicos, biogeográficos, ecológicos, etológicos, filogenéticos, moleculares y taxonómicos, transformándose en un género de "actualidad" en el ambiente herpetológico. Estos trabajos han llevado a que Liolaemus modificara su numero de especies, de 160 en el año 2000 (Etheridge y Espinoza, 2000) a más de 240 en principios del 2012 (Abdala et al., 2012; Avila et al., 2013).
El género Liolaemus está formado por numerosos grupos y clados entre los cuales se destaca el grupo monofilético de Liolaemus anomalus (Abdala, 2007a). Es uno de los grupos de Liolaemus con mayor cantidad de arreglos taxonómicos (Müller, 1928; Donoso Barros, 1971; Cei, 1971, 1981, 1986,1993; Nuñez y Yañez, 1984; Laurent, 1984; Etheridge 1995; Abdala, 2007a). Actualmente el grupo estaría formado por 4 especies reconocidas: L. anomalus, L. pseudoanomalus, L. ditadai y L. lentus, y tres que se describen en este trabajo.
El grupo de Liolaemus anomalus, está formado por especies de lagartijas de las cuales se conoce muy poco sobre su biología y hábitos en general (Cei, 1986; 1993; Abdala, 2007a). La mayoría de las especies del grupo presentan un endemismo marcado (Abdala, 2007a; Abdala et al., 2012). A diferencia de la mayoría de los Liolaemus, las especies del grupo de L. anomalus presentan poblaciones muy escasas, tornándose muy difícil encontrarlos o avistarlos (Cei, 1986; Abdala, 2007a). A su vez la mayoría de estas especies están asociadas a ambientes salobres con escasa vegetación, algunos antropizados y expuestas a elevadas temperaturas (Cei, 1986; 1993; Abdala, 2007a).
Estas lagartijas tienden a utilizar cuevas abandonadas, arbustos o costrones de sal y arena para ocultarse (Cei, 1986, 1993; Abdala, 2007a). Suelen ser muy crípticos respecto al sustrato, lo que aumenta la dificultad en su avistaje. Son lagartijas ovíparas e insectívoras y no tienen dimorfismo sexual (Abdala, 2007a). Debido a que presentan poblaciones con densidad poblacional muy baja con respecto a la mayoría de los Liolaemus, se conoce muy poco sobre su actividad diaria y estacional. Se han postulado varias hipótesis, una de las cuales propone que son lagartijas crepusculares (Cei, 1986), sin embargo hemos encontrado algunos ejemplares de L. anomalus y una de las especies que se describe en este trabajo en horas de la siesta bajo temperaturas de más de 35°. Abdala (2007a) menciona que en algunas especies la actividad diaria en verano es unimodal mientras que en otras es bimodal, comenzando, en la mayoría de los casos, durante la mañana, de 8:30 a 10:30 hs., y por la tarde desde 18 a 20:30 hs. La totalidad de las especies del grupo de L. anomalus habitan en el área fitogeográfica de Monte (Cabrera y Willink, 1980).
La mayoría de las especies de este grupo presentan algún grado de amenaza y aunque se conoce muy poco sobre su biología, el marcado endemismo sumado a la baja densidad poblacional, las ubica con un alto riesgo de extinción (Abdala et al., 2012).
El grupo de Liolaemus anomalus se distribuye desde el centrosur de Catamarca hasta el suroeste de La Pampa, en Argentina (Abdala, 2007a).
A primera vista, debido a su fisonomía (forma de la cabeza y cuerpo), dan la impresión de ser integrantes de algún género de policrótidos como Diplolaemus o Leiosaurus, y no del género Liolaemus (Abdala, 2007a). Si además tenemos en cuenta algunas características morfológicas singulares dentro del género (cola casi igual que el largo del cuerpo y con escasa autotomía, la cabeza más ancha o igual que su largo, fleje o "peine" palpebral desarrollado y bajo número de poros precloacales), es comprensible que se hayan realizado numerosos arreglos taxonómicos para las especies de este grupo (Cei, 1986, 1993; Etheridge, 1995; Abdala, 2007a).
El grupo de Liolaemus anomalus, luego de ser propuesto como género y subgénero por varios autores (Müller, 1928; Nuñez y Yañez, 1984; Laurent, 1984) fue ubicado por Etheridge (1995) basal en el grupo de L. signifer o "argentino" argumentando la hipertrofia del músculo puboisquiotibialis que cubre la capa media del músculo flexor tibialis interno, pero sin presencia de parche femoral. Sin embargo, en los estudios morfológicos desarrollados por Abdala et al. (2006) y Abdala (2007a), se pudo comprobar la presencia del parche femoral en todas las especies relacionadas al grupo de L. anomalus. En los análisis cladísticos de Evidencia Total (Abdala, 2007a), se recuperó el grupo de Liolaemus anomalus dentro del grupo de L. boulengeri (Fig. 1).
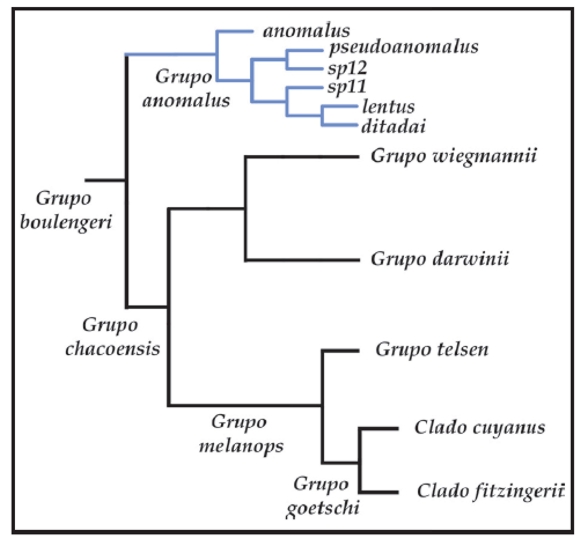
Figura 1. Hipótesis de relaciones propuesta por Abdala (2007), en base a caracteres morfológicos y moleculares. En azul, el grupo de Liolaemus anomalus. L. sp. 11 y L. sp. 12 corresponden a taxa que se describen en este trabajo.
La ubicación del grupo de Liolaemus anomalus, dentro del grupo de L. boulengeri es congruente con los análisis basados en caracteres moleculares de Schulte et al. (2000), Morando (2004), Espinoza et al. (2004), Avila et al. (2006, 2008, 2010) y el análisis basado en caracteres osteológicos de Lobo y Abdala (2002). En dichos trabajos se utilizó a L. pseudoanomalus como representante del grupo de L. anomalus. En todos estos trabajos, L. pseudoanomalus apareció como grupo hermano del grupo de L. wiegmannii, mientras que en el estudio de Abdala (2007a) el grupo de L. anomalus se ubicó como grupo hermano del grupo de L. boulengeri sensu Etheridge (1995) (Fig. 1).
Con respecto a las relaciones internas, dentro del grupo de Liolaemus anomalus la única hipótesis propuesta es la de Abdala (2007a), ya que es el único estudio previo en el cual se utilizan la totalidad de las especies descriptas para el grupo (Fig. 1).
Taxonomía y composición del grupo de Liolaemus anomalus
Inicialmente Burmeister (1861) describe Leiosaurus marmoratus, para la Planicie del oeste de Catamarca. Koslowsky (1896) describe a Liolaemus anomalus para la provincia de la Rioja, considerando esta especie intermedia con el género Liolaemus y Helocephalus.
Müller (1928) describe Phrynosaura werneri, especie de la que no menciona localidad tipo ni lugar aproximado donde fuera coleccionada, e incluye en este género a Phrynosaura marmoratus.
Gallardo (1966) describe Liolaemus lentus para la provincia de La Pampa, especie que luego fuera sinonimizada con Phrynosaura werneri por Donoso Barros (1969) sobre la base de la descripción original de L. lentus, sin observación del material tipo de ninguna de las dos especies. Solo se conocen los ejemplares con los cuales se realizó la descripción de L. lentus, ya que no se han vuelto a encontrar individuos de dicha especie en el lugar citado en su descripción original.
Posteriormente, Donoso Barros (1971) incluye en el género Ctenoblepharys, a Liolaemus anomalus y Leiosaurus marmoratus. Cei (1979), considera estas dos especies pertenecientes al género Liolaemus. Cei (1981) cambia de nombre a L. marmoratus por Liolaemus pseudoanomalus por estar ocupado marmoratus por Gravenhorst (1838). Cei (1979) sinonimiza a P. werneri y L. lentus con L. anomalus.
Nuñez y Yánez (1984) describen el género Abas que incluye a Liolaemus anomalus y L. pseudoanomalus y lo caracterizan por presentar párpados con peine breve, diámetro ocular menor que la longitud desde el borde anterior del ojo hasta la escama rostral, cola igual o menos larga que el largo hocico-cloaca, escudos de la cabeza diferenciados, perfil isognato, región loreal levemente deprimida y escamas dorsales imbricadas y sin quillas.
Laurent (1984) propone el género Ceiolaemus, incluyendo a Liolaemus anomalus y L. pseudoanomalus, con base en caracteres morfométricos, escamas dorsales sin quillas y presencia de dientes pterigoideos (erróneamente mencionados como palatinos).
Cei (1986, 1993) desestima como género a Ceiolaemus y lo propone como subgénero. Posteriormente, los géneros Abas y Ceiolaemus, fueron sinonimizado con Liolaemus por Etheridge (1995).
Cei (1983) describe como subespecie a Liolaemus anomalus ditadai, de las Salinas Grandes, en el límite entre Córdoba y Catamarca, Argentina. Esta especie está escasamente representada en los principales museos del país y fue hallada nuevamente luego de varios años de búsqueda (Abdala et al., 2012).
Espinoza et al. (2004), en un estudio filogenético relacionan a Liolaemus pseudoanomalus con L. duellmani, especie a priori perteneciente a otro grupo de Liolaemus (Grupo de L. montanus) debido a los caracteres morfológicos que presenta, claramente diferentes a los presentados por el grupo de L. anomalus. Esta hipótesis (la inclusión de L. duellmani en el grupo de L. anomalus) es tomada por Pincheira et al. (2008) y Lobo et al. (2010).
Abdala (2007a), evidencia la presencia del parche femoral en todas las especies del grupo de Liolaemus anomalus; recupera al grupo como clado monofilético, en posición basal del grupo de L. boulengeri (Fig. 1). Además Abdala (2007a) propone que las poblaciones de La Laja, en San Juan, y las del Salar de Pipanaco, Catamarca, forman parte de dos nuevas entidades, además revalida a L. lentus y eleva al rango de especie a L. ditadai.
En este trabajo se realiza un análisis taxonómico y filogenético del grupo de Liolaemus anomalus tomando como base el estudio de Abdala (2007a), en el cual además se redescribe a L. anomalus, L. ditadai, L.lentus y L. pseudoanomalus, se describen tres nuevas especies para el grupo, se aportan datos que actualizan el conocimiento de las diferentes especies que integran el grupo y se analizan las congruencias con las hipótesis presentadas por los diferentes autores. También se aportan datos generales de interés biológico que ayudan a un mejor entendimiento de las especies del grupo.
MATERIALES Y MÉTODOS
Los especimenes estudiados representan a ejemplares en su mayoría adultos de las cuatro especies conocidas para el grupo de Liolaemus anomalus y otras tres que se describen en este trabajo (Apéndice I).
Análisis filogénetico
Para el desarrollo de los análisis filogenéticos se tomo como base la matriz morfológica utilizada por Abdala (2007a), a la cual se incluyeron las siguientes especies: Liolaemus abaucan, L. boulengeri, L. calchaqui, L. canqueli, L. casamiquelai, L. cuyanus, L. crepuscularis, L. darwinii, L. donosobarrosi, L. dorbignyi, L. duellmani, L. espinozai, L. fitzingerii, L. goetschi, L. grosseorum, L. huacahuasicus, L. inacayali, L. josei, L. kingii, L. koslowskyi, L. laurenti, L. lavillai, L. lineomaculatus, L. loboi, L. mapuche, L. martorii, L. melanops, L. morenoi, L. multimaculatus, L. nigriceps, L. olongasta, L. ornatus, L. quilmes, L. rothi, L. sagei, L. salinicola, L. senguer, L. scapularis, L. sp 9, L. tehuelche, L. telsen, L. uspallatensis, L. wiegmannii y L. xanthoviridis. En el Apéndice II se incluyen solo las especies que presentan variaciones taxonómicas, en su distribución o se hayan estudiado nuevos ejemplares con respecto a la matriz de Abdala (2007a).
Se estudiaron los caracteres corrientemente utilizados en Liolaemus, descriptos o citados principalmente por Laurent (1985); Etheridge (1993, 1995, 2000); Cei (1986); Lobo (2001) y Abdala (2007a). La descripción de los colores en vida fue realizada en el campo y en base a fotografías tomadas al capturar los individuos. La terminología del patrón de coloración del cuerpo sigue a Abdala (2007a). Las observaciones de escamación y medidas corporales fueron tomadas utilizando lupa binocular (10-40x) y calibre de precisión de 0,02 mm. Los dibujos ilustrando los caracteres, fueron realizados utilizando cámara clara y un calibre de precisión de 0.1 mm.
Los ejemplares fueron coleccionados con lazo, fijados con formaldehído al 10% y conservados en alcohol 70%. Para las colectas se obtuvieron los permisos de las Direcciones de Fauna de cada Provincia.
La matriz morfológica está constituida por 50 taxa terminales y 142 caracteres. Se agregaron a los caracteres utilizados en Abdala (2007a), 11 nuevos caracteres: Carácter 52: Desarrollo del pliegue supraescapular: Binario polimórfico. Ausente (0); Presente (1). (Fig. 2A); Carácter 53: Desarrollo del bulto anterior en el pliegue supraescapular: Binario polimórfico. Ausente (0); Presente (1). (Fig. 2 B); Carácter 54: Desarrollo del bulto posterior en el pliegue supraescapular: Binario polimórfico. Ausentes (0); Presentes (1). (Fig. 2 B); Carácter 64: Mancha desde el ojo a supralabiales. Multiestado polimórfico. Ausente (0); Solo una línea diagonal posterior (1); Con una línea diagonal anterior discontinua y posterior presente (2); Presente con forma de "V" invertida (3). (Fig. 3-1); Carácter 65: Supralabiales manchadas alternadamente: Binario no polimórfico. Uniformes (0); Alternadas (1). (Fig. 3-1); Carácter 66: Infralabiales manchadas alternadamente: Multiestado no polimórfico. Uniforme (0); Difuso (1); Evidentes y marcados (2). (Fig. 3-1); Carácter 67: Anillos del brazo: Multiestado polimórfico. Ausentes (0); Difuso (1); Evidentes y marcados (2). (Fig. 3-2); Carácter 68: Anillos del antebrazo: Multiestado polimórfico. Ausente (0); Difuso (1); Evidentes y marcados (2). (Fig. 3-2); Carácter 69: Anillos del cuarto dedo de la mano: Multiestado polimórfico. Ausente (0); Difuso (1); Evidentes y marcados (2). (Fig.5); Carácter 139: Forma del borde de las escamas del parche femoral: Binario no polimórfico. Con borde redondeado (0); Con borde cuadrangular (1). (Fig. 3-3) y Carácter 140: Perfil de las escamas del parche femoral: Binario no polimórfico. Sin muesca (0); con muesca (1). (Fig. 3-3). De los 142 caracteres utilizados, 33 son continuos y 109 discretos, los cuales fueron discriminados en binarios no polimórficos, binarios polimórficos, multiestado no polimórfico y multiestado polimórfico. Siguiendo a Abdala (2007a) los caracteres multiestados, fueron discriminados en aditivos (postulando series de transformación del estado de carácter) y no aditivos (considerando un paso o costo para la transformación ente los estados de carácter). De los nuevos caracteres incluídos en este trabajo, se consideraron aditivos los caracteres 64, 67, 68 y 69. Los caracteres binarios polimórficos (Wiens, 1995) se trataron como tales. Los multiestados polimórficos fueron tratados como tales con los valores encontrados para cada taxón.

Figura 2. A: Carácter 52, exhibiendo la presencia del pliegue supra escapular. B: Carácter 53 y 54. Muestra la presencia de los bultos anterior y posterior del pliegue supraescapular
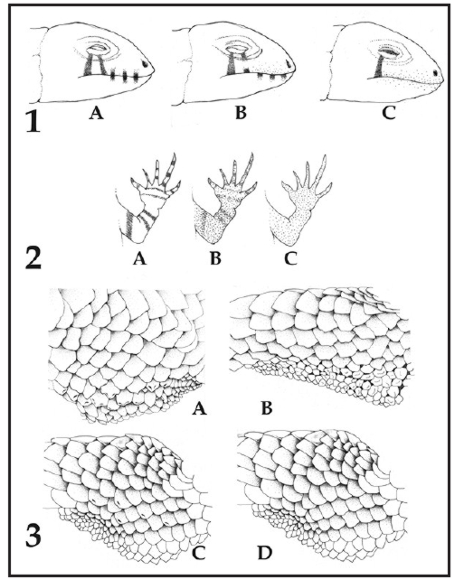
Figura 3. 1: Carácter 64, muestra las manchas desde el ojo a las supralabiales. A: Presente con forma de "V" invertida; B: Con una línea diagonal anterior discontinua y posterior presente C: Solo una línea diagonal posterior. 1: Caracteres 65 y 66, muestra las escamas supralabiales e infralabiales manchadas alternadamente. A: supralabiales e infralabiales manchadas. B: solo infralabiales manchadas alternadamente. C: con coloración uniforme, sin manchas alternadas en supra e infralabiales. 2: Carácteres 67, 68 y 69, muestra los anillos del brazo, antebrazo y del cuarto dedo de la mano. A: Evidentes y marcados B: difusos. C: Ausentes. 3: Caracteres 139 y 140, exhibiendo la forma y perfil de las escamas del parche femoral. A: Escamas con borde cuadrangular y con "muesca". B: Escamas con borde cuadrangular y sin "muesca". C: Escamas con borde redondeado y con "muesca". D: Escamas con borde redondeado y sin "muesca". Dibujos: V. Juárez H.
En el análisis filogenético se utilizó el criterio de parsimonia como criterio de optimalidad. El software utilizado para realizar la búsqueda de las hipótesis filogenéticas fue TNT 1.0 (Tree Analysis Using New Technology, versión 1.0; Goloboff et al., 2003) ya que es el único programa que permite el análisis de caracteres continuos sin convertirlos para su tratamiento en caracteres discretos. Los caracteres continuos fueron tratados utilizando la metodología propuesta por Goloboff et al. (2006), donde los caracteres continuos son analizados como tales, evitando la discretización de los mismos. Para cada carácter se utilizo el rango formado por la media ± desviación estándar. Como los caracteres continuos son tomados bajo diferentes escalas, un procedimiento conocido como "estandarización" o "reescalado" fue usado para evitar usar algunos caracteres con mayor influencia sobre otros en el análisis. La estandarización se realizó utilizando un script (mkstandb.run) asociado al software utilizado para los análisis (TNT, Goloboff et al., 2003). Con este script se uniformizan los costos máximos de transformación que puede haber entre dos caracteres continuos. De esta manera desde el estado más chico hasta el más grande, en un carácter continuo, se toma un valor específico respecto de lo que cuesta una transformación en un carácter discreto. Para este análisis se considero a 2 como mayor costo de transformación.
Las pruebas que se realizaron para buscar los árboles más cortos o con la cantidad menor de pasos fueron búsquedas heurísticas. La matriz fue tratada analizando los caracteres con pesos iguales y bajo pesos implicados (Goloboff, 1993). En este estudio se le dio el valor de 1 a 20 para "K". Para cada búsqueda se realizaron 500 replicaciones. A fin de obtener una medida de soporte de los resultados obtenidos se realizaron pruebas de remuestreo simétrico "Symmetric Resampling" con 500 réplicas con una probabilidad de deleción de 0,33.
Análisis estadístico
Para analizar posibles diferencias entre Liolaemus anomalus, Liolaemus pseudoanomalus y la especie que se describe en primer lugar, se trabajo con 18 individuos por especie, a los cuales se les midieron 8 caracteres morfométricos: Relación entre el largo y el ancho de la cabeza; Relación entre el ancho y el largo del torso; Relación entre el ancho del cuerpo o torso y el largo hocico-claoca; Relación entre la longitud del muslo y el largo del torso; Relación entre el alto y el ancho del oído; Relación entre el alto y el largo de la cabeza; Relación entre la longitud de la mano y el largo hocico-claoca y Relación entre el alto de la cabeza y el alto del oído.
Para el análisis de estos datos se utilizo el programa Statistica 7 (Statsoft, 2000), se seleccionó el modulo modelo general lineal, y el cual cuenta con el Análisis multivariante de la varianza (MANOVA). Se empleo este análisis estadístico debido a que se cuenta con numerosas variables dependientes (Caracteres de relación), que en forma conjunta podrían explicar un porcentaje de la variación encontrada entre las diferentes especies.
Los valores que se tuvieron en cuenta fueron: P (probabilidad), Lambda de Wilks (Varianza entre grupos/Varianza entre grupos + varianza dentro de grupos) y el del estadístico F. Cabe destacar que se trabajo con un a = 0,05.
RESULTADOS
Taxonomía
Redescripción de especies del grupo de Liolaemus anomalus
Liolaemus anomalus Koslowsky 1896
1896 Liolaemus anomalus. Koslowsky. Revista del Museo de La Plata 7: 447-457
1928 Phynosaura werneri. Müller. Zool. Anz. 77: 64.
1966 Liolaemus lentus. Gallardo. Neotropica 12(37): 17.
1969 Ctenoblepharis anomalus. Donoso Barros. Boln. Soc. Biol. Concepción 41: 93-94.
1970 Ctenoblephris anomalus. Peters y Donoso Barros, Bull. US natn. Mus.297, 2: 103.
1970 Phrynosaura werneri. Peters y Donoso Barros, Bull. US natn. Mus. 297, 2: 221.
1974 Ctenoblepharis werneri. Cei, J. Herp. 8(1): 71-75.
1979 Liolaemus anomalus. Cei, J. Herp. 13(2): 183-186.
1984 Ceiolaemus anomalus. Laurent, J. Herp. 18(4): 359.
1986 Liolaemus anomalus. Cei, J. Monografie IV: 527 pp.
1995 Liolaemus anomalus anomalus. Etherdige, R. American Museum of Nat. His. 3142, 34 pp.
1998 Liolaemus pseudoanomalus. Avila, L., Acosta, J.C. y Murúa, A. Cuad. Herp. 12 (1):11-29.
1998 Liolaemus pseudoanomalus. Acosta, J. y Murúa, F. Multequina 7: 49-59.
1998 Liolaemus anomalus. Acosta, J. y Murúa, F. Mul tequina 7: 49-59.
1999 Liolaemus anomalus. Marquez, J. Multequina 8: 1-10.
2000 Liolaemus anomalus. Avila, L., Morando, M. y Montero en: Lavilla, E., Richard, E. y Scrocchi, G. AHA. 97 pp.
2003 Liolaemus pseudoanomalus. Avila, L., Morando, M. y Belver, L. Herp. Soc. 38(3): 52-53.
2006 Liolaemus pseudoanomalus. Avila, L., Morando, M. y Sites Jr, J. W. Biol. Journal of the Linnean Society, 89: 241-275.
2007 Liolaemus anomalus. Abdala, C. S. Cuad. Herp. 21(1): 59-63.
Recapitulación histórica y elección de Lectotipo de Liolaemus anomalus
En los lotes de Liolaemus anomalus coleccionados por Julio Koslowsky en la provincia de La Rioja, depositados en el Museo de La Plata (MLP) y que él posiblemente usara para realizar la descripción de esta especie, solo encontramos 2 ejemplares, un macho (MLP S. 785) y una hembra (MLP S. 786), ambos adultos (Fig. 4).

Figura 4. A: Lectotipo y paralectotipo de Liolaemus anomalus. B: Lamina III de la descripción original de Liolaemus anomalus, Koslowsky (1986).
Este material estuvo perdido durante muchos años hasta que Williams y Cei (1983) dieron a conocer el redescubrimiento del material que utilizó Koslowsky para la descripción de esta especie en el MLP; brindaron las medidas de los dos ejemplares llamandolos a ambos "lectotipo"; las medidas brindadas en primer lugar corresponden al ejemplar MLP S. 785, y las medidas brindadas luego, al ejemplar MLP S. 786 (Ferraro y Williams, 2006).
Considerando la aparente pérdida del material tipo de Liolaemus anomalus, Cei (1979) designó un "neotipo" (MF-22232) para 40 km al oeste del Encón, provincia de San Juan, Argentina, y 18 "neoparatipos", categoría no aceptada por el Código Internacional de Nomenclatura Zoológica (Articulo 72.1.3; edición 2000) y por lo tanto no válidos. Esta población es considerada en este trabajo como una nueva especie (ver descripción de nuevas especies para el grupo de L. anomalus), no correspondiendo a una población de L. anomalus. Al existir el material original, la designación de "neotipo" (Cei, 1979) queda sin efecto (artículo 75.8 del Código Internacional de Nomenclatura Zoológica edición 2000).
En el trabajo original, Koslowsky (1896) realiza una descripción sin designar serie tipo ni holotipo, ni detallando en base a qué ejemplares realizó la descripción de Liolaemus anomalus, pero como dato importante incluye en la lámina III del trabajo seis excelentes fotos de dos ejemplares adultos (MLP S. 785 y 786) y un juvenil de L. anomalus (Fig. 4B).
Posteriormente Ferraro y Williams (2006) con base en la publicación original de Liolaemus. anomalus y las buenas fotografías que ésta contiene, designan como "lectotipo" al ejemplar macho MLP S. 785, y como "paralectotipo" al ejemplar hembra MLP S. 786 (Fig. 4A). Analizando y comparando las fotografías con los ejemplares colectados por Koslowsky coincidimos con la propuesta de Ferraro y Williams (2006) en la designación del lectotipo y paralectotipo, ya que se puede afirmar con alto grado de exactitud que las fotos corresponden a estos dos ejemplares de L. anomalus. Las fotos en vista ventral (izquierda) y dorsal (centro) corresponden al individuo MLP S. 785, las fotos en vista dorsal (derecha) y vista lateral izquierda de la cabeza corresponden al individuo MLP S. 786 (Fig. 4).
Asimismo, afianzamos la hipótesis de la designación del "lectotipo" MLP S. 785 en base a la descripción original. En dicho texto, Koslowsky menciona caracteres generales de la especie con caracteres tomados en particular de un ejemplar macho que coincidirían con el ejemplar MLP S. 785 (Fig. 4A). Koslowsky menciona: "Hay tres ó cuatro poros anales en el macho" (3 en el ejemplar MLP S. 785); "alrededor del medio cuerpo hay de 81 á 88 escamas" (85 en el ejemplar MLP S. 785); "grandes manchas plomizas sobre cada lado del dorso y de la cola, arregladas de tal manera que forman cinco fajas transversales sobre el cuerpo" (similar al ejemplar MLP S. 785).
Localidad Tipo
Koslowsky, en la descripción de Liolaemus anomalus, designa como localidad tipo "Provincia de la Rioja" en base a material coleccionado por él y por Hauthal (integrante del grupo expedicionario del Museo de La Plata). A principios de la década del 1890 Koslowsky realizó grandes expediciones por el oeste de las provincias de La Rioja y Catamarca, con el objetivo de relevar de forma exacta la geografía, geología, flora y fauna de dichas provincias.
La designación de la localidad tipo de Liolaemus anomalus (provincia de La Rioja) era aceptada para los estándares de descripción de especies de aquella época, sin embargo, hoy en día, al tener un mayor conocimiento del género Liolaemus, la designación de una localidad tipo poco precisa trae complicaciones a la hora de determinar con certeza la procedencia del material colectado por Koslowsky. Asimismo dentro de la provincia de La Rioja hay varias poblaciones de L. pseudoanomalus, especie cercana filogenéticamente a L. anomalus. Por este motivo, la poca exactitud de la localidad tipo trajo como consecuencia la confusión en la determinación de las verdaderas poblaciones de L. anomalus y L. pseudoanomalus. Con el pasar de los años la diferenciación y distribución de L. anomalus y L. pseudoanomalus se fueron confundiendo y mezclando, y de esta manera las colecciones herpetológicas del país fueron incluyendo y determinando estas especies sin un criterio claro.
Lamentablemente, los diarios de campo de Koslowsky se perdieron en un incendio (J. Williams, com. pers.), por lo que no podemos saber con exactitud el recorrido que realizó en sus expediciones y determinar con mayor precisión donde coleccionó los ejemplares de L. anomalus.
Sin embargo tenemos varias evidencias que nos acercan a una hipótesis firme. En primer lugar, los pueblos de Chilecito y Patquía, hacia fines del siglo XIX, eran pueblos grandes y lugar de paso casi obligado para los exploradores, con lo cual es probable que en algún punto de dicha región haya sido colectado el material tipo utilizado por Koslowsky para la descripción de L. anomalus. Otro punto importante que apoya esta hipótesis, es la publicación de Koslowsky (1895), "Batracios y reptiles de Rioja y Catamarca", en donde describe varios anfibios y reptiles de dichas provincias. En ese trabajo, Koslowsky, menciona en varios párrafos las localidades de Chilecito y Patquia, lugares donde colectaron ejemplares de reptiles y anfibios. Por ejemplo en la descripción de Hyla riojana, dice: "Esta especie fué hallada en una fuente que se llamaba «Aguadita» en el camino de Patquia a Chilecito, unos quince leguas de este pueblito, Provincia de la Rioja"; con lo cual tenemos la certeza que la ruta entre estos pueblos donde actualmente se encuentran ejemplares de L. anomalus era recorrida por los naturalistas. Conjuntamente, si tenemos en cuenta que en la descripción de L. anomalus, Koslowsky, dice: "Vive en la provincia de La Rioja; el señor Hauthal coleccionó el año pasado tres ejemplares en aquella provincia", refiriendose al año 1895, año de la expedición a La Rioja y Catamarca y que derivo en la publicación de Koslowsky (1895), muy probablemente Hauthal haya coleccionado los ejemplares de L. anomalus entre Patquía y Chilecito, en 1895. Si bien debemos hipotetizar en base a solo algunos datos puntuales y varios supuestos, pensamos que las evidencias son suficientes como para suponer que el lectotipo y paralectotipo provienen de esa zona de La Rioja.
En base a los estudios desarrollados en este trabajo, y teniendo en cuenta la descripción original de Liolaemus anomalus, la publicación de Koslowsky (1895), la comparación con el material colectado y examinado en diferentes museos, restringimos la localidad tipo de Liolaemus anomalus a las áreas en La Rioja comprendidas entre Chilecito y Patquia, ya que son las que mejor se corresponden con el material tipo colectado por Koslowsky. Con lo cual la especie se encontraría distribuida desde Chilecito - Nonogasta (entre los cordones de las sierras de Velasco al este y Famatina al oeste), a Patquía, Guayapa y Talampaya (Fig. 5).
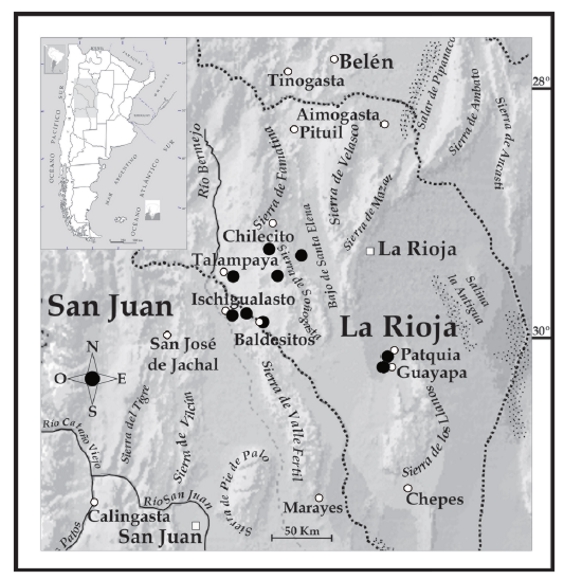
Figura 5. Distribución propuesta para Liolaemus anomalus. Los círculos negros corresponden a las diferentes poblaciones conocidas de L. anomalus.
Diagnosis
Liolaemus anomalus pertenece al grupo de L. boulengeri (Abdala, 2007a) por presentar un parche de escamas agrandadas en la cara posterior del muslo (Etheridge, 1995; Abdala, 2007a; Abdala et al., 2006). Dentro del grupo de L. boulengeri se diferencia de las especies del grupo de L. wiegmannii (Etheridge, 2000) (L. arambarensis, L. azarai, L. cranwelli, L. cuyumhue, L. lutzae, L. multimaculatus, L. occipitalis, L. rabinoi, L. riojanus, L. salinicola, L. scapularis y L. wiegmannii) por tener una hilera de escamas loreolabiales (nunca dos o tres), cuatro escamas en contacto con la mental (nunca seis), fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola. Se diferencia de las especies del grupo de L. darwinii (Abdala, 2007a) (L. abaucan L. albiceps, L. calchaqui, L. chacoensis, L. cinereus, L. crepuscularis, L. darwinii, L. diaguita, L. espinozai, L. grosseorum, L. irregularis, L. koslowskyi, L. laurenti, L. lavillai, L. montanezi, L. olongasta, L. ornatus, L. quilmes, L. uspallatensis) por tener cabeza casi tan larga como ancha, "fleje" o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola. También se diferencia por carecer de dicromatismo sexual evidente y línea negra transversal al ojo. Se distingue de las especies del grupo de L. melanops (Abdala, 2007a) (L. boulengeri, L canqueli, L. casamiquelai, L. chehuachekenk, L. cuyanus, L. donosobarrosi, L. fitzingerii, L. goestchi, L. hermannunezi, L. inacayali, L. josei, L. mapuche, L. martorii, L. melanops, L. morenoi, L. loboi, L. puelche, L. sagei, L. rothi, L. senguer, L. tehuelche, L. telsen y L. xanthoviridis) por tener dientes posteriores con coronas de bordes rectos, cabeza casi tan larga como ancha, fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola.
Dentro del grupo de Liolaemus anomalus, L. anomalus se diferencia de L. ditadai por tener un patrón de coloración claramente diferente (Fig. 6), ausencia de manchas dorsolaterales y postescapular evidente (presentes en L. ditadai), con presencia de anillos en el antebrazo, brazo y cola (sin anillos en L. ditadai), con menor cantidad o ausencia de escamas celestes en los lados del cuerpo y cola, región gular sin melanismo o con numerosas escamas oscuras, con presencia de bulto anterior y posterior en el pliegue supra-escapular (ausentes en L. ditadai) (Tabla 1), con mayor número de escamas alrededor del cuerpo (70-91; X= 77.6 vs. 60-68; X = 65.7), con mayor número de escamas ventrales (80-94; X= 85.3 vs. 72-82; X= 77.5), mayor número de escamas dorsales desde el occipucio hasta los miembros posteriores (84-106; X= 92.6 vs. 69-87; X= 80.3) (Tabla 2), mayor relación entre el ancho de la cabeza y largo hocico - cloaca, menor relación entre el largo y ancho de la cabeza y menor largo hocico - cloaca (max. LHC 68.6 vs. 92.6 mm) (Tabla 3).
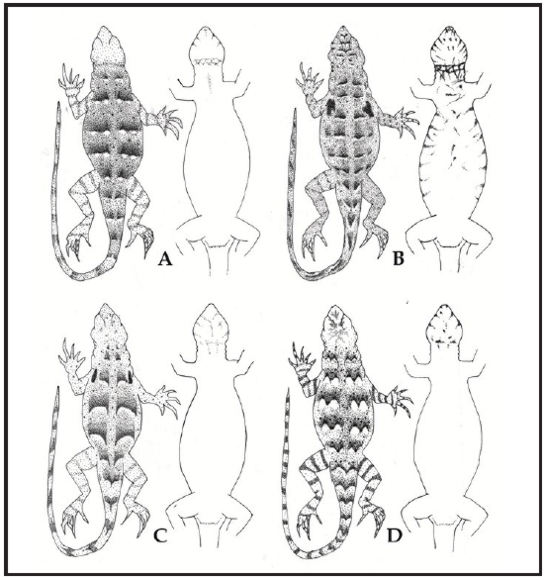
Figura 6. Diseño de los patrones de coloración dorsal y ventral de Liolaemus anomalus (A), Liolaemus ditadai (B), Liolaemus lentus (C) y Liolaemus pseudoanomalus (D). Dibujos: V. Juárez H.
Tabla 1. Exhibe algunos caracteres utilizados en la filogenia referidos al patrón de coloración dorsal y ventral, pliegue supra escapular y bultos en el pliegue supraescapular, exponiendo las diferencias entre las especies del grupo de L. anomalus.
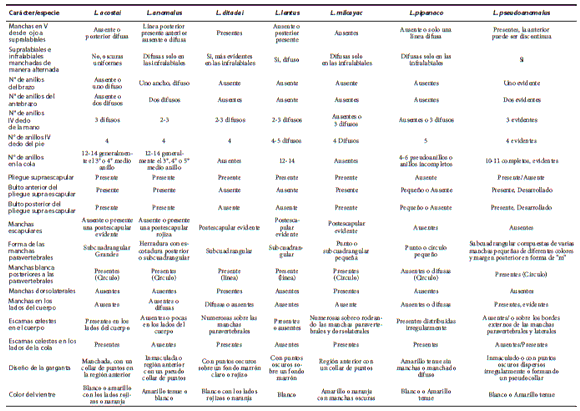
Tabla 2. Diferencias en el numero de escamas entre las especies del grupo de Liolaemus anomalus. Los valores indicados corresponden al rango entre el minimo y máximo y entre paréntesis se indica la media obtenida.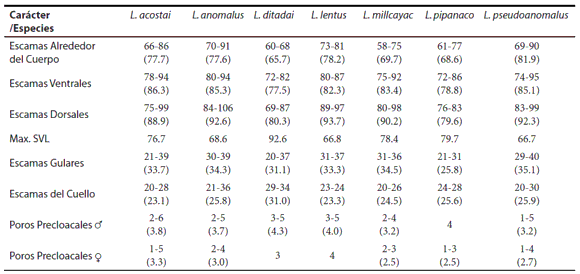
Tabla 3. Diferencias correspondientes a los caracteres morfométricos analizados entre las especies del grupo de Liolaemus anomalus. Los valores indicados corresponden al rango entre el minimo y máximo y entre paréntesis se indica la media obtenida.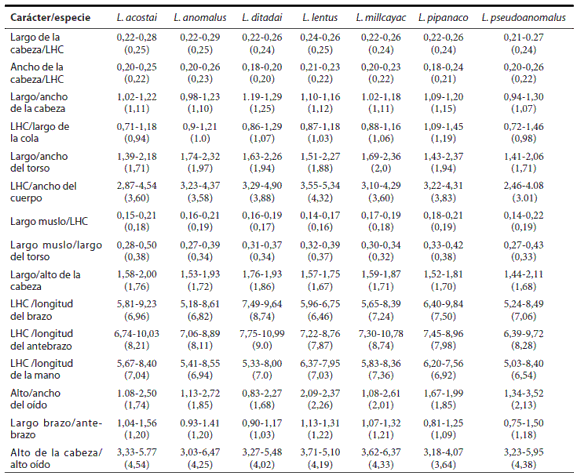
Se diferencia de Liolaemus lentus por tener un patrón de coloración diferente (Fig. 6), con ausencia de mancha postescapular negra evidente (presentes en L. lentus), con presencia de anillos oscuros en el antebrazo y brazo (sin anillos en L. lentus), con machos sin melanismo gular, con presencia de bulto anterior y posterior en el pliegue supra-escapular (ausentes en L. lentus) (Tabla 1) y menor relación entre el largo del muslo y largo hocico - cloaca (Tabla 3).
Se diferencia de Liolaemus pseudoanomalus por tener un patrón de coloración claramente diferente (Fig. 6), con forma, diseño y color de las manchas paravertebrales diferentes, con parte anterior de la mancha en "V" invertida desde el ojo a las supralabiales ausente o difusa (evidente en L. pseudoanomalus), supralabiales generalmente claras, nunca alternando manchas claras y oscuras como en L. pseudoanomalus anillos en el antebrazo, brazo y cola mucho más evidentes y definidos en L. pseudoanomalus, manchas en los lados del cuerpo difusas (evidentes en L. pseudoanomalus), mayor número de anillos en la cola (12-14 vs. 10-11) (Tabla 1), por tener diferencias significativas en las siguientes relaciones: Alto y ancho del oído; largo y ancho del torso y ancho del cuerpo/LHC (ver resultados de análisis estadístico y Tabla 3).
Descripción del Lectotipo
MLP.S 785 (Fig. 4A)
Macho adulto. Longitud hocico-cloaca 59.9 mm. La cabeza es 1.1 veces más larga (14.6 mm) que ancha (12.7 mm). Altura de la cabeza 7.9 mm. Ancho del cuello 10.0 mm. Interórbitas 8.6 mm. Diámetro longitudinal del ojo 4.8 mm. Longitud oído-ojo 5.6 mm. Alto del oído 2.4 mm y ancho 1,2 mm. Distancia del ojo al labio superior 1.7 mm y distancia entre las narinas 2.1 mm. Longitud de la subocular 3.4 mm. Longitud del torso 27.8 y de la cola 53.7 mm. Ancho del cuerpo 15.2 mm. Longitud del muslo 10.4 mm. Longitud de la tibia 9.7 mm. Longitud del pie 12.6 mm. Longitud del húmero 6.8 mm, antebrazo 7.3 mm y mano 7.7 mm. Longitud de la región pigal 6.5 mm y ancho de cloaca 8.1 mm.
Superficie dorsal de la cabeza lisa, con dieciséis escamas dorsales. Rostral más ancha que alta, en contacto con seis escamas. Mental de mayor tamaño que la rostral, de forma trapezoidal, en contacto con cuatro escamas. Sin contacto entre la nasal y rostral, separadas por una escama. Dos internasales más altas que anchas. Con siete escamas rodeando la nasal que está separada de la canthal por dos escamas. Hay ocho escamas entre la rostral y frontal. Frontal no dividida. Con dos postrostrales. Parietal mayor que interparietal y en contacto con tres escamas. Preocular separada de las loreolabiales por una escama. Tres superciliares y diez ciliares superiores. Borde anterior del oído con escamas auriculares diferenciadas; borde superior sin escama diferenciada. Con trece temporales, que carecen de quillas. Subocular en contacto con siete loreolabiales, sin contactar con la cuarta supralabial. Seis supraoculares. Siete supralabiales. Ocho loreolabiales. Con sies infralabiales, la segunda en contacto con dos escamas ventralmente. Siete escamas escudos. Segundo escudo separados por una escama.
Con 85 escamas alrededor del cuerpo. Con 95 escamas dorsales desde el occipucio hasta el miembro posterior, con el extremo posterior redondeado, yuxtapuestas y sin quilla. Con 27 hileras de escamas en el dorso. Con 84 escamas ventrales (desde mental hasta la cloaca, siguiendo la línea media ventral del cuerpo) más grandes que las dorsales, yuxtapuesto y laminares. Con 34 escamas gulares, yuxtapuesta y sin quillas. Sin pliegue gular. Con 36 escamas en el cuello, desde el borde posterior del oído hasta el hombro, por el pliegue longitudinal. Tres poros precloacales. Escamas antehumerales granulares, yuxtapuesta y bien diferenciadas del resto. Pliegue auricular, antehumeral y longitudinal marcados. Escamas del pliegue longitudinal lisas y sin quilla. Cuarto dedo de la mano con 16 escamas y del pie con 22. Con parche femoral. Escamas dorsales de la cola sin mucrón y con quilla, ventrales lisas. Infracarpales laminares, imbricadas y trífidas. Infratarsales laminares que terminan en un pequeño mucrón, imbricadas y lisas. Pliegue postescapular presente, bulbo anterior y posterior en el pliegue postescapular desarrolados (Fig. 2A y B). Escamas del parche cuadrangular sin muesca (Fig. 3-3B). Escamas del centro de la palma del pie simples (con quilla y mucrón).
Coloración en alcohol: Dorso y lados de la cabeza de color gris oscuro con numerosas escamas y pequeñas manchas oscuras. Subocular color gris verdoso con manchas oscuras. Supralabiales y loreolabiales de color gris uniforme e infralabiales presentan una alternancia de color claro y oscuro (Fig 4B).
Cuerpo dorsalmente con 5 series de manchas paraverterbrales de forma subcuadrangular y sin bandas dorsolaterales. Las manchas paravertebrales con mancha blanca posterior de forma circular. El dorso y lados del cuerpo de color gris claro, salpicado irregularmente de numerosas escamas de color negro (Fig. 7 A). Las escamas del dorso del cuerpo, con un patrón de manchado atigrado homogéneo. Carece de línea vertebral. Ausencia de mancha postescapular. Con pocas manchas celestes dispersas en los lados del cuerpo. Los miembros anteriores y posteriores son de color gris claro, con 1 anillo en el antebrazo, 2 en el brazo y 3 en el cuarto dedo de la mano levemente marcada (Fig. 3-2B). Cola de color gris claro, con manchas longitudinales y escasas escamas celestes.

Figura 7. A: Ejemplares adultos, macho (izquierda) y hembra (derecha) de Liolaemus anomalus de Ischigualasto, San Juan, Argentina. Foto: J. C. Acosta. B: Hembra adulta de Liolaemus anomalus de Baldecitos, San Juan, Argentina. Foto: E. Sanabria. C y D: Macho adulto de Liolaemus anomalus de cercanías de Nonogasta, La Rioja, Argentina. Fotos: C. S. Abdala.
Ventralmente la región mandibular es de color blanco con manchas desde las infralabiales y en la región anterior del cuello forman un pseudocollar de puntos. Vientre, miembros anteriores, posteriores, cloaca y cola son totalmente blancos.
Variación en Liolaemus anomalus
Basada en diecisiete ejemplares (nueve macho y ocho hembras). La superficie dorsal de la cabeza es lisa. Nasal rodeada de 6-8 (X= 7.0; DS= 0.5) escamas. Supralabiales 7-10 (X= 7.5; DS= 1.3), 7-10 (X= 8.2; DS= 0.7) loreolabiales dispuestas en una sola hilera. Supraoculares 5-6 (X= 5.6; DS= 0.5). Parietales de mayor tamaño que las interparietales, rodeada de 6-9 (X= 6.6; DS= 0.8) escamas. 12-16 (X= 14.2; DS= 1.2) escamas entre la rostral y el occipucio. Infralabiales 5-8 (X= 6.8; DS= 0.8). Gulares 30-39 (X= 34.2; DS= 2.7). Temporales 10-14 (X= 11.8; DS= 1.3) sin quillas. Oído siempre más alto (X= 2.0 mm; DS= 0.5) que ancho (X= 1.1 mm; DS= 0.3). Pliegues del cuello desarrollados. Cabeza más larga (X= 14.7 mm; DS= 1.6) que ancha (X= 13.0 mm; DS= 1.8), y alta (X= 8.2 mm; DS= 0.9). Longitud del tronco (X= 31.7mm; DS= 5.0). Longitud hocico - cloaca en machos (X= 57.8 mm; DS= 9.6) y en hembras (X= 57.5 mm; DS= 6.9). Longitud del brazo (X= 7.1 mm; DS = 0.5). Longitud del antebrazo (X= 8.5 mm; DS = 1,1). Longitud de la mano (X= 8.3 mm; DS= 1.1). Longitud del muslo (X= 10.7 mm; DS= 1.3). Número de escamas alrededor del cuerpo de 70-91 (X= 77.6; DS = 5.6). Escamas dorsales entre occipucio y muslos 84-106 (X= 92.6; DS= 5.1). Escamas dorsales con borde redondeado, sin quillas y yuxtapuestas. Cuarto dedo de la mano con 15-19 (X= 16.5; DS= 1.3) laminillas digitales. Cuarto dedo del pie con 21-25 (X= 23.1; DS= 1.4) laminillas digitales. Infracarpales con escamas laminares, imbricadas, trífidas. Infratarsales con escamas laminares, imbricadas y lisas. Parte posterior del muslo con parche femoral. Ventrales más grandes que las dorsales 80-94 (X= 85.3; DS= 3.8). Cola de mayor longitud (X= 1.0 mm; DS= 0.1) que la longitud hocico-cloaca. Machos con 2-5 (X= 3.6; DS= 1.0) poros precloacales, 7 de 7 hembras con 2-4 (X= 3.0; DS= 0.6) poros precloacales. Pliegue supraescapular presente, bulbo anterior y posterior en el pliegue postescapular desarrollados. Escamas del parche con bordes redondeados o cuadrangulares pero siempre sin muescas. Escamas trífidas en el centro de la palma del pie.
Variación de coloración en la especie (Fig. 7): Sin dicromatismo sexual evidente. El color de la cabeza puede ser muy diverso, siendo levemente más oscura que el cuerpo, varía desde el rosado o rojizo, al gris y castaño. En algunos ejemplares de Ischigualasto, el color de la cabeza es gris muy oscuro casi negro (Fig. 7A). En los lados de la cabeza, se destacan manchas alternas, claras y oscuras, difusas en las infralabiales y homogéneas en las supralabiales. La mancha en "V" invertida desde el ojo a las supralabiales, la posterior es completa y mientras que la anterior esta ausente o difusa. El cuerpo puede tener gran variedad de colores, castaño, rosado, rojizo, gris claro, gris oscuro y casi negro (estos dos ultimos se manifiestan solamente en algunos ejemplares de Ischigualasto, San Juan) (Fig. 7A). Sin arco negro antehumeral y línea vertebral. Las manchas paravertebrales tienen forma de herradura con escotadura posterior o subcuadrangulares, de color negro, rojizas o castaño oscuro, con mancha blanca posterior en forma de círculo (Fig. 7B y 7C). Las escamas del dorso del cuerpo tienen un patrón de manchado atigrado homogéneo. Sin manchas ni bandas dorsolaterales. Manchas difusas en los lados del cuerpo. Mancha postescapular ausente o difusa de color rojizo, nunca negra evidente. Los miembros varían del gris al castaño. Brazo con 1 anillo ancho difuso, 2 anillos difusos en el antebrazo y 2 o 3 anillos en el 4° dedo de la mano (Fig. 3-2B). Con 4 anillos en el cuarto dedo del pie. La cola presenta un color similar al de la región dorsal del cuerpo, con pocas escamas de color celeste en algunos ejemplares. Con 12 a 14 anillos completos en la cola, el 3°, 4° o 5° anillo es incompleto.
Ventralmente tienen color blanco en la región mandibular, gular, pectoral, abdominal, cloacal y cola. Hay algunas manchas desde la mandíbula hasta el cuello formando un pseudocollar; en algunos ejemplares estas manchas se pueden extender hasta la región abdominal. Algunos ejemplares pueden presentar manchas amarillas en la región gular, abdomen y fémur, con algunas escamas naranjas esparcidas irregularmente (Fig. 7D).
Distribución (Fig. 5)
Liolaemus anomalus se distribuye en el Bajo de Santa Elena, en La Rioja, entre Chilecito y Nonogasta (limitado entre los cordones de las sierras de Velasco al este y Famatina al oeste), hasta Patquia y Guayapa. Al noroeste hay poblaciones en Talampaya. Asimismo también asignamos las poblaciones de Baldecitos e Ischigualasto (provincia de San Juan) a Liolaemus anomalus (Fig. 5) con base en sus caracteres morfológicos.
Historia Natural
En los trabajos de Acosta y Murúa, 1998; Avila et al. (1998); Marquez et al. (1999), se cita como PC (Presencia Confirmada) a Liolaemus anomalus en el Parque Natural Ischigualasto y es considerada como "Máxima Prioridad de Conservación", pero la misma aparece como no amenazada en la lista oficial, lo que sería un punto a considerar ya que en estos trabajos también se cita la presencia como vulnerable a L. pseudoanomalus, considerada en este trabajo también una población de L. anomalus. La consideración de Maxima Prioridad de Conservacion para L. anomalus se debe a la poca información sobre su bio-ecología, su escasa presencia y al difícil avistaje en el campo.
J. C. Acosta (com. pers.), destaca que deben ser analizados la actividad humana relacionadas con la ganadería, sobre todo en el área de Hoyada y la entrada del parque, para estimar el impacto que los mismos tienen sobre la fauna y flora y descartar que esto sea un posible efecto de la escasa presencia de Liolaemus anomalus.
En la categorización de los anfibios y reptiles de Argentina (Lavilla et al., 2000), Liolaemus anomalus es considerada como Insuficientemente conocida (IC), mientras que en la ultima recategorización (Abdala et al., 2012), ha sido categorizada como Vulnerable (V).
Se conoce muy poco sobre su biología y comportamiento. Se han encontrado ejemplares de esta especie, al sur de Chilecito, a horas de la siesta con temperaturas que superaban los 35° C.
Liolaemus ditadai Cei 1983
1983 Liolaemus anomalus ditadai. Cei, J. Deserta 7: 172-178.
1995 Liolaemus ditadai. Etherdige, R. American Museum of Nat. His. 3142, 34 pp.
1986 Liolaemus ditadai. Cei, J. Monografie IV: 527 pp.
2000 Liolaemus anomalus. Avila, L., Morando, M. y Montero en: Lavilla, E., Richard, E. y Scrocchi, G. AHA. 97 pp.
2007 Liolaemus ditadai. Abdala, C. Cuad. Herp. 21(1): 59-63.
2007 Liolaemus ditadai. Abdala, C. Zootaxa 1538: 1-84.
El Holotipo corresponde a una macho adulto (MLP 701), colectado en las Salinas Grandes, a 4 km del límite Córdoba - Catamarca.
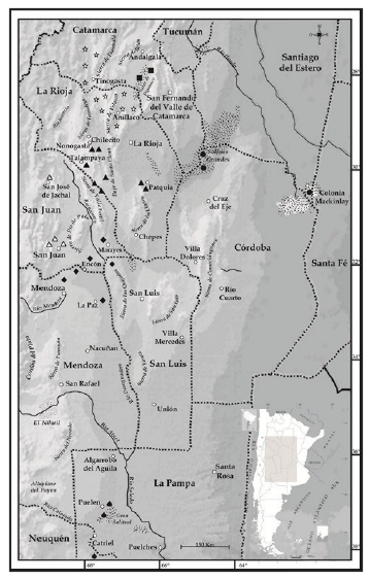
Figura 8. Mapa de distribución de las especies del grupo de Liolaemus anomalus. Triangulo Negro: Liolaemus anomalus. Círculo negro: Liolaemus ditadai. Gota negra: Liolaemus lentus. Estrella: Liolaemus pseudoanomalus. Triangulo blanco: Liolaemus acostai. Cuadrado negro: Liolaemus pipanaco. Rombo negro: Liolaemus millcayac.

Figura 9. A, B y C: Ejemplares machos adultos de Liolaemus ditadai de las Salinas Grandes, Córdoba, Argentina. A y C: Vista dorsal, B: Vista ventral. Fotos: C. S. Abdala.
Diagnosis
Liolaemus ditadai pertenece al grupo de L. boulengeri (Abdala, 2007a) por presentar un parche de escamas agrandadas en la cara posterior del muslo (Etheridge, 1995; Abdala, 2007a; Abdala et al. 2006). Dentro del grupo de L. boulengeri se diferencia de las especies del grupo de L. wiegmannii (Etheridge, 2000) (L. arambarensis, L. azarai, L. cranwelli, L. cuyumhue, L. lutzae, L. multimaculatus, L. occipitalis, L. rabinoi, L. riojanus, L. salinicola, L. scapularis y L. wiegmannii) por tener una hilera de escamas loreolabiales (nunca dos o tres), cuatro escamas en contacto con la mental (nunca seis), fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola. Se diferencia de las especies del grupo de L. darwinii (Abdala, 2007a) (L. abaucan L. albiceps, L. calchaqui, L. chacoensis, L. cinereus, L. crepuscularis, L. darwinii, L. diaguita, L. espinozai, L. grosseorum, L. irregularis, L. koslowskyi, L. laurenti, L. lavillai, L. montanezi, L. olongasta, L. ornatus, L. quilmes, L. uspallatensis) por tener cabeza casi tan larga como ancha, "fleje" o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola, también se diferencia por carecer de dicromatismo sexual evidente y línea negra transversal al ojo. Se diferencia de las especies del grupo de L. melanops (Abdala, 2007a) (L. boulengeri, L. canqueli, L. casamiquelai, L. chehuachekenk, L. cuyanus, L. donosobarrosi, L. fitzingerii, L. goestchi, L. hermannunezi, L. inacayali, L. josei, L. mapuche, L. martorii, L. melanops, L. morenoi, L. loboi, L. puelche, L. sagei, L. rothi, L. senguer, L. tehuelche, L. telsen, y L. xanthoviridis) por tener dientes posteriores con coronas de bordes rectos, cabeza casi tan larga como ancha, fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola.
Dentro del grupo de Liolaemus anomalus, L. ditadai se diferencia de L. anomalus por tener un patrón de coloración claramente diferente (Figs. 6 y 9), con presencia de manchas dorsolaterales y postescapular evidente (ausentes en L. anomalus), sin presencia de anillos en el antebrazo, brazo y cola (presentes en L. anomalus), con mayor cantidad de escamas celestes en los lados del cuerpo y cola, región gular melánica o con numerosas escamas oscuras, sin presencia de bulto anterior y posterior en el pliegue supra-escapular (presentes en L. anomalus) (Tabla 1), con menor número de escamas alrededor del cuerpo (60-68; X= 65.7 vs. 70-91; X= 77.6), con menor número de escamas ventrales (72-82; X= 77.5 vs. 80-94; X= 85.3), menor número de escamas dorsales desde el occipucio hasta los miembros posteriores (69-87; X= 80.3 vs. 84-106,;X= 92.6) (Tabla 2), menor relación entre el ancho de la cabeza y largo hocico - cloaca, mayor relación entre el largo y ancho de la cabeza (Tabla 3), y mayor largo hocico - cloaca (max. LHC 92.6 vs. 68.6 mm) (Tabla 2). Se diferencia de Liolaemus lentus por tener un patrón de coloración claramente diferente (Fig. 6), con presencia de manchas dorsolaterales (ausentes en L. lentus), mayor cantidad de escamas celestes en el dorso y lados del cuerpo y cola, cola nunca anillada (Tabla 1), menor número de escamas alrededor del cuerpo (60-68; X = 65.7 vs. 73-81; X= 78.2), menor número de escamas dorsales desde el occipucio hasta los miembros posteriores (69-87; X= 80.3 vs. 89-97; X= 93.7), mayor número de escamas en el cuello (29-34; X= 31.0 vs. 23-24; X= 23.3) (Tabla 2), menor relación entre el ancho de la cabeza y largo hocico - cloaca, mayor relación entre el largo y ancho de la cabeza, mayor relación entre el largo y alto de la cabeza, menor relación entre el largo del brazo y largo del antebrazo (Tabla 3), mayor largo hocico - cloaca (max. LHC 92.6 vs. 68.6 mm) (Tabla 2).
Se diferencia de Liolaemus pseudoanomalus por tener un patrón de coloración claramente diferente (Fig. 6), con mancha postescapular grande evidente, forma y diseño de las manchas paravertebrales diferentes, con manchas dorsolaterales (ausentes en L. pseudoanomalus), sin presencia de anillos en el antebrazo, brazo y cola (presentes en L. pseudoanomalus), con mayor cantidad de escamas celestes en los lados del cuerpo y cola, región gular melánica o con numerosas escamas oscuras, sin presencia de bulto anterior y posterior en el pliegue supra-escapular (desarrollados en L. pseudoanomalus) (Tabla 1), menor número de escamas alrededor del cuerpo (60-68; X = 65.7 vs. 76-90; X= 81.9), menor número de escamas dorsales desde el occipucio hasta los miembros posteriores (69-87; X= 80.3 vs. 85-99; X= 92.3), mayor número de escamas en el cuello (29-34; X= 31.0 vs. 20-30; X= 25.9) (Tabla 2), menor relación entre el ancho de la cabeza y largo hocico - cloaca (Tabla 3), y mayor largo hocico - cloaca (max. LHC 92.6 vs. 66.7 mm) (Tabla 2).
Variación en Liolaemus ditadai
Basada en seis ejemplares (tres macho y tres hembras). La superficie dorsal de la cabeza es lisa. Nasal rodeada de 7-8 (X= 7.5; DS= 0,5) escamas. Supralabiales 6-9 (X= 7.0; DS= 1.0), 6-9 (X= 8.0; DS= 1.1) loreolabiales dispuestas en una sola hilera. Supraoculares 4-6 (X = 5.0; DS = 0,9). Parietales de mayor tamaño que las interparietales, rodeada de 6-7 (X = 6.3; DS = 0,5) escamas. 14-15 (X= 14.1; DS= 0.4) escamas entre la rostral y el occipucio. Infralabiales 6-7 (X = 6.1; DS= 0.4). Gulares 20-37 (X= 24.8; DS= 6.7). Temporales 10-13 (X = 11.0; DS = 1.1) sin quillas. Oído siempre más alto (X = 2.1 mm; DS = 0,7) que ancho (X= 1,3 mm; DS= 0,3). Pliegues del cuello desarrollados. Cabeza más larga (X= 15.6 mm; DS= 4.5) que ancha (X= 12.5 mm; DS= 3.7), y alta (X= 8.4 mm; DS= 2.6). Longitud del tronco (X= 32.4 mm; DS= 7.3). Longitud hocico - cloaca en machos (X= 70.5 mm; DS= 15.6) y en hembras (X= 59.4 mm; DS= 21.0). Longitud del brazo (X= 7.3 mm; DS= 2.1). Longitud del antebrazo (X= 7.6 mm; DS= 2.6). Longitud de la mano (X= 9.2 mm; DS= 1.8). Longitud del muslo (X= 11.2 mm; DS= 2.7). Número de escamas alrededor del cuerpo de 60-68 (X= 63.1; DS= 3.0). Escamas dorsales entre occipucio y muslos 69-87 (X= 79.8; DS= 6.5). Escamas dorsales con borde redondeado, sin quillas y yuxtapuestas. Cuarto dedo de la mano con 13-17 (X= 15.0; DS= 1.4) laminillas digitales. Cuarto dedo del pie con 19-24 (X= 21.0; DS = 2.0) laminillas digitales. Infracarpales con escamas laminares, imbricadas, trífidas. Infratarsales con escamas laminares, imbricadas y lisas. Parte posterior del muslo con parche femoral. Ventrales más grandes que las dorsales 72-82 (X= 77.5; DS= 4.6). Cola de mayor longitud (X= 1.0 mm; DS= 0.1) que la longitud hocico-cloaca. Machos con 3-5 (X= 4.3; DS= 1.1) poros precloacales, 2 de 2 hembras con 3 (X= 3.0; DS= 0.0) poros precloacales. Pliegue postescapular presente, bulbo anterior y posterior en el pliegue postescapular ausentes. Escamas del parche con bordes redondeados o cuadrangulares pero siempre sin muescas (Fig. 3-3 B y 3D). Escamas de la palma del centro de pie simples (con quilla y mucrón).
Variación de coloración (Fig. 9): Dorso y lados de la cabeza de color castaño claro, con numerosas escamas manchas blancas y castaño oscuro. Subocular color castaño claro con manchas oscuras. Supralabiales e infralabiales presentan una alternancia de color blanco y castaño claro (Fig. 3-1A). Mancha en "V" invertida, desde el ojo hasta las infralabiales, presente, tanto la línea posterior como la anterior completas, claramente visibles (Fig. 3-1A)
Cuerpo con siete pares de manchas paraverterbrales, de forma subcuadrangular, de color castaño claro, cada una con una línea posterior de color negro. Con mancha blanca que bordea la mancha paravertebral en forma de línea. El color del dorso y lados del cuerpo, es castaño claro, salpicado con numerosas escamas de color blanco. Con numerosas escama celestes, que se encuentran limitadas a las manchas paravertebrales (Figs. 9A y 9C). Carece de línea vertebral. Sin bandas dorsolaterales. Las escamas del dorso del cuerpo con un patrón de manchado atigrado homogéneo. Manchas en los lados del cuerpo ausente o difuso. Con postescapular evidente, color negro. Los miembros anteriores y posteriores son de color castaño claro, con numerosas escamas de color blanco y celeste. Sin anillos en brazo y antebrazo, 2 o 3 anillos difusos en el 4° dedo de la mano (Fig. 3-2C). Cuarto dedo del pie con 4 anillo. La cola dorsal y lateralmente, en su base tiene el color castaño claro, pero sobresalen las manchas de color blanco. Con numerosas escamas de color celeste y ausencia de anillos en la cola.
Ventralmente la región mandibular, pecho, abdomen, miembros y cola de color blanco (Fig. 9B). Desde las infralabiales, salen manchas oscuras formando en la región gular, un pseudocollar. Con manchas difusas en el pecho y abdomen (Fig. 9B). En la región gular y lados del abdomen, con escamas de color naranja. La región del muslo con manchas amarillas (Fig. 9B).
Distribución (Fig. 8)
Liolaemus ditadai se distribuye en las Salinas Grandes, entre en límite de las provincias de Cordoba, Catamarca y La Rioja. La población que habita en Colonia Mackinlay, Departamento Rivadavia, provincia de Santiago del Estero, Argentina, y fuera asignada a L. ditadai por Abdala (2007b), deberá ser revisada con mayor detalle si se logra capturar más ejemplares a fin de determinar su verdadera identidad ya que con solo un ejemplar (mal fijado), es difícil establecer su entidad taxonómica.
Historia Natural
El trabajo realizado por Abdala (2007b), resalta lo difícil que fue poder hallar esta especie en numerosos viajes de campo a la localidad tipo y lugares aledaños. Esto podría deberse al gran anegamiento que sufrió el área de las Salinas Grandes, durante muchos años. Este último dato fue confirmado, por comentarios personales del Dr. Ricardo Martori y en observaciones realizadas en posteriores viajes realizados a la zona. En noviembre de 2009, efectuamos una búsqueda muy minuciosa e intensa, durante dos días, desde las primeras horas de la mañana hasta el anochecer. En dicha cruzada, logramos avistar y capturar solo dos ejemplares, uno cada día casi a la misma hora (18:00 hs), esto podría deberse a las altas temperaturas registradas. Otro factor que posiblemente incide directamente en la baja población, es la escasa vegetación que presenta el área.
Cei (1986), destaca que Liolaemus ditadai, son lagartos que viven bajo cuevas o abrigos ocasionales (piedras, ramas); que su dieta se compone de insectos y las hembras realizarían la puesta de huevos a fines de enero y comienzos de febrero.
En la categorización de los anfibios y reptiles de Argentina (Lavilla et al., 2000), Liolaemus ditadai es considerada como Insuficientemente conocida (IC), mientras que en la ultima recategorización (Abdala et al., 2012), ha sido categorizada como Amenazada (A).
Liolaemus lentus Gallardo 1966
1966 Liolaemus lentus. Gallardo. Neotropica 12(37): 15-29.
1969 Phrynosaura werneri. Donoso Barros, R. Bole tín Soc. de Biol. de Concepción 41: 93-94.
1979 Liolaemus anomalus. Cei, J. Journal of Herp. 13: 183-186.
1986 Liolaemus anomalus. Cei, J. Monografie IV: 527 pp.
2007 Liolaemus lentus. Abdala, C. Zootaxa 1538: 1-84.

Figura 10. Holotipo (A) y paratipos (B y C) de Liolaemus lentus.
El holotipo corresponde a una hembra adulta (MACN 22071 ) colectado en los Altos de Cochicó, Cochicó, Departamento Puelén, La Pampa, Argentina. La etimología de la especie hace referencia a la denominación de los pobladores de la zona donde fue hallada a esta lagartija: "lagarto lerdo".
Diagnosis
Liolaemus lentus pertenece al grupo de L. boulengeri (Abdala, 2007a) por presentar un parche de escamas agrandadas en la cara posterior del muslo (Etheridge, 1995; Abdala, 2007a; Abdala et al., 2006). Dentro del grupo de L. boulengeri se diferencia de las especies del grupo de L. wiegmannii (Etheridge, 2000) (L. arambarensis, L. azarai, L. cranwelli, L. cuyumhue, L. lutzae, L. multimaculatus, L. occipitalis, L. rabinoi, L. riojanus, L. salinicola, L. scapularis y L. wiegmannii) por tener una hilera de escamas loreolabiales (nunca dos o tres), cuatro escamas en contacto con la mental (nunca seis), fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola. Se diferencia de las especies del grupo de L. darwinii (Abdala, 2007a) (L. abaucan L. albiceps, L. calchaqui, L. chacoensis, L. cinereus, L. crepuscularis, L. darwinii, L. diaguita, L. espinozai, L. grosseorum, L. irregularis, L. koslowskyi, L. laurenti, L. lavillai, L. montanezi, L. olongasta, L. ornatus, L. quilmes, L. uspallatensis) por tener cabeza casi tan larga como ancha, "fleje" o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola, también se diferencia por carecer de dicromatismo sexual evidente y línea negra transversal al ojo. Se diferencia de las especies del grupo de L. melanops (Abdala, 2007a) (L. boulengeri, L. canqueli, L. casamiquelai, L. chehuachekenk, L. cuyanus, L. donosobarrosi, L. fitzingerii, L. goestchi, L. hermannunezi, L. inacayali, L. josei, L. mapuche, L. martorii, L. melanops, L. morenoi, L. loboi, L. puelche, L. sagei, L. rothi, L. senguer, L. tehuelche, L. telsen y L. xanthoviridis) por tener dientes posteriores con coronas de bordes rectos, cabeza casi tan larga como ancha, fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola.
Dentro del grupo de Liolaemus anomalus, Liolaemus lentus se diferencia de L. anomalus por tener un patrón de coloración diferente (Fig. 6), con presencia de mancha postescapular negra evidente (ausente en L. anomalus), sin presencia de anillos oscuros en el antebrazo y brazo (presentes en L. anomalus), con presencia en los machos de melanismo gular, con ausencia de bulto anterior y posterior en el pliegue supra-escapular (presentes en L. anomalus) (Tabla 1) y mayor relación entre el largo del muslo y largo hocico - cloaca (Tabla 3).
Se diferencia de Liolaemus ditadai por tener un patrón de coloración claramente diferente (Fig. 6), sin manchas dorsolaterales (presentes en L. ditadai), menor cantidad o ausencia de escamas celestes en el dorso y lados del cuerpo y cola, cola anillada (Tabla 1), mayor número de escamas alrededor del cuerpo (73-81; X= 78.2 vs. 60-68; X= 65.7), mayor número de escamas dorsales desde el occipucio hasta los miembros posteriores (89-97; X= 93.7 vs. 69-87 X= 80.3), menor número de escamas en el cuello (23-24; X= 23.3 vs. 29-34; X= 31.0) (Tabla 2), mayor relación entre el ancho de la cabeza y largo hocico - cloaca, menor relación entre el largo y ancho de la cabeza, menor relación entre el largo y alto de la cabeza, mayor relación entre el largo del brazo y largo del antebrazo (Tabla 3) y menor largo hocico - cloaca (max. 66.8 LHC 68.6 vs. 92.6 mm) (Tabla 2).
Se diferencia de Liolaemus pseudoanomalus por tener un patrón de coloración claramente diferente (Fig. 6), con mancha en "V" invertida desde el ojo a las supralabiales incompleta con barra anterior ausente (completa y evidente en L. pseudoanomalus), con mancha postescapular grande evidente, forma y diseño de las manchas paravertebrales diferentes, con mancha blanca posterior a las paravertebrales difusa y con forma de línea (marcada y forma circular en L. pseudoanomalus), sin presencia de anillos en el antebrazo y brazo y manchas en los lados del cuerpo (presentes en L. pseudoanomalus), con mayor cantidad de anillos en la cola (12-14 vs. 10-11) pero menos definidos que en L. pseudoanomalus, región gular melánica o con numerosas escamas oscuras y sin presencia de bulto anterior y posterior en el pliegue supra-escapular (desarrollados en L. pseudoanomalus) (Tabla 1).
Variación en Liolaemus lentus
Basada en dos ejemplares (un macho y una hembra). Para analizar la coloración además se utilizó la fotografía de Perez y Avila (2011). La superficie dorsal de la cabeza es lisa. Nasal rodeada de 7-9 (X= 7.3; DS= 0.6) escamas. Supralabiales 7-8 (X= 7.6; DS= 0.6), 7-8 (X= 8.3; DS=0.6) loreolabiales dispuestas en una sola hilera. Supraoculares 6-7 (X= 6.3; DS= 0,6). Parietales de mayor tamaño que las interparietales, rodeada de 6-7 (X= 6.6; DS= 0,6) escamas. 13-17 (X= 15.3; DS= 2.1) escamas entre la rostral y el occipucio. Infralabiales 6-8 (X= 7.0; DS= 1.0). Gulares 31-37 (X= 33.3; DS= 3.2). Temporales 12-15 (X= 13.0; DS= 1.1) sin quillas. Oído siempre más alto (X= 2.2 mm; DS= 0,3) que ancho (X= 1,0 mm; DS= 0.1). Pliegues del cuello desarrollados. Cabeza más larga (X= 15.3 mm; DS= 0.7) que ancha (X= 13.6 mm; DS= 0.2), y alta (X= 9.1 mm; DS= 0.5). Longitud del tronco (X= 26.6 mm; DS= 3.1). Longitud hocico - cloaca en machos (X= 60.8 mm; DS= 1.1) y en hembras (X= 67.9 mm; DS= 0.0). Longitud del brazo (X= 7.7 mm; DS= 0.1). Longitud del antebrazo (X= 9.4 mm; DS= 0.6). Longitud de la mano (X= 8.7 mm; DS= 0.3). Longitud del muslo (X= 9.7 mm; DS = 0.1). Número de escamas alrededor del cuerpo de 73-81 (X= 78.0; DS= 4.3). Escamas dorsales entre occipucio y muslos 89-97 (X= 93.6; DS= 4.1). Escamas dorsales con borde redondeado, sin quillas y yuxtapuestas. Cuarto dedo de la mano con 12-16 (X= 14.3; DS= 2.1) laminillas digitales. Cuarto dedo del pie con 19-22 (X= 20.3; DS= 1.5) laminillas digitales. Infracarpales con escamas laminares, imbricadas, trífidas. Infratarsales con escamas laminares, imbricadas y lisas. Parte posterior del muslo con parche femoral. Ventrales más grandes que las dorsales 80-87 (X= 82.3; DS = 4.4). Cola de mayor longitud (X= 1.0 mm; DS= 0.2) que la longitud hocico-cloaca. Machos con 3-5 (X= 4.0; DS= 1.4) poros precloacales, hembra con 4 (X= 4.0; DS= 0.0) poros precloacales. Con una cabeza alargada y angosta en la base y un cuello largo y levemente ensanchado en la base. Pliegue postescapular presente (Fig. 2 A), bulbo anterior y posterior en el pliegue postescapular ausentes. Escamas del parche con bordes redondeados o cuadrangulares pero siempre sin muescas (Fig. 3-3B y 3D). Escamas del centro de pie simples (con quilla y mucrón).
Coloración en alcohol (Fig. 10): Dorso y lados de la cabeza de color gris claro o gris oscuro, con algunas escamas de color castaño claro. Supralabiales e infralabiales presentan una alternancia de color blanco y castaño claro, más evidente en las infralabiales (Fig. 3-1). Región temporal superior e inferior sin escamas celestes. Mancha en "V" invertida desde el ojo hasta supralabiales, ausente o solo presente la línea posterior. Cuerpo, con 6 pares de manchas paraverterbrales en forma subcuadrangular, con manchas blancas posteriores en forma de línea. El dorso y lados del cuerpo es de color castaño claro, con escasas escamas blancas y celestes. Las escamas del dorso del cuerpo con un patrón de manchado atigrado homogéneo. Carece de línea vertebral, arco negro antehumeral, bandas dorsolaterales y manchas laterales. Con postescapular evidente. Los miembros anteriores y posteriores son de color gris, con escasas escamas de color blanco. Sin anillos en brazo y antebrazo, el 4° dedo de la mano con 2 o 3 anillos difusos (Fig. 3-2C). Cuarto dedo del pie con 4-5 anillos. La cola dorsal y lateralmente, en su base tiene el color gris, con 12 anillos, sin escamas celestes.
Ventralmente la región mandibular, pecho, abdomen, miembros y cola de color blanco. Mandíbula con manchas evidentes las cuales forman un collar en la región anterior al cuello extendiéndose hasta el vientre.
Coloración en vida: El color de la cabeza es gris claro. Con pocas escamas oscuras en el dorsa y lados de la cabeza. Mancha en "V" invertida desde el ojo hasta las supralabiales completa. El es de color castaño. Carece de línea vertebral, arco negro antehumeral, bandas dorsolaterales y manchas laterales. Las manchas paravertebrales en forma subcuadrangulares, de color negro, menos densa en el centro de la mancha, aparentando estar dividivas en dos. Sin escamas blancas y celestes entre las manchas paravertebrales. Con mancha postescapular evidente, de color negro. Escasas manchas celestes en la región lateral del cuerpo. Los miembros del mismo color que el cuerpo, con manchas oscuras y claras distribuidas irregularmente. Sin anillos en los brazos y antebrazos. Con 5 anillos cuarto dedo del pie. La cola presenta un color similar al de la región dorsal del cuerpo, con anillos anteriores completos, divididos en los lados de la cola, sin escamas celestes.
Distribución (Fig. 8)
Liolaemus lentus es conocido solo para las Salinas de Cochicó, departamento de Puelen, provincia de La Pampa, Argentina y para la provincia de Río Negro, 18.3 km al Norte de Ingeniero Huergo (33º56'22,3"S, 67º14'43,6"W, 294 m) según Perez y Avila (2011).
Historia Natural
Gallardo 1966, en las observaciones de la especie, describe que el Salitral de Cochico está rodeado por bardas, entre la vegetación arbustiva que crece en la periferia se destacan zampa blanca (Atriplex lampa), zampa crespa (Zampa undulata), jume negro (Suaeda divaricata). Es conocido por los pobladores como un lagarto lento por lo que se hace evidente en su nombre.
Es una especie que habita en un lugar donde se hace muy complicado su busqueda y encuentro debido a la falta de agua, las altas temperaturas y la baja densidad poblacional de esta especie. En los últimos 10 años se realizaron 6 campañas a la localidad tipo y zonas aledañas, sin conseguir ver ni capturar un solo ejemplar de Liolaemus lentus. Hay sectores donde supuestamente habita esta especie que esta muy antropizada, debido a la ganadería intensiva que ejercen los puesteros de la zona.
En la categorización de los anfibios y reptiles de Argentina (Lavilla et al., 2000), Liolaemus lentus es considerada como Insuficientemente conocida (IC), mientras que en la ultima recategorización (Abdala et al., 2012), ha sido categorizada como Amenazada (A).
Liolaemus pseudoanomalus (Cei 1981)
1861 Leiosaurus marmoratus. Burmeister. Reise durch die la Plata Staaten, Halle, 2: 1-538.
1928 Phynosaura marmoratus. Müller. Zool. Anz.77: 66.
1970 Phrynosaura marmoratus. Peters y Donoso Barros, Bull. US natn. Mus. 297(2): 221.
1971 Ctenoblepharys marmoratus. Donoso Barros. Herp. Review 3: 85.
1979 Liolaemus marmoratus. Cei, J. Herp. 13 (3): 297.
1980 Liolaemus marmoratus. Cei, J. Herp. 14 (2): 192
1981 Liolaemus anomalus. Cei, J. Herp. 15 (2): 253.
1984 Liolaemus pseudoanomalus. Laurent, J. Herp. 18 (4): 359.
1986 Liolaemus pseudoanomalus. Cei, J. Monografie IV: 527 pp.
2000 Liolaemus pseudoanomalus. Shulte, J., R. Macey, J., Espinoza, R. Biol. Journal of the Linnean Soc. 69: 75-102.
2000 Liolaemus anomalus. Avila, L., Morando, M. y Montero. En: Lavilla, E., Richard, E. y Scrocchi, G. AHA. 97 pp.
2001 Liolaemus pseudoanomalus. Lobo, F. y Abdala, C. Herp. 15 (2): 119-135.
2002 Liolaemus pseudoanomalus. Lobo, F. y Abdala, C. Herp. 16 (2): 137-150.
2003 Liolaemus pseudoanomalus. Avila, L., Morando, M. y Belver, L. Herp. Soc. 38(3): 52-53.
2004 Liolaemus pseudoanomalus. Morando, M. Tesis Doctoral U.N.T.
2006 Liolaemus pseudoanomalus. Abdala, V., Abdala, C. y Tulli, M.J. Zootaxa 1205: 5-68.
2006 Liolaemus pseudoanomalus. Halloy, M. y Castillo, M. Herp. 9 (2): 127-133.

Figura 11. Ejemplares machos adultos de Liolaemus pseudoanomalus. A y C: del Señor de la Peña, Anillaco, La Rioja, Argentina. B: de Palo Blanco, Fiambalá, Catamarca, Argentina. Fotos: C. S. Abdala
Fue descrita originalmente como Leiosaurus marmoratus, para luego ser considerada como integrante del género Phrynosaura (Müller, 1928) y posteriormente del género Ctenoblepharys (Donoso Barros, 1971). Cei (1979) ubica esta especie definitivamente en el género Liolaemus, pero como el nombre de Liolaemus marmoratus había sido utilizado por Gravenhorst (1838), Cei (1981) reemplaza a L. marmoratus por L. pseudoanomalus haciendo referencia a la afinidad de esta especie con Liolaemus anomalus. No se designa holotipo en la descripción original y Müller (1928) realiza una descripción más detallada de los ejemplares de la serie tipo utilizada por Burmeister (1861) para describir la especie. En la misma redescripción, Müller (1928) menciona que dichos ejemplares se encuentran en mal estado de conservación. La localidad tipo de L. pseudoanomalus, corresponde a la planicie estéril del oeste de Catamarca, cerca de Alpaquinchi y Anapa; también antes de entrar a la Quebrada de la Troya, al norte de Anillaco, Catamarca, Argentina.
Diagnosis
Liolaemus pseudoanomalus pertenece al grupo de L. boulengeri (Abdala, 2007a) por presentar un parche de escamas agrandadas en la cara posterior del muslo (Etheridge, 1995; Abdala, 2007a; Abdala et al., 2006). Dentro del grupo de L. boulengeri se diferencia de las especies del grupo de L. wiegmannii (Etheridge, 2000) (L. arambarensis, L. azarai, L. cranwelli, L. cuyumhue, L. lutzae, L. multimaculatus, L. occipitalis, L. rabinoi, L. riojanus, L. salinicola, L. scapularis y L. wiegmannii) por tener una hilera de escamas loreolabiales (nunca dos o tres), cuatro escamas en contacto con la mental (nunca seis), fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola. Se diferencia de las especies del grupo de L. darwinii (Abdala, 2007a) (L. abaucan L. albiceps, L. calchaqui, L. chacoensis, L. cinereus, L. crepuscularis, L. darwinii, L. diaguita, L. espinozai, L. grosseorum, L. irregularis, L. koslowskyi, L. laurenti, L. lavillai, L. montanezi, L. olongasta, L. ornatus, L. quilmes, L. uspallatensis) por tener cabeza casi tan larga como ancha, "fleje" o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola, también se diferencia por carecer de dicromatismo sexual evidente y línea negra transversal al ojo. Se diferencia de las especies del grupo de L. melanops (Abdala, 2007a) (L. boulengeri, L canqueli, L. casamiquelai, L. chehuachekenk, L. cuyanus, L. donosobarrosi, L. fitzingerii, L. goestchi, L. hermannunezi, L. inacayali, L. josei, L. mapuche, L. martorii, L. melanops, L. morenoi, L. loboi, L. puelche, L. sagei, L. rothi, L. senguer, L. tehuelche, L. telsen y L. xanthoviridis) por tener dientes posteriores con coronas de bordes rectos, cabeza casi tan larga como ancha, fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola.
Dentro del grupo de Liolaemus anomalus, se diferencia de L. anomalus, por tener un patrón de coloración claramente diferente (Fig. 6), con forma, diseño y color de las manchas paravertebrales diferentes, con mancha en "V" invertida desde el ojo a las supralabiales evidente (barra anterior ausente o difusa en L. anomalus), supralabiales alternando manchas claras y oscuras (generalmente claras en L. anomalus), anillos en el antebrazo, brazo y cola mucho mas evidentes y definidos en L. pseudoanomalus, manchas en los lados del cuerpo evidentes (difusas en L. anomalus), menor numero de anillos en la cola (10-11 vs. 12-14) (Tabla 1), y por tener diferencias significativas en las siguientes relaciones: Alto y ancho del oído; ancho y largo del torso y ancho del cuerpo/LHC (ver resultados de análisis estadístico y Tabla 3).
Se diferencia de Liolaemus ditadai por tener un patrón de coloración claramente diferente (Fig. 6), sin mancha postescapular, forma y diseño de las manchas paravertebrales diferentes, sin manchas dorsolaterales (presentes en L. ditadai), con anillos marcados en el antebrazo, brazo y cola (ausentes en L. ditadai), con menor cantidad o ausencia de escamas celestes en los lados del cuerpo y cola, región gular nunca melánica o con numerosas escamas oscuras, con bulto anterior y posterior en el pliegue supraescapular (ausentes en L. ditadai) (Tabla 1), mayor número de escamas alrededor del cuerpo (76-90; X=81.9 vs. 60-68; X= 65.7), mayor número de escamas dorsales desde el occipucio hasta los miembros posteriores (85-99; X= 92.3 vs. 69-87; X= 80.3), menor número de escamas en el cuello (20-30; X= 25.9 vs. 29-34; X= 31.0) (Tabla 2), mayor relación entre el ancho de la cabeza y largo hocico - cloaca, y menor largo hocico - cloaca (max. LHC 66.7 vs. 92.6 mm) (Tabla 3).
Se diferencia de Liolaemus lentus por tener un patrón de coloración claramente diferente (Fig. 6), con mancha en "V" invertida desde el ojo a las supralabiales evidente (con barra anterior ausente en L. lentus), sin mancha postescapular, forma y diseño de las manchas paravertebrales diferentes, con mancha blanca posterior marcada y forma circular (difusa y con forma de línea en L. lentus), con presencia de anillos en el antebrazo y brazo y manchas en los lados del cuerpo (ausentes en L. lentus), con menor cantidad de anillos en la cola (10-11 vs. 12-14) pero más definidos que en L. lentus, nunca región gular melánica o con númerosas escamas oscuras y con gran desarrollo de bulto anterior y posterior en el pliegue supra-escapular (ausentes en L. lentus) (Tabla 1).
Variación en Liolaemus pseudoanomalus
Basada en treinta y ocho ejemplares (veinte cinco macho y trece hembras). La superficie dorsal de la cabeza es lisa. Nasal rodeada de 6-8 (X= 7.0; DS= 0.7) escamas. Supralabiales 7-8 (X 7.2; DS= 0.4), 7-9 (X= 8.0; DS=0.5) loreolabiales dispuestas en una sola hilera. Supraoculares 5-7 (X= 6.0; DS= 0,7). Parietales de mayor tamaño que las interparietales, rodeada de 5-8 (X= 6.6; DS= 0,8) escamas. 13-18 (X= 15.2; DS= 1.1) escamas entre la rostral y el occipucio. Infralabiales 6-8 (X= 7.0; DS= 0.4). Gulares 29-30 (X= 35.0; DS= 3.1). Temporales 10-13 (X= 0.9; DS= 0.1). Pliegues del cuello desarrollados. Cabeza más larga (X= 13.4 mm; DS= 1.4) que ancha (X= 12.5 mm; DS= 1.1), y alta (X= 8.0 mm; DS= 0.8). Longitud del tronco (X= 31.8 mm; DS= 3.3). Longitud hocico - cloaca en machos (X= 55.9 mm; DS= 5.0) y en hembras (X= 49.5 mm; DS= 1.6). Longitud del brazo (X= 6.8 mm; DS= 0.8). Longitud del antebrazo (X= 8.0 mm; DS= 1.0). Longitud de la mano (X= 8.6 mm; DS= 1.0). Longitud del muslo (X= 10.3 mm; DS= 1.0). Número de escamas alrededor del cuerpo de 69-90 (X= 81.3; DS= 5.2). Escamas dorsales entre occipucio y muslos 83-99 (X= 92.0; DS= 4.0). Escamas dorsales con borde redondeado, sin quillas y yuxtapuestas. Cuarto dedo de la mano con 13-19 (X= 16.3; DS= 1.4) laminillas digitales. Cuarto dedo del pie con 22-28 (X= 24.0; DS= 1.5) laminillas digitales. Infracarpales con escamas laminares, imbricadas, trífidas. Infratarsales con escamas laminares, imbricadas y lisas. Parte posterior del muslo con parche femoral. Ventrales más grandes que las dorsales 74-95 (X= 85.1; DS= 5.5). Cola de mayor longitud (X= 0.9 mm; DS= 0.1) que la longitud hocico-cloaca. Machos con 1-5 (X= 3.2; DS= 0.8) poros precloacales, hembra con 1-4 (X= 2.7; DS= 0.8) poros precloacales. Con una cabeza corta y un cuello más ancho que la cabeza. Pliegue postescapular presente o a veces ausente (Fig. 2 A), bulbo anterior y posterior en el pliegue postescapular bien desarrolados (Fig. 2 B). Escamas del parche con bordes redondeados o cuadrangulares con muescas (Fig. 3-3A y 3C). Escamas del centro de pie simples (con quilla y mucrón).
Variación en coloracion en la especie (Fig. 11): Liolaemus pseudoanomalus presenta dicromatismo sexual leve. Dorso y lados de la cabeza de color rojizo o castaño oscuro, con numerosas manchas negras, escamas blancas ausente o escasas. Supralabiales e infralabiales presentan una alternancia de color claro y oscuro (Fig. 3-1A). La mancha en "V" invertida, desde el ojo hasta las infralabiales, presente, evidente, la anterior puede ser discontinua (Fig. 3-1A y 1B). Tronco con 5 series de manchas paraverterbrales en forma subcuadrangular de color rojizas, negras o castaño oscuro, compuesta de varias manchas pequeñas de color blanco azuladas. Con un margen porterior en forma de "m"; con mancha posterior en forma circular (Fig. 6). El color del torso y lados del cuerpo es castaño oscuro o rojizo. Las escamas del dorso del cuerpo tienen un patrón manchado atigrado homogéneo. Carece de línea vertebral y de bandas dorsolaterales. Con manchas laterales evidentes (Fig. 11 A y 11 B). Sin manchas escapulares. Los miembros anteriores y posteriores son de color castaño oscuro o rojizo, con escasas escamas de color blanco, sin escamas celestes. Miembros anteriores y posteriores con anillos muy marcados; brazo con 1 anillo, antebrazo con 2 anillos y 3 anillos en el 4° dedo de la mano (Fig. 3-2; 11). Cuarto dedo del pie con 4 anillos. La cola dorsal y lateralmente, de color castaño oscuro o rojizo. En los lados de la cola con muy pocas escamas color celeste, sin embargo en la mayoria de loe ejemplares estas escamas están ausentes. Con 11-12 anillos completos muy evidentes (Fig. 11 A y 11 B).
Ventralmente la región mandibular, miembros anteriores, cloaca y cola de color blanco. La garganta, pecho, parte central del vientre y fémur de color amarillo (Fig. 11C). Mandíbula con manchas difusas las cuales forman un pseudocollar en la región gular (Fig. 6). Las hembras presentan una diferencia leve con los machos en el diseño de las manchas paraverterbrales, de color rojizas más claras y con menor densidad de manchas negras y blancas.
Distribución (Fig. 8)
Liolaemus pseudoanomalus ha sido hallado en el suroeste de Catamarca en su Localidad tipo, en cercanias de Palo Blanco y Fiambalá. En la Rioja habita en las localidades de Anillaco, El señor de la Peña, Aimogasta y Antinaco.
Historia Natural
Burmeister (1861), describe un lagarto muy velóz y que trata de esconderse debajo de los cactus. Se caracteriza por ser una especie muy difícil de hallar, agresivo y crepuscular, sin embargo C. S. Abdala en sus viajes colecto algunos ejemplares a hora del mediodía con temperaturas elevadas. Viven en ambientes eremicos, pedregosos y tienden a esconderse en pequeñas cuevas en el suelo o bajo rocas. En cuanto al comportamiento Halloy (2006), describe numerosos pataleos acompañado de unos pocos cabeceos, al igual que también se observo el característico "tongue flicks" o lengüeteo.
En la categorización de los anfibios y reptiles de Argentina (Lavilla et al., 2000), Liolaemus pseudoanomalus es considerada como Insuficientemente conocida (IC), mientras que en la ultima recategorización (Abdala et al., 2012), ha sido categorizada como No Amenazada (NA).
Descripción de nuevas especies para el grupo de Liolaemus anomalus
Liolaemus acostai sp nov.
1986 Liolaemus anomalus. Cei, J. Monografie IV: 527 pp.
1986 Liolaemus pseudoanomalus. Cei, J. Monografie IV: 527 pp.
1997 Liolaemus pseudoanomalus. Acosta, J.C. y Murúa, A. Herp. Rev. 28 (1)
1998 Liolaemus pseudoanomalus. Avila, L., Acosta, J.C. y Murúa, A. Cuad. Herp. 12 (1):11-29.
2002 Liolaemus pseudoanomalus. Villavicencio, J., Acosta, J.C., Canovas, M. y Marinero, J. Multequina 11: 51-60.
2003 Liolaemus pseudoanomalus. Avila, L., Morando, M. y Belver, L. Herp. Soc. 38(3): 52-53.
2003 Liolaemus pseudoanomalus. Villavicencio, J., Acosta, J.C., Canovas, M. y Marinero, J. Rev. Esp. Herp. 17: 18-92.
2006 Liolaemus pseudoanomalus. Abdala, V., Abdala, C. y Tulli, M.J. Zootaxa 1205: 5-68.
2006 Liolaemus pseudoanomalus. Villavicencio, J., Acosta, J.C., Canovas, M. y Marinero, J. Bulletin of the Maryland Herp. Soc. vol. 42 n°1.
2006 Liolaemus pseudoanomalus. Canovas, M., Acosta, J.C., Villavicencio, J. y Marinero, J. Cuad. Herp., 19(2): 57-61.
2007 Liolaemus sp 11 Abdala, C. S. Zootaxa 1538: 1-84.
2007 Liolaemus pseudoanomalus. Villavicencio, J., Acosta, J.C., Marinero, J y Canovas, M. Amphibia- Reptilia 28: 163-165.
Holotipo.- FML 22438. Macho adulto. Baños del Salado. La Laja, 20 Km. al norte de San Juan capital, Departamento Albardón, Provincia de San Juan, Argentina. Col. C. S. Abdala, A. Laspiur, J. Nori y G. Scrocchi. Octubre de 2008.
Paratipos.- FML 22437; 439-443. Seis ejemplares. Mismos datos que holotipo.
FML 16336-337. Dos ejemplares. La Laja, departamento Albardón, Provincia de San Juan, Argentina. Col. C. S. Abdala, A. Laspiur, J. Marinero y E. Sanabria. Octubre de 2004.
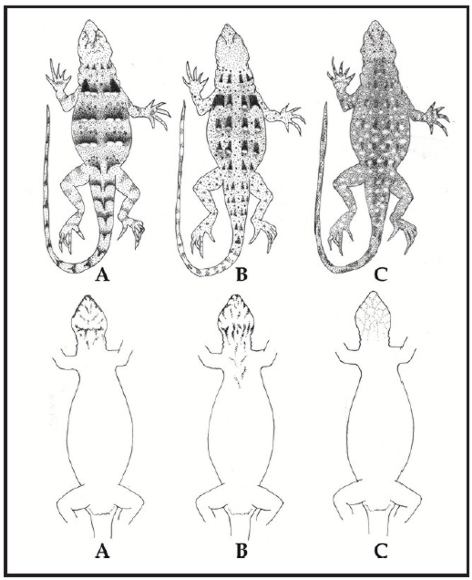
Figura 12. Diseño de los patrones de coloración dorsal y ventral de las nuevas especies del grupo de Liolaemus anomalus. A: Liolaemus acostai. B: Liolaemus millcayac. C: Liolaemus pipanaco. Dibujos: V. Juárez.

Figura 13. Ejemplares adultos de Liolaemus acostai sp. nov. de. La Laja, San Juan, Argentina. A y B: Holotipo (macho) C: Paratipo macho. D: Paratipo hembra. Fotos: C. S. Abdala.
Diagnosis
Liolaemus acostai pertenece al grupo de Liolaemus boulengeri (Abdala, 2007a) por presentar un parche de escamas agrandadas en la cara posterior del muslo (Etheridge, 1995; Abdala, 2007a; Abdala et al., 2006). Dentro del grupo de L. boulengeri se diferencia de las especies del grupo de L. wiegmannii (Etheridge, 2000) (L. arambarensis, L. azarai, L. cranwelli, L. cuyumhue, L. lutzae, L. multimaculatus, L. occipitalis, L. rabinoi, L. riojanus, L. salinicola, L. scapularis y L. wiegmannii) por tener una hilera de escamas loreolabiales (nunca dos o tres), cuatro escamas en contacto con la mental (nunca seis), fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola. Se diferencia de las especies del grupo de L. darwinii (Abdala, 2007a) (L. abaucan L. albiceps, L. calchaqui, L. chacoensis, L. cinereus, L. crepuscularis, L. darwinii, L. diaguita, L. espinozai, L. grosseorum, L. irregularis, L. koslowskyi, L. laurenti, L. lavillai, L. montanezi, L. olongasta, L. ornatus, L. quilmes, L. uspallatensis) por tener cabeza casi tan larga como ancha, "fleje" o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola, también se diferencia por carecer de dicromatismo sexual evidente y línea negra transversal al ojo. Se diferencia de las especies del grupo de L. melanops (Abdala, 2007a) (L. boulengeri, L canqueli, L. casamiquelai, L. chehuachekenk, L. cuyanus, L. donosobarrosi, L. fitzingerii, L. goestchi, L. hermannunezi, L. inacayali, L. josei, L. mapuche, L. martorii, L. melanops, L. morenoi, L. loboi, L. puelche, L. sagei, L. rothi, L. senguer, L. tehuelche, L. telsen, L. xanthoviridis) por tener dientes posteriores con coronas de bordes rectos, cabeza casi tan larga como ancha, fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola.
Dentro del grupo de Liolaemus anomalus, Liolaemus acostai se diferencia de L. anomalus por presentar un patrón de coloración diferente (Fig. 12), con presencia, en varios machos, de una mancha postescapular grande evidente y una preescapular pequeña, ausentes en L. anomalus, una mancha en "V" invertida desde el ojo a las supralabiales ausente o posterior difusa (marcada en L. anomalus), infralabiales generalmente oscuras, nunca alternando manchas claras y oscuras, manchas paravertebrales mas grandes y distinguibles, sin manchas en los lados del cuerpo, con algunas escamas celestes en los lados del cuerpo y cola, machos con lados del vientre rojizos o naranja (blanco o amarillo en L. anomalus) (Tabla 1), por tener mayor largo hocico - cloaca (max LHC 76.7 vs. 68.6 mm), menor relación entre el alto/ancho del oído menor y mayor relación entre el largo de la mano y el largo hocico - cloaca (ver resultados del análisis estadístico y Tabla 3).
Se diferencia de Liolaemus ditadai por tener un patrón de coloración claramente diferente (Fig. 12), color dorsal del cuerpo más oscuro, manchas paravertebrales de diferente forma y color (nunca celestes con el borde interno oscuro y borde externo posterior blanco), ausencia de manchas dorsolaterales (presentes en L. ditadai), menor cantidad de escamas celestes en los lados del cuerpo y cola, nunca región gular melánica o con numerosas escamas oscuras, cola anillada (sin anillos en L. ditadai), presencia de bulto anterior y posterior en el pliegue supra-escapular (ausentes en L. ditadai) (Tabla 1), mayor número de escamas alrededor del cuerpo (66-86; X= 77.7 vs. 60-68; X= 65.7), mayor número de escamas ventrales (78-94; X= 86.3 vs. 72-82; X= 77.5), menor número de escamas en el cuello (20-28; X= 23.1 vs. 29-34; X=31.0) y menor largo hocico - cloaca (max. LHC 76.7 vs. 92.6 mm) (Tabla 2).
Se diferencia de Liolaemus lentus por tener un patrón de coloración diferente (Fig. 12), manchas paravertebrales de diferente forma y color, con presencia de escamas celestes en los lados de la cola (ausentes en L. lentus), machos sin melanismo gular, presencia de bulto anterior y posterior en el pliegue supraescapular (ausentes en L. lentus) (Tabla 1), mayor largo hocico - cloaca (max. LHC 76.7 vs. 66.8 mm) (Tabla 2), menor relación entre el ancho del cuerpo y largo hocico cloaca, menor relación entre el largo y ancho del oido (Tabla 3).
Se diferencia de Liolaemus pseudoanomalus por tener un patrón de coloración diferente (Fig. 12), con presencia, en varios machos de una mancha postescapular grande evidente y una preescapular pequeña, forma, diseño y color de las manchas paravertebrales diferentes, mancha en "V" invertida desde el ojo a las supralabiales ausente o posterior difusa (evidentes en L. pseudoanomalus), infralabiales generalmente oscuras, nunca alternando manchas claras y oscuras como en L. pseudoanomalus, anillos en el antebrazo, brazo y cola mucho mas evidentes y definidos en L. pseudoanomalus que en L. acostai, ausencia de manchas en los lados del cuerpo (presentes en L. pseudoanomalus), mayor número de anillos en la cola (12-14 vs. 10-11) (Tabla 1), mayor largo- cloaca (max. LHC 76.7 vs. 66.7 mm) (Tabla 2), mayor relación entre el largo hocico - cloaca y el ancho del cuerpo (ver resultados del análisis estadístico y Tabla 3).
Descripción del holotipo (FML 22438) (Fig. 13A y 13B)
Macho adulto. Longitud hocico-cloaca 65.4 mm. La cabeza es 0.2 veces más larga (16.7 mm) que ancha (15.3 mm). Altura de la cabeza 9.9 mm. Ancho del cuello 13.4 mm. Interórbitas 10.2 mm. Diámetro longitudinal del ojo 5.7 mm. Longitud oído-ojo 7.1 mm. Alto del oído 2.9 mm y ancho 1,2 mm. Distancia del ojo al labio superior 2.2 mm y distancia entre las narinas 2.4 mm. Longitud de la subocular 3.7 mm. Longitud del torso 32.6 y de la cola 72.9 mm. Ancho del cuerpo 18.8 mm. Longitud del muslo 13.5 mm. Longitud de la tibia 11.2 mm. Longitud del pie 16.2 mm. Longitud del húmero 10.5 mm, antebrazo 8.7 mm y mano 9.6 mm. Longitud de la región pigal 8.0 mm y ancho de cloaca 9.4 mm.
Superficie dorsal de la cabeza lisa, con diecinueve escamas dorsales. Rostral más ancha que alta, en contacto con seis escamas. Mental de mayor tamaño que la rostral, de forma trapezoidal, en contacto con cuatro escamas. Sin contacto entre la nasal y rostral, separadas por una escama. Dos internasales más altas que anchas. Con ocho escamas rodeando la nasal que está separada de la canthal por dos escamas. Hay cinco escamas entre la rostral y frontal. Frontal no dividida. Con dos postrostrales. Parietal mayor que interparietal y en contacto con siete escamas. Preocular separada de las loreolabiales por dos escama. Tres superciliares y doce ciliares superiores. Borde anterior del oído con escamas auriculares diferenciadas; borde superior con escama diferenciada. Con doce temporales, que carecen de quillas. Subocular en contacto con sies loreolabiales, sin contactar con la cuarta supralabial. Seis supraoculares. Siete supralabiales. Nueve loreolabiales. Con ocho infralabiales, la segunda en contacto con dos escamas ventralmente. Seis escamas escudos. Segundos escudos están separados por dos escamas.
Con 77 escamas alrededor del cuerpo. Con 99 escamas dorsales desde el occipucio hasta el miembro posterior, con el extremo posterior redondeado, yuxtapuestas y sin quillas. Con 28 hileras de escamas en el dorso. Con 90 escamas ventrales (desde mental hasta la cloaca, siguiendo la línea media ventral del cuerpo) más grandes que las dorsales, yuxtapuesto y laminares. Con 21 escamas gulares, yuxtapuesta y sin quillas. Sin pliegue gular. Con 24 escamas en el cuello, desde el borde posterior del oído hasta el hombro, por el pliegue longitudinal. Tres poros precloacales. Escamas antehumerales granulares, yuxtapuesta y bien diferenciadas del resto. Pliegue auricular, antehumeral y longitudinal marcados. Escamas del pliegue longitudinal lisas y sin quilla. Cuarto dedo de la mano con 17 escamas y del pie con 26. Con parche femoral. Escamas dorsales de la cola sin mucrón y con quilla, ventrales lisas. Infracarpales laminares, imbricadas y trífidas. Infratarsales laminares que terminan en un pequeño mucrón, imbricadas y lisas. Pliegue postescapular presente, bulbo anterior y posterior en el pliegue postescapular desarrolados (Fig. 2 A y B). Escamas del parche con bordes redondeados sin muesca (Fig. 3-3D). Escamas del centro de pie simples (con quilla y mucrón).
Coloración en vida (Fig. 13)
Dorso y lados de la cabeza de color castaño, salpicada con algunas escamas pardo rojizas, con escasas escamas blancas. Supralabiales e infralabiales sin manchas alternas claras y oscuras (Fig. 3-1C). Mancha en "V" invertida, desde ojo hasta supralabiales, ausente o solo presente la linea posterior y difusa (Fig. 3-1C). Cuerpo con 5 pares de manchas paraverterbrales de forma subcuadrangular, de color castaño oscuro, con mancha blanca posterior en forma de circulo (Fig. 12). El dorso del cuerpo, miembros y cola de color castaño claro, pardo rojizo o rosado, salpicados con escamas de color blanco, celeste y negras. Carece de línea vertebral, bandas dorsolaterales y manchas en la región lateral. Sin mancha postescapular. Las escamas del dorso del cuerpo con un patrón de manchado atigrado homogéneo. Brazo con 1 anillo difuso, 2 anillos difusos en el antebrazo y 3 anillos en el 4° dedo de la mano (Fig. 3-2B). Con 4 anillos en el cuarto dedo del pie. Dorsalmente la cola presenta 5 anillos completos, el resto son incompletos. Cola sin escamas celestes.
Ventralmente la región mandibular, miembros anteriores, vientre, cloaca y cola son de color blanco (Figs. 12, 13B). En la región de la mandíbula, con manchas difusas que forman un pseudocollar en la región anterior del cuello (Fig. 12). Tanto la región gular, lados del vientre y fémur con manchas amarillas con algunas escamas naranjas (Fig. 13B).
Variación en Liolaemus acostai
Basada en treinta y siete ejemplares (veinticuatro macho y trece hembras). La superficie dorsal de la cabeza es lisa. Nasal rodeada de 6-8 (X= 7.0; DS= 0,5) escamas. Supralabiales 6-11 (X= 8.2; DS= 1.5), 6-11 (X= 7.8; DS= 1.2) loreolabiales dispuestas en una sola hilera. Supraoculares 5-8 (X= 6.1; DS= 0,9). Parietales de mayor tamaño que las interparietales, rodeada de 6-9 (X= 7.1; DS= 0,8) escamas. 12-19 (X= 14.9; DS= 1.3) escamas entre la rostral y el occipucio. Infralabiales 5-8 (X= 6.7; DS= 0.7). Gulares 21-39 (X= 33.7; DS= 3.6). Temporales 10-12 (X= 10.8; DS= 0.8) sin quillas. Oído siempre más alto (X= 1.9 mm; DS= 0,4) que ancho (X= 1,1 mm; DS= 0,2). Pliegues del cuello desarrollados. Cabeza más larga (X= 15.7 mm; DS= 1.9) que ancha (X= 14.1 mm; DS= 1.8) y alta (X= 8.5 mm; DS= 1.1). Longitud del tronco (X= 30.4 mm; DS= 5.3). Longitud hocico - cloaca en machos (X = 65.6 mm; DS = 7.4) y en hembras (X = 58.9 mm; DS = 7.5). Longitud del brazo (X= 8.8 mm; DS= 1.1). Longitud del antebrazo (X= 7.5 mm; DS= 1,1). Longitud de la mano (X= 8.7 mm; DS= 1.0). Longitud del muslo (X= 11.2 mm; DS= 1.3). Número de escamas alrededor del cuerpo de 66-86 (X= 77.8; DS= 5.0). Escamas dorsales entre occipucio y muslos 75-99 (X= 88.2; DS= 5.0). Escamas dorsales con borde redondeado, sin quillas y yuxtapuestas. Cuarto dedo de la mano con 13-18 (X= 15.7; DS= 1.0) laminillas digitales. Cuarto dedo del pie con 21-26 (X= 23.0; DS= 1.4) laminillas digitales. Infracarpales con escamas laminares, imbricadas, trífidas. Infratarsales con escamas laminares, imbricadas y lisas. Parte posterior del muslo con parche femoral. Ventrales más grandes que las dorsales 78-94 (X= 86.2; DS= 4.6). Cola de mayor longitud (X= 0.94 mm; DS= 0.1) que la longitud hocico-cloaca. Machos con 2-6 (X= 3.8; DS= 1.1) poros precloacales, 13 de 13 hembras con 1-5 (X= 3.2; DS= 1.0) poros precloacales. Con cabeza alargada y cuello comprimido (o marcada estrangulación). Pliegue postescapular presente, bulbo anterior y posterior en el pliegue postescapular desarrollados (Fig. 2 A y B). Escamas del parche con bordes redondeados sin muesca (Fig. 3-3D). Escamas del centro de pie trífidas.
Variación de coloración en la especie (Fig. 13C y 13D): Sin dicromatismo sexual evidente. El color de la cabeza varía desde gris a castaño oscuro, con abundantes manchas pardo rojizas y negras, siendo siempre más oscuro que el cuerpo. Región dorsal con escasas escamas celestes o blancas esparcidas irregularmente. Mancha en "V" invertida desde el ojo hasta las supralabiales ausente o posterior difusa (Fig. 3-1C). Supralabiales e infralabiales de color uniforme, sin manchas alternas (Fig. 3-1C). El cuerpo puede tener gran variedad de colores, castaño oscuro, rosado y pardo rojizo. Carece de línea vertebral, arco negro antehumeral, bandas dorsolaterales y manchas en la región lateral. Las manchas paravertebrales en forma subcuadrangulares grandes, de color negro o castaño oscuro, con mancha posterior blanca en forma de punto. Es común observar escamas blancas, celestes entre las manchas paravertebrales. Con manchas postescapular evidente de color negro, en algunos ejemplares puede estar ausente o ser tenue. Las escamas del dorso del cuerpo con un patrón de manchado atigrado homogéneo. Con gran número de escamas y pequeñas manchas celestes y blancas en la región lateral del cuerpo. Los miembros varían del gris al castaño oscuro. Brazo con 1 anillo difuso o ausente, 2 anillos difusos o ausentes en el antebrazo y 3 anillos en el 4° dedo de la mano (Fig. 3-2B). Con 4 anillos en el cuarto dedo del pie. La cola presenta un color similar al de la región dorsal del cuerpo, lateralmente con escamas celestes. Con 12 a 14 anillos, el 3° o 4° es incompleto formando un medio anillo.
Ventralmente la región gular, pectoral, abdominal, cloacal y cola de color blanco. Hay manchas difusas desde la mandíbula hasta el cuello donde forman un collar. Tanto la región gular y muslo, con manchas amarillas y lados del vientre con algunas escamas y manchas grandes de color naranja.
Distribución (Fig. 8)
Esta nueva especie fue encontrada en su localidad tipo, La Laja, en el Barreal de Matagusano, ambas localidades del departamento de Albardón, también a 20 km al norte de Caucete, departamento de Caucete y en San José de Jachal, todas localidades de la provincia de San Juan, Argentina.
Etimología
Esta especie es en honor a nuestro amigo el Lic. Juan Carlos Acosta, docente e investigador de la Universidad Nacional de San Juan el cual ha trabajado para el crecimiento de la herpetología Argentina y especialmente de la provincia de San Juan durante m ás de 20 años.
Historia Natural
Gracias a los trabajos realizados de Villavicencio et al. (2002, 2003, 2006, 2007), es posible conocer un poco más sobre la nueva especie. Según Villavicencio (2003), los machos tienen mayor longitud hocico-cloaca que las hembras, pero no presentan diferencias en el uso del espacio (Villavicencio et al., datos no publicados), ni en su alimentación (Belver y Avila, 1999), por lo que estos factores no serían las causas del dimorfismo sexual. Sin embargo se podría considerar la hipótesis de que este dimorfismo sería la consecuencia de una intensa agresividad entre los machos por el acceso de las hembras y por la defensa territorial (Villavicencio et al., 2003).
Villavicencio et al. (2002), destacan una "termoconformidad" mucho más heliotérmica que tigmotérmica para esta especie, no detectándose diferencias de temperatura corporal entre sexos, edad o estaciones (Villavicencio et al., datos no publicados). La termoconformidad permite que la temperatura corporal cambie en los individuos activos siguiendo el ritmo de variación de la temperatura ambiental. La mayoría de los ejemplares se encontraron bajo rocas (Villavicencio et al., 2006) y unos pocos en lugares abiertos. El ambiente donde viven se caracteriza por ser homogéneo con heterogeneidad espacial y cobertura baja. Tienen un patrón de actividad unimodal con picos de actividad de 9 a 11 hs., siendo la estación primavera-verano las de mayor actividad.
Debemos alertar sobre el estado de estas poblaciones, principalmente la de La Laja, San Juan (Loc. Tipo), que en los últimos 12 años ha sido diezmada, increíblemente por profesores y estudiantes de biología de la Universidad de San Juan. Dichos docentes y alumnos concurrían asiduamente al área a realizar diferentes investigaciones y/o actividades docentes de campo, pero al mismo tiempo colectaban sin sentido alguno y fácilmente bajo las "piedras lajas" numerosos ejemplares de esta lagartija. Hasta el año 2004 era muy fácil encontrar ejemplares en la zona, en el año 2008 la población era muy escasa y actualmente hay que recorrer intensamente mayores recorridos en el área y durante varias horas, para observar un ejemplar de Liolaemus acostai.
Liolaemus pipanaco sp nov.
2003 Liolaemus pseudoanomalus. Avila, L., Morando, M. y Belver, L. Herp. Soc. 38(3): 52-53.
2007 Liolaemus sp 12 . Abdala, C. S. Zootaxa 1538: 1-84.
Holotipo.- FML 19225. Macho adulto. Salar de Pipanaco. Entrando por puesto Rio Blanco, a 36 Km al sur de Andalgalá, Departamento de Andalgalá, Provincia de Catamarca, Argentina. S 27°49'42,4" O 66°14'59,0" 737 m s.n.m. Col: C. S. Abdala, J. P. Juliá, S. Quinteros, G. Scrocchi y J. C. Stazzonelli. Marzo de 2007.
Paratipos.- MCN 2622-23. Mismos datos que holotipo. FML 18396. Pasando por puesto Rio Blanco, al oeste para llegar al Salar de Pipanaco, Departamento de Andalgalá, Provincia de Catamarca, Argentina. S 27°49'23.9" O 65°14'38.2", 731 m s.n.m. Col: C. S. Abdala, S. Quinteros y G. Scrocchi. Octubre de 2007.

Figura 14. Ejemplares de Liolaemus pipanaco sp. nov. del Salar de Pipanaco, Catamarca, Argentina. A y B: Holotipo (macho adulto). C: Paratipo macho adulto. D: Paratipo hembra adulta. E: Paratipo macho juvenil. Fotos: C. S. Abdala.
Diagnosis
Liolaemus pipanaco pertenece al grupo de L. boulengeri (Abdala, 2007a) por presentar un parche de escamas agrandadas en la cara posterior del muslo (Etheridge, 1995; Abdala, 2007a; Abdala et al., 2006). Dentro del grupo de L. boulengeri se diferencia de las especies del grupo de L. wiegmannii (Etheridge, 2000) (L. arambarensis, L. azarai, L. cranwelli, L. cuyumhue, L. lutzae, L. multimaculatus, L. occipitalis, L. rabinoi, L. riojanus, L. salinicola, L. scapularis, L. wiegmannii) por tener una hilera de escamas loreolabiales (nunca dos o tres), cuatro escamas en contacto con la mental (nunca seis), fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola. Se diferencia de las especies del grupo de L. darwinii (Abdala, 2007a) (L. abaucan L. albiceps, L. calchaqui, L. chacoensis, L. cinereus, L. crepuscularis, L. darwinii, L. diaguita, L. espinozai, L. grosseorum, L. irregularis, L. koslowskyi, L. laurenti, L. lavillai, L. montanezi, L. olongasta, L. ornatus, L. quilmes, L. uspallatensis) por tener cabeza casi tan larga como ancha, "fleje" o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola, también se diferencia por carecer de dicromatismo sexual evidente y línea negra transversal al ojo. Se diferencia de las especies del grupo de L. melanops (Abdala, 2007a) (L. boulengeri, L canqueli, L. casamiquelai, L. chehuachekenk, L. cuyanus, L. donosobarrosi, L. fitzingerii, L. goestchi, L. hermannunezi, L. inacayali, L. josei, L. mapuche, L. martorii, L. melanops, L. morenoi, L. loboi, L.puelche, L. sagei, L. rothi, L. senguer, L. tehuelche, L. telsen, L. xanthoviridis) por tener dientes posteriores con coronas de bordes rectos, cabeza casi tan larga como ancha, fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola.
Dentro del clado de Liolaemus anomalus, L. pipanaco se diferencia de L. acostai por presentar un patrón de coloración dorsal claramente diferente (Fig. 12), con manchas paravertebrales menos marcadas, de menor tamaño, en forma de punto o circulo, nunca subcuadrangulares, color del cuerpo castaño o gris claro nunca gris oscuro, castaño o rojizo, ausencia de mancha postescapular (presentes en algunos machos de L. acostai), menor número de anillos en la cola (4-5 incompletos vs. 12-14 completos), ausencia de pliegue supraescapular, bultos en los extremos del pliegue supraescapular ausentes o menos marcado que en L. acostai, machos nunca con grandes manchas rojizas en los lados del vientre (Tabla 1), menor número de escamas dorsales desde el occipucio hasta los miembros posteriores (76-83; X= 79.6 vs. 80-99; X= 88.9) (Tabla 2) y mayor relación entre largo hocico - cloaca y largo de la cola (Tabla 3).
Se diferencia de Liolaemus anomalus por tener un patrón de coloración dorsal claramente diferente (Fig. 12), con mayor cantidad de escamas celestes y/o blancas en el dorso y lados del cuerpo y cola, por tener las manchas paravertebrales más difusas, pequeñas y menos marcadas, ausencia de anillos oscuro en el antebrazo y brazo (presentes en L. anomalus), menor número de anillos en la cola (4-5 incompletos vs. 12-14 completos), ausencia de pliegue supraescapular, bultos en los extremos del pliegue supraescapular ausentes o menos marcado que en L. anomalus (Tabla 1), menor número de escamas dorsales desde el occipucio hasta los miembros posteriores (76-83; X= 79.6 vs. 84-106; X= 92.6), menor número de escamas gulares (21-31; X= 25.8 vs. 30-39 x= 34.3) y mayor largo hocico-cloaca (max LHC 79.7 mm vs 68.6 mm) (Tabla 2).
Se diferencia de Liolaemus ditadai, por tener un patrón de coloración dorsal claramente diferente (Fig. 12), con las manchas paravertebrales más difusas, más pequeñas y menos marcadas, por carecer de manchas escapulares y manchas dorsolaterales, por tener menos escamas celestes en el dorso del cuerpo, con región anterior de la cola con 4-5 anillos incompletos (ausentes en L. ditadai) (Tabla 1), ausencia de pliegue supraescapular, menor número de escamas en el cuello (24-28; X= 25.6 vs. 29-34; X= 31.0), por tener menor longitud hocico-cloaca (max LHC 79.7 mm vs 92.6 mm) (Tabla 2), con menor relación entre el largo y ancho de la cabeza,mayor relación entre el largo hocico - cloaca y largo de la cola y presentar menor relación entre el largo y alto de la cabeza (Tabla 3)
Se diferencia de Liolaemus lentus por tener un patrón de coloración dorsal claramente diferente (Fig. 12), con las manchas paravertebrales más difusas, más pequeñas y menos marcadas, por carecer de manchas escapulares, con mayor cantidad de escamas celestes y/o blancas en el dorso y lados del cuerpo y cola, con menor número de anillos en la cola (4-5 incompletos vs. 12-14 completos), ausencia de pliegue supraescapular (Tabla 1), por tener menor número de escamas dorsales desde el occipucio a miembros posteriores (76-83; X= 79.6 vs. 89-97; X= 93.7), con menor número de escamas gulares (21-31; X= 25.8 vs. 31-37; X= 33.3), por presentar mayor longitud hocico-cloaca (max. LHC 79.7 mm vs 66.7 mm) (Tabla 2), con mayor relación entre el largo hocico - cloaca y largo de la cola, con mayor relación entre el largo hocico - cloaca y largo del brazo y menor relación entre el largo y ancho del oído (Tabla 3).
Se diferencia de Liolaemus pseudoanomalus por presentar un patrón dorsal de coloración claramente diferente (Fig. 12), con mancha en "V" invertida desde el ojo a las supralabiales ausente o muy difusa (muy evidentes en L. pseudoanomalus), con manchas paravertebrales menos marcadas y evidentes, más pequeñas, en forma de punto o circulo, nunca transversales al cuerpo, con el color del cuerpo castaño o gris claro, nunca gris oscuro, rosado o rojizo, presencia de escamas blancas y/o celestes en los lados del cuerpo y cola (ausentes en L. pseudoanomalus), con ausencia de manchas evidentes en los lados del cuerpo y anillos oscuros en el antebrazo y brazo (presentes en L. pseudoanomalus), con menor número de anillos en la cola (4-5 incompletos vs. 10-11 completos), con bultos en los extremos del pliegue supraescapular ausentes o menos marcado que en L. pseudoanomalus (Tabla 1), por tener menor número de escamas alrededor del cuerpo (61-77; X= 68.6 vs. 69-90; X= 81.9), con menor número de escamas dorsales desde el occipucio a miembros posteriores (76-83; X= 79.6 vs. 83-99; X= 92.3), menor número de escamas gulares (21-31; X= 25.8 vs. 29-40; X= 35.1), mayor longitud hocico-cloaca (max. LHC 79.7 mm vs. 66.7 mm) (Tabla 2) y con mayor relación entre el largo hocico - cloaca y largo de la cola (Tabla 3).
Descripción del holotipo (FML 19225) (Fig. 14)
Macho adulto. Longitud hocico-cloaca 79.7 mm. La cabeza es 1,2 veces más larga (17.4 mm) que ancha (14.8 mm). Altura de la cabeza 10.1 mm. Ancho del cuello 11.1 mm. Interórbitas 9.3 mm. Diámetro longitudinal del ojo 4.7 mm. Longitud oído-ojo 6.5 mm. Alto del oído 2.6 mm y ancho 1,5 mm. Distancia del ojo al labio superior 2.1 mm y distancia entre las narinas 2.4 mm. Longitud de la subocular 4,2 mm. Longitud del torso 41.8 mm y de la cola 86.5 mm. Ancho del cuerpo 18.8 mm. Longitud del muslo 14.0 mm. Longitud de la tibia 11.5 mm. Longitud del pie 18,8 mm. Longitud del húmero 8.1 mm, antebrazo 10.0 mm y mano 10.5 mm. Longitud de la región pigal 7,1 mm y ancho de cloaca 8,1 mm.
Superficie dorsal de la cabeza lisa, con quince escamas dorsales. Rostral más ancha que alta, en contacto con seis escamas. Mental de mayor tamaño que la rostral, de forma trapezoidal, en contacto con cuatro escamas. Sin contacto entre la nasal y rostral, separadas por una escama. Dos internasales más altas que anchas. Con seis escamas rodeando la nasal que está separada de la canthal por dos escamas. Hay ocho escamas entre la rostral y frontal. Frontal dividida en cuatro. Con dos postrostrales. Parietal mayor que interparietal y en contacto con siete escamas. Preocular separada de las loreolabiales por una escama. Cuatro superciliares y doce ciliares superiores. Borde anterior del oído sin escamas auriculares diferenciadas; borde superior sin escama diferenciada. Con nueve temporales, que carecen de quillas. Subocular en contacto con cuatro loreolabiales, sin contactar con la cuarta supralabial. Cuatro supraoculares. Diez supralabiales. Siete loreolabiales. Con cinco infralabiales, la segunda en contacto con dos escamas ventralmente. Cinco escamas escudos. Segundos escudos están separados por dos escamas.
Con 64 escamas alrededor del cuerpo. Con 80 escamas dorsales desde el occipucio hasta el miembro posterior, con el extremo posterior redondeado, yuxtapuestas y sin quillas. Con 23 hileras de escamas en el dorso. Con 73 escamas ventrales (desde mental hasta la cloaca, siguiendo la línea media ventral del cuerpo) más grandes que las dorsales, yuxtapuesto y laminares. Con 21 escamas gulares, yuxtapuesta y sin quillas. Sin pliegue gular. Con 25 escamas en el cuello, desde el borde posterior del oído hasta el hombro, por el pliegue longitudinal. Cuatro poros precloacales. Escamas antehumerales granulares, yuxtapuesta y bien diferenciadas del resto. Pliegue auricular, antehumeral y longitudinal marcados. Escamas del pliegue longitudinal lisas y sin quilla. Cuarto dedo de la mano con 17 escamas y del pie con 22. Con parche femoral. Escamas dorsales de la cola sin mucrón y con quilla, ventrales lisas. Infracarpales laminares, imbricadas y trífidas. Infratarsales laminares que terminan en un pequeño mucrón, imbricadas y lisas. Con cabeza alargada y cuello comprimido (o marcada estrangulación). Ausencia de pliegue postescapular, bulbo anterior y posterior en el pliegue postescapular poco desarrollado o ausente. Escamas del parche con bordes redondeados sin muesca. Escamas del centro de pie simples (con quilla y mucrón).
Coloración en vida (Fig. 14)
Dorso y lados de la cabeza de color castaño claro con numerosas escamas y pequeñas manchas castaño oscuro, escasas escamas blancas y celestes. Infralabiales con alternancia de colores claros y oscuros, supralabiales de color homogéneo (Fig. 3-1B). Mancha en "V" invertida desde el ojo hasta supralabiales ausente o solo presente la línea posterior, difusa (Fig. 3-1C). Cuerpo con 7 pares de manchas paraverterbrales tenues, en forma de puntos o círculos pequeños. Con mancha blanca posterior a cada mancha paravertebral, en forma de círculo, difusa o ausente (Fig. 12). Cuerpo de color castaño claro, con numerosas escamas de color celeste y blanco. Carece de línea vertebral, manchas laterales y manchas dorsolaterales. Sin manchas postescapular. Las escamas del dorso del cuerpo con un patrón de manchado atigrado homogéneo. Los miembros son de color castaño claro. Brazo y antebrazo sin anillos, con 3 anillos difusos o ausentes en el 4° dedo de la mano (Fig. 3-2C). Con 5 anillos en el 4° dedo del pie. La cola de color castaño claro, con numerosas manchas y escamas blancas y celestes. Cola con 5 o 6 pseudoanillos o anillos incompletos.
Ventralmente la región mandibular, miembros anteriores, cloaca y cola de color blanco. Región gular, abdominal y fémur de color amarillo tenue (Fig. 14). Mandíbula y región gular con manchas muy difusas (Fig. 14).
Variación en Liolaemus pipanaco
Basada en seis ejemplares. La superficie dorsal de la cabeza es lisa. Nasal rodeada de 6-7 (X= 6.4; DS= 0.5) escamas. Supralabiales 6-7 (X= 6.8; DS= 0.4), 7-9 (X= 8.0; DS= 0.7) loreolabiales dispuestas en una sola hilera. Supraoculares 4-6 (X= 4.6; DS= 0.8). Parietales de mayor tamaño que las interparietales, rodeada de 5-9 (X= 6.2; DS= 1,8) escamas. 15-17 (X= 16.0; DS= 0.7) escamas entre la rostral y el occipucio. Infralabiales 5-6 (X= 5.8; DS= 0.4). Gulares 21-31 (X= 25.8; DS= 4.4). Temporales 9-14 (X= 10.6; DS= 2.7) sin quillas. Oído siempre más alto (X= 2.4 mm; DS= 0.6) que ancho (X= 1,3 mm; DS= 0,3). Pliegues del cuello desarrollados. Cabeza más larga (X= 15.0 mm; DS= 2.1) que ancha (X= 13.0 mm; DS= 1.5), y alta (X= 8.9 mm; DS= 1.5). Longitud del tronco (X= 31.9 mm; DS= 7.5). Longitud hocico - cloaca en hembras (X= 59.4 mm; DS= 10.3). Longitud del brazo (X= 7.9 mm; DS= 1.4). Longitud del antebrazo (X= 8.5 mm; DS= 1.0). Longitud de la mano (X= 9.1 mm; DS= 1.2). Longitud del muslo (X= 11.9 mm; DS= 1.9). Número de escamas alrededor del cuerpo de 61-77 (X= 68.6; DS= 6.7). Escamas dorsales entre occipucio y muslos 76-83 (X= 79.6; DS= 2.7). Escamas dorsales con borde redondeado, sin quillas y yuxtapuestas. Cuarto dedo de la mano con 13-17 (X= 15.4; DS= 1.8) laminillas digitales. Cuarto dedo del pie con 21-23 (X= 22.2; DS= 0.84) laminillas digitales. Infracarpales con escamas laminares, imbricadas, trífidas. Infratarsales con escamas laminares, imbricadas y lisas. Parte posterior del muslo con parche femoral. Ventrales más grandes que las dorsales 72-86 (X= 78.8; DS= 6.5). Cola de mayor longitud (X= 75.2mm; DS= 8.1) que la longitud hocico-cloaca. Hembras con 1-3 (X= 2.5; DS= 1.0) poros precloacales. Con cabeza alargada y cuello comprimido (o marcada estrangulación). Ausencia de pliegue postescapular, bulbo anterior y posterior en el pliegue postescapular poco desarrollado o ausente (Fig. 2 A y B). Escamas del parche con bordes redondeados sin muesca (Fig. 3-3D). Escamas trífidas del centro de pie.
Variación de coloración en la especie (Fig. 15): Sin dicromatismo sexual evidente. El color de fondo de la cabeza es de color castaño. Con numerosas escamas blancas y sin escamas celestes. En los lados de la cabeza, con manchas alternas, claras y oscuras, en las escamas infralabiales, mientras que las supralabiales son de color uniforme (Fig. 3-1C). Mancha en V invertida desde el ojo hasta supralabiales ausente o solo presente la línea posterior, pero difusa (Fig. 3-1C). Cuerpo de color gris o castaño claro. Las manchas paravertebrales en forma de punto o subcuadrangulares, de color castaño oscuro. Con mancha posterior a cada mancha paravertebral de color blanco, en forma de punto. Numerosas escamas blancas y celestes concentradas sobre las manchas paravertebrales y en los laterales. Manchas laterales ausentes o difusas. Sin mancha postescapular. Sin arco negro antehumeral, línea vertebral y bandas dorsolaterales. Las escamas del dorso del cuerpo con un patrón de manchado atigrado homogéneo (Fig. 3-1). Los miembros varían del color gris al castaño. Sin anillos en brazo y antebrazo, 3 anillos difusos o ausentes en el 4° dedo de la mano (Fig. 3-2C). Con 5 anillo en el cuarto dedo del pie. La cola de color castaño claro, con numerosas escamas blancas y celestes. Cola con 4-5 pseudoanillos o anillos incompletos.
Ventralmente la región mandibular, miembros, cloaca y cola de color blanco. Región gular y abdominal de color amarillo tenue. Mandíbula y región gular con manchas muy difusas.
Distribución (Fig. 8)
Solo es conocida para su localidad tipo, probablemente se distribuya a lo largo de todo el Salar de Pipanaco, llegando hasta la provincia de La Rioja en el extremo sur del salar.
Etimología
El nombre de esta especie es asignado por el lugar en donde habita, el Salar de Pipanaco, extenso salar que se ubica al oeste del cordón montañoso del Ambato Andalgalá, Catamarca.
Historia Natural
Liolaemus pipanaco habita en un área extremadamente árida. El suelo esta compuesto por sal y arena con una importante cantidad de "mica" (silicatos de aluminio), que refracta considerablemente la luz del sol. En este ambiente, hay pocos arbustos y las temperaturas son muy elevadas. Los ejemplares de esta especie, fueron encontrados entre las 10 y 13:00 hs. bajo un sol abrasador. En varias partes de este salar se forman grandes "costras" de arena y sal formando refugios para los lagartos que se cobijan bajo estas. No se conocen más datos sobre su biología y comportamiento.
Liolaemus millcayac sp nov.
1986 Liolaemus anomalus. Cei, J. Monografie IV: 527 pp.
1998 Liolaemus anomalus. Avila, L., Acosta, J.C. y Murúa, A. Cuad. Herp. 12 (1):11-29.
2003 Liolaemus pseudoanomalus. Avila, L., Morando, M. y Belver, L. Herp. Soc. 38(3):52-53.
Holotipo.- FML 3339. Macho adulto. A un Km de la ruta nacional 40, desvio hacia las Lagunas del Rosario, Departamento de Lavalle, Provincia de Mendoza, Argentina. Col. C. S. Abdala, J. L. Acosta. A. Laspiur y R. Semhan. Diciembre de 2010.
Paratipos.- FML 33338, 840-841. 3 ejemplares. Mismos datos que holotipo.
FML 22434-436. 3 ejemplares. 46 Km al noroeste de El Encón, Ruta Nacional 141, Departamento 25 de Mayo, Provincia de San Juan, Argentina. Col. C. S. Abdala, A. Laspiur, J. Nori y G. Scrocchi. Octubre de 2008.
MLP S. 088-90. 2 ejemplares. Alto Pencoso, Departamento de Belgrano, Provincia de San Luis, Argentina.

Figura 15. Ejemplares adultos de Liolaemus millcayac sp. nov. A y B: Holotipo (macho) de un Km de la RN 40, desvío hacia las Lagunas del Rosario o Guanacache, Lavalle, Mendoza, Argentina. C, D y E: Paratipos de 46 Km. al noroeste de El Encón, RN 141, 25 de Mayo. San Juan, Argentina. C y D: machos, E: hembra.
Diagnosis
Liolaemus millcayac sp. nov. pertenece al grupo de Liolaemus boulengeri (Abdala, 2007a) por presentar un parche de escamas agrandadas en la cara posterior del muslo (Etheridge, 1995; Abdala, 2007a; Abdala et al., 2006). Dentro del grupo de L. boulengeri se diferencia de las especies del grupo de L. wiegmannii (Etheridge, 2000) (L. arambarensis, L. azarai, L. cranwelli, L. cuyumhue, L. lutzae, L. multimaculatus, L. occipitalis, L. rabinoi, L. riojanus, L. salinicola, L. scapularis, L. wiegmannii) por tener una hilera de escamas loreolabiales (nunca dos o tres), cuatro escamas en contacto con la mental (nunca seis), fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola. Se diferencia de las especies del grupo de L. darwinii (Abdala, 2007a) (L. abaucan L. albiceps, L. calchaqui, L. chacoensis, L. cinereus, L. crepuscularis, L. darwinii, L. diaguita, L. espinozai, L. grosseorum, L. irregularis, L. koslowskyi, L. laurenti, L. lavillai, L. montanezi, L. olongasta, L. ornatus, L. quilmes, L. uspallatensis) por tener cabeza casi tan larga como ancha, "fleje" o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola, también se diferencia por carecer de dicromatismo sexual evidente y línea negra transversal al ojo. Se diferencia de las especies del grupo de L. melanops (Abdala, 2007a) (L. boulengeri, L canqueli, L. casamiquelai, L. chehuachekenk, L. cuyanus, L. donosobarrosi, L. fitzingerii, L. goestchi, L. hermannunezi, L. inacayali, L. josei, L. mapuche, L. martorii, L. melanops, L. morenoi, L. loboi, L. puelche, L. sagei, L. rothi, L. senguer, L. tehuelche, L. telsen, L. xanthoviridis) por tener dientes posteriores con coronas de bordes rectos, cabeza casi tan larga como ancha, fleje o "peine" palpebral desarrollado, machos con menor número de poros precloacales y menor relación entre el largo hocico-cloaca y longitud de la cola.
Dentro del clado de Liolaemus anomalus, L. millcayac se diferencia de L. acostai por presentar un patrón de coloración dorsal claramente diferente (Fig. 12), con manchas paravertebrales más pequeñas, con presencia de manchas dorsolaterales (ausentes en L. acostai), dorso del cuerpo y manchas paravertebrales con escamas celestes y lados del cuerpo con mayor número de escamas celestes, cola nunca anillada como en L. pipanaco (Tabla 1), menor número de escamas alrededor del cuerpo (58-75; X= 69.7 vs. 66-86; X= 69.7) (Tabla 2).
Se diferencia de Liolaemus anomalus presentar un patrón de coloración dorsal claramente diferente (Fig. 12), con ausencia de mancha en "V" invertida desde el ojo a las supralabiales (presente en L. anomalus), sin presencia de anillos marcados en el antebrazo, brazo y cola, presencia de mancha postescapular grande, mucho más evidente que en los ejemplares de L. anomalus que presentan mancha postescapular rojiza, con presencia de manchas dorsolaterales (ausentes en L. anomalus), dorso del cuerpo y manchas paravertebrales con escamas celestes y lados del cuerpo con mayor número de escamas celestes (Tabla 1), por tener mayor longitud hocico-cloaca (max LHC 78.4 mm vs. 68.6 mm) (Tabla 2) y mayor relación entre el largo hocico - cloaca y la longitud del antebrazo (Tabla 3).
Se diferencia de Liolaemus ditadai por presentar un patrón de coloración dorsal claramente diferente (Fig. 12), con ausencia de mancha en "V" invertida desde el ojo a las supralabiales (presente en L. ditadai), manchas paravertebrales más pequeñas, con bultos en los extremos del pliegue supraescapular (ausentes en L. ditadai) (Tabla 1), menor número de escamas del cuello (20-26; X= 24.5 vs. 29-34; X= 31.0) (Tabla 2), menor relación entre el largo y ancho de la cabeza , menor relación entre el largo y alto de la cabeza, menor longitud hocico-cloaca (max LHC 78.4 mm vs. 92.6 mm) (Tabla 2).
Se diferencia de Liolaemus lentus por presentar un patrón de coloración dorsal claramente diferente (Fig. 12), manchas paravertebrales más pequeñas, con presencia de manchas dorsolaterales (ausentes en L. lentus), manchas paravertebrales, dorso y lados del cuerpo y la cola con escamas celestes (ausentes en L. lentus), con bultos en los extremos del pliegue supraescapular (ausentes en L. lentus), cola nunca anillada en su base como en L. lentus (Tabla 1), por tener menor número de escamas alrededor del cuerpo (58-75; X= 69.7 vs. 73-81; X= 78.2) (Tabla 2), menor relación entre el largo hocico - cloaca y ancho del cuerpo, mayor relación entre el largo del muslo y largo hocico - cloaca, menor relación entre el alto y ancho del oído (Tabla 3), mayor longitud hocico-cloaca (max LHC 78.4 mm vs. 66.8 mm) (Tabla 2).
Se diferencia de Liolaemus pipanaco por tener un patrón de coloración dorsal claramente diferente (Fig. 12), con manchas paravertebrales más marcadas, de mayor tamaño, con presencia de manchas escapulares y manchas dorsolaterales, con presencia de pliegue supraescapular, cola sin anillos en L. millcayac, sin bultos o poco marcados en los extremos del pliegue supraescapular presentes en L. millcayac (Tabla 1), por tener mayor número de escamas dorsales desde el occipucio a miembros posteriores (80-98; X= 90.2 vs. 76-83; X= 79.6) y mayor número de escamas gulares (31-36; X= 34.5 vs. 21-31; X= 25.8) (Tabla 2).
Se diferencia de Liolaemus pseudoanomalus por presentar un patrón de coloración dorsal claramente diferente (Fig. 12), con ausencia de mancha en "V" invertida desde el ojo a las supralabiales (muy evidentes en L. pseudoanomalus), manchas paravertebrales más pequeñas, con presencia de manchas dorsolaterales (ausentes en L. pseudoanomalus), sin presencia de anillos marcados en el antebrazo, brazo y cola, presencia de mancha postescapular grande (ausente en L. pseudoanomalus), manchas paravertebrales, dorso y lados del cuerpo y la cola con escamas celestes (ausentes en L. pseudoanomalus) (Tabla 1), por tener menor número de escamas alrededor del cuerpo (58-75; X= 69.7 vs. 69-90; X= 81.9), mayor longitud hocico-cloaca (max LHC 78.4 mm vs. 66.7 mm) (Tabla 2).
Descripción del holotipo (FML 3339) (Fig. 15A y B)
Macho adulto, largo hocico - cloaca 74.71 mm. Largo del cuerpo 39.97 mm. Cabeza más larga (17.8 mm) que ancha (15.8 mm). Alto de la cabeza 10.5 mm. Diámetro del ojo 6.8 mm. Distancia interorbital 9.8 mm. Distancia entre el meato auditivo y la orbita del ojo 6.7 mm. Alto del meato auditivo: 2.6 mm, ancho: 1.0 mm. Distancia entre la orbita y comisura de la boca 2.0 mm. Distancia entre internarinas 2.4 mm. Subocular 5.9 mm. Largo del muslo 13.0 mm, tibia 10.9 mm, y pie 16.6 mm. Largo del antebrazo 10.6 mm., del brazo 8.9 mm., de la mano 9.1 mm. Largo de la cola 80.5 mm, base de la cola 11.0 mm.
Superficie dorsal de la cabeza lisa, con catorce escamas dorsales. Rostral más ancha que alta, en contacto con seis escamas. Mental de mayor tamaño que la rostral, de forma trapezoidal, en contacto con cuatro escamas. Sin contacto entre la nasal y rostral, separadas por una escama. Tres internasales más altas que anchas. Con siete escamas rodeando la nasal que está separada de la canthal por dos escamas. Hay cuatro escamas entre la rostral y frontal. Frontal dividida en más de 5 escamas. Con tres postrostrales. Parietal mayor que interparietal y en contacto con seis escamas. Preocular separada de las loreolabiales por una escama. Cuatro superciliares y doce ciliares superiores. Borde anterior del oído con dos escamas auriculares diferenciadas, borde superior con tres escamas diferenciadas. Con diez temporales, que carecen de quillas. Subocular en contacto con cinco loreolabiales, sin contactar con la cuarta supralabial. Seis supraoculares. Siete supralabiales. Nueve loreolabiales. Con siete infralabiales, la segunda en contacto con dos escamas ventralmente. Seis escamas escudos. Segundos escudos están separados por dos escamas.
Con 74 escamas alrededor del cuerpo. Con 85 escamas dorsales desde el occipucio hasta el miembro posterior, con el extremo posterior redondeado, yuxtapuestas y sin quilladas. Con 25 hileras de escamas en el dorso. Con 84 escamas ventrales (desde mental hasta la cloaca, siguiendo la línea media ventral del cuerpo) más grandes que las dorsales, yuxtapuesto y laminares. Con 35 escamas gulares, yuxtapuesta y sin quillas. Sin pliegue gular. Con 25 escamas en el cuello, desde el borde posterior del oído hasta el hombro, por el pliegue longitudinal. Dos poros precloacales. Escamas antehumerales granulares, yuxtapuesta y bien diferenciadas del resto. Pliegue auricular, antehumeral y longitudinal marcados. Escamas del pliegue longitudinal lisas y sin quilla. Cuarto dedo de la mano con 17 escamas y del pie con 24. Con parche femoral. Escamas dorsales de la cola sin mucrón y con quilla, ventrales lisas. Infracarpales laminares, imbricadas y trífidas. Infratarsales laminares que terminan en un pequeño mucrón, imbricadas y lisas. Pliegue postescapular presente, bulbo anterior y posterior en el pliegue postescapular desarrollado (Fig. 2 A y B). Escamas del parche con bordes redondeados sin muesca (Fig. 3-3D). Escamas del centro de pie simples (con quilla y mucrón).
Coloración en vida (Fig. 15A y 15B)
Dorso y lados de la cabeza de color castaño claro. Con numerosas escamas y pequeñas manchas de color blanco y castaño oscuro, distribuidas de manera irregular en toda la cabeza. Con algunas escamas amarillas en los lados de la cabeza. Subocular de color amarillo. Infralabiales con manchas alternas de color blanco y castaño claro, supralabiales de color uniforme (Fig. 3-1A). Región temporal sin escamas celestes. Mancha en "V" invertida desde el ojo a las supralabiales ausente.
Tronco con 6 pares de manchas paraverterbrales con forma de punto o subcuadrangular, de color castaño oscuro, con mancha blanca posterior en forma de círculo (Fig. 12, 15). El color del torso y lados del cuerpo, es castaño claro, salpicado con numerosas escamas de color blanco. Con escamas celestes concentradas en las manchas paravertebrales y algunas dispersas en el dorso y lados del cuerpo. Carece de línea vertebral. Sin bandas dorsolaterales. Las escamas del dorso del cuerpo con un patrón de manchado atigrado homogéneo. Sin manchas en los lados del cuerpo. Con mancha postescapular evidente, de color negro. Los miembros anteriores y posteriores son de color castaño claro, ambos presentan numerosas escamas de color blanco y pocas escamas celestes y amarillas. Sin anillos oscuros en el brazo y antebrazo (Fig. 3-2C). El 4° dedo de la mano con 3 anillos difusos o ausentes. Cuarto dedo del pie con 4 anillo difusos. La cola dorsal y lateralmente, de color castaño claro, con escamas y manchas de color blanco. En los lados de la cola hay numerosas escamas de color celeste. Cola sin anillos (Fig. 15).
Ventralmente la región mandibular, miembros anteriores, cloaca y cola, de color blanco. La región gular, abdominal y del muslo con manchas amarillas (Fig. 15). Desde las infralabiales salen manchas oscuras formando en la región gular un collar de puntos, las manchas son difusas en el pecho (Fig. 12). En la región laterales del abdomen con escamas de color naranja (Fig. 15).
Variación en Liolaemus millcayac
Basada en dieciséis ejemplares (ocho machos y siete hembras). La superficie dorsal de la cabeza es lisa. Nasal rodeada de 6-8 (X= 7.0; DS= 0,5) escamas. Supralabiales 7-8 (X= 7.2; DS= 0.4), 7-9 (X= 8,2; DS= 0.6) loreolabiales dispuestas en una sola hilera. Supraoculares 4-6 (X= 5,6; DS= 0,7). Parietales de mayor tamaño que las interparietales, rodeada de 5-8 (X= 6.3; DS= 0,8) escamas. 11-15 (X= 13.5; DS= 1.3) escamas entre la rostral y el occipucio. Infralabiales 6-8 (X= 7.1; DS= 0,6). Gulares 31-36 (X= 34.7; DS= 1,6). Temporales 10-15 (X= 11.7; DS= 1.5) sin quillas. Oído siempre más alto (X= 2,3 mm; DS= 0,5) que ancho (X= 1,1 mm; DS= 0,2). Pliegues del cuello desarrollados. Cabeza más larga (X= 15.6 mm; DS= 3.2) que ancha (X= 14.1mm; DS= 2.7), y alta (X= 9.1 mm; DS= 1.6). Longitud del tronco (X= 36.6 mm; DS= 8.1). Longitud hocico - cloaca en machos (X= 74.5 mm; DS= 2.5) y en hembras (X= 53.2 mm; DS= 14.3). Longitud del brazo (X= 7.6 mm; DS= 1.8). Longitud del antebrazo (X= 9.0 mm; DS= 1,8). Longitud de la mano (X= 8.8 mm; DS= 1.5). Longitud del muslo (X= 11.6 mm; DS= 2.5). Número de escamas alrededor del cuerpo de 58-75 (X= 69.2; DS= 5.2). Escamas dorsales entre occipucio y muslos 80-98 (X= 89.4; DS= 5.7). Escamas dorsales con borde redondeado, sin quillas y yuxtapuestas. Cuarto dedo de la mano con 7-18 (X= 14.5; DS= 3.2) laminillas digitales. Cuarto dedo del pie con 20-24 (X= 22.1; DS= 1,7) laminillas digitales. Infracarpales con escamas laminares, imbricadas, trífidas. Infratarsales con escamas laminares, imbricadas y lisas. Parte posterior del muslo con parche femoral. Ventrales más grandes que las dorsales 75-92 (X= 81.2; DS= 4.6). Cola de mayor longitud (X= 59.9 mm; DS= 22.8) que la longitud hocico-cloaca. Machos con 3-4 (X= 3.5; DS= 0.7) poros precloacales, 4 de 4 hembras con 2-3 (X= 2.6; DS= 0.5) poros precloacales. Con cabeza alargada y cuello comprimido (o marcada estrangulación).Con cabeza alargada y cuello comprimido (o marcada estrangulación). Pliegue postescapular presente, bulbo anterior y posterior en el pliegue postescapular desarrollado (Fig. 2 A y B). Escamas del parche con bordes redondeados sin muesca (Fig. 3-3D). Escamas del centro de pie simples (con quilla y mucrón).
Variación de coloración en la especie (Fig. 15C a 15E): Sin dicromatismo sexual evidente. La cabeza presenta un color de fondo castaño. Con escasas escamas blancas y sin escamas celestes. Infralabiales con manchas alternas de color blanco y castaño claro, supralabiales de color uniforme (Fig. 3-1B, 15). Mancha en V invertida desde el ojo hasta supralabiales ausente (Fig. 3-1C, 15). Cuerpo de color gris o castaño. Las manchas paravertebrales en forma de punto o subcuadrangulares, de color castaño oscuro. Con mancha blanca posterior a cada mancha paravertebral, pequeña, en forma de punto. Numerosas escamas celestes concentradas sobre las manchas paravertebrales y en los lados del cuerpo. Con mancha postescapular evidentes de color negro, en ocasiones rodeada por escamas naranjas (Fig. 15). Con manchas dorsolaterales. Sin arco negro antehumeral, línea vertebral, bandas dorsolaterales y manchas en los lados del cuerpo. Las escamas del dorso del cuerpo con un patrón de manchado atigrado homogéneo. Los miembros varían del gris al castaño. Sin anillos en brazo y antebrazo, 3 anillos difusos o ausentes en el 4° dedo de la mano (Fig. 3-2C). Cuarto dedo del pie con 4 anillo difusos. Cola de color castaño claro, con escamas y manchas de color blanco y con numerosas escamas de color celeste. Cola sin anillos oscuros.
Ventralmente la región mandibular, miembros anteriores, cloaca y cola de color blanco. La región gular, abdominal y del muslo, con manchas amarillas. Mandíbula con manchas oscuras formando en la región gular un collar de puntos, las manchas son difusas en el pecho. En los lados del abdomen con numerosas escamas de color naranja (Fig. 36).
Distribución (Fig. 8)
Esta especie esta distribuida en el norte de Mendoza (departamento de Lavalle), sur de la provincia de San Juan, desde aproximadamente 50 km al oeste del Encón hasta límite con provincia de La Rioja, en Macasín. Tambien se distribuye en Alto Pencoso y Sierra del Gigante al noroeste de la provincia de San Luis.
Etimología
Esta especie esta dedicada a los "Huarpes Millcayac", uno de los pueblos originarios que habitaron la zona del norte de Mendoza y San Luis así como el Sur de San Juan donde se distribuye esta especie. Los Huarpes eran muy altos y delgados, vivian de manera sedentaria dedicados a la cestería. En la actualidad viven en la zona de las desecadas Lagunas de Guanacache , comunidades criollas que pueden acreditar su directa filiación huarpe; el senso nacional argentino del 2010, arrojo como resultado que actualmente descienden en primera generación del pueblo huarpe 12.710 personas en las provincias de Mendoza, San Juan y San Luis, de los cuales 2.610 residen en comunidades.
Historia Natural
Cei (1986), describe a Liolaemus millcayac como una población de L. anomalus, y menciona que habita en un ambientes desérticos y halófilos de Atriplex, Prosopis strombulifera, etc., los cuales están expuestos a lluvias de verano. Se caracterizan por hacer cuevas de 20-25 cm por encima de montículos que se forman alrededor de las matas. Son lagartos muy resistentes a altas temperaturas (45°C). A partir de septiembre se inician los cortejos sobre las dunas, destacándose el macho por los vistosos y rápidos rodeos alrededor de la hembra. La misma coloca entre 7 y 8 huevos, en una sola postura. Son insectívoros, con alto porcentaje de hormigas y se caracterizan por ser muy agresivo.
Análisis estadístico Liolaemus anomalus, L. pseudoanomalus y L. acostai
Los resultados con el análisis de MANOVA, nos indican que los 8 caracteres seleccionados de relación, en forma conjunta estarían explicando un 37% de la variación en las especies (Fig. 16).
Figura 16. Grafico que exhibe los resultados realizados con el análisis de MANOVA, entre L. pseudoanomalus, L. anomalus y L. acostai sp. nov., indicando el porcentaje de variación en las especies (eje x) con los caracteres dependientes seleccionados (eje y). Wilks Lambda = 0.37748; F (16/64)= 3.2951; p= 0.00019. Las barras verticales indican los intervalos de confianza 0.95. Referencias: (-o-): Relación entre el largo/ancho de la cabeza. (---?---): Relación entre el ancho/largo del torso. (.?.): Relación entre el largo del cuerpo y el Largo Hocico-Cloaca. (-?-): Relación entre el largo del muslo y el largo del torso. (-.-): Relación entre el largo/ancho del meato auditivo. (): Relación entre el alto/largo de la cabeza. (-.-): Relación entre el largo de la mano y el Largo Hocico-Cloaca. (---?---): Relación entre el alto de la cabeza y el alto del meato auditivo.
El análisis univariado aportado por el MANOVA, mostró que de los 8 caracteres de relaciones seleccionados, solo 3 fueron significativos al momento de discriminar especies (Fig. 17).
Figura 17. Análisis univariado de MANOVA exhibiendo los tres caracteres que fueron significativos al momento de discriminar especies entre L. acostai, L. pseudoanomalus y L. anomalus. A: Relación entre el alto y el ancho del oido, F(16, 84)= 8,09, p< 0.001. B: Relación entre el alto y el largo del torso, F(16, 84) = 6,94, p< 0.001. C: Relación entre el ancho del cuerpo y LHC, F(16, 84)=14,93, p< 0.001. Las diferentes letras en cada gráfico indican las diferencias significativas entre los contrastes. Las barras verticales indican los intervalos de confianza 0,95.
Los caracteres referidos a las relaciones entre: largo y ancho de la cabeza, longitud del fémur y largo del torso, alto/largo de la cabeza, longitud de la mano/ LHC y alto de la cabeza/alto del oído, no presentaron diferencias significativas entre las especies.
Las variables significativas fueron las siguientes:
La relación entre el alto y el ancho del oído es una variable significativa (P=0.001) y separa Liolaemus anomalus de Liolaemus acostai y Liolaemus pseudoanomalus (Fig. 17A).
La relación entre el ancho y el largo del torso es una variable significativa (P=0.001) y separa Liolaemus pseudoanomalus de Liolaemus anomalus (Fig. 17B).
La relación entre el ancho del cuerpo y el LHC es una variable significativa (P=0.001) y separa Liolaemus pseudoanomalus de Liolaemus anomalus y Liolameus acostai (Fig. 17C).
Análisis Cladístico
Para analizar las relaciones dentro del grupo de Liolaemus anomalus y su ubicación en el grupo de L. boulengeri se realizaron 21 corridas diferentes, una con pesos iguales (PI) y las otras 20 bajo pesos implicados de Goloboff (1993), con valores de 1 a 20 para la constante K.
En todas las corridas realizadas, el grupo de Liolaemus anomalus resultó monofilético, y en todas las corridas utilizando pesos implicados aparece incluido dentro del grupo de L. chacoensis de Abdala (2007).
En la hipótesis PI (Fig. 18), el grupo de Liolaemus anomalus es grupo hermano del grupo de L. boulengeri, coincidiendo con la hipótesis de Abdala (2007a) que le da identidad al grupo de L. chacoensis (equivalente al grupo de L. boulengeri de Etheridge (1995); y que no incluye a las especies del grupo de L. anomalus).
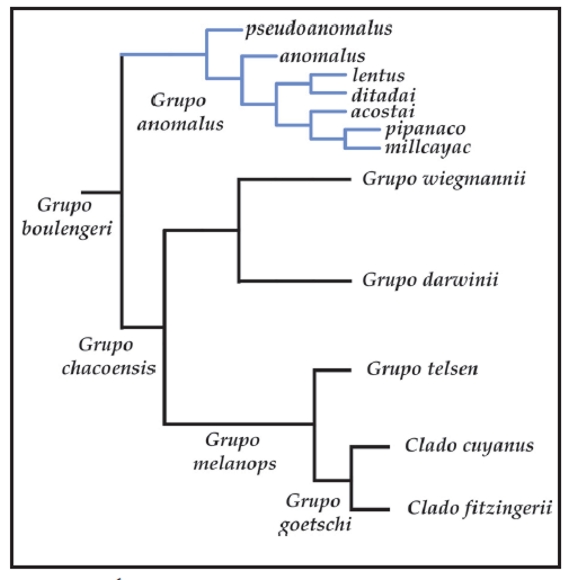
Figura 18. Árbol que exhibe las relaciones filogenéticas del grupo de L. anomalus, tratando los caracteres con pesos iguales (IG). En esta hipótesis filogenética el grupo de L. anomalus es hermano del grupo de L. chacoensis, dentro del grupo de L. boulengeri. Longitud: 1033.660 CI: 0.251 IR: 0.557.
Utilizando pesos implicados el grupo de Liolaemus anomalus siempre se relacionó como grupo hermano del grupo de L. wiegmannii, sostenido por las siguientes sinapomorfías: el número de escamas ventrales, el número de escamas del cuello, el número de escamas infradigitales del cuarto dedo del pie, el número de escamas infradigitales del cuarto dedo de la mano, el número de poros en los machos, la relación entre el largo de la cabeza y largo hocico - cloaca y los machos con mancha postescapular presente.
Utilizando pesos implicados obtuvimos dos hipótesis, una con K= 1 y 2 (Fig. 19) y otra con K= 3 a 20 (Fig. 20). La hipótesis con K= 1 y 2 presenta el clado formado por el grupo de L. anomalus, y el grupo de L. wiegmannii como hermano del grupo de L. darwinii, mientras que en la hipótesis con K= 3 a 20 el grupo de L. darwinii se encuentra más relacionado con el grupo de L. melanops (Abdala, 2007a).
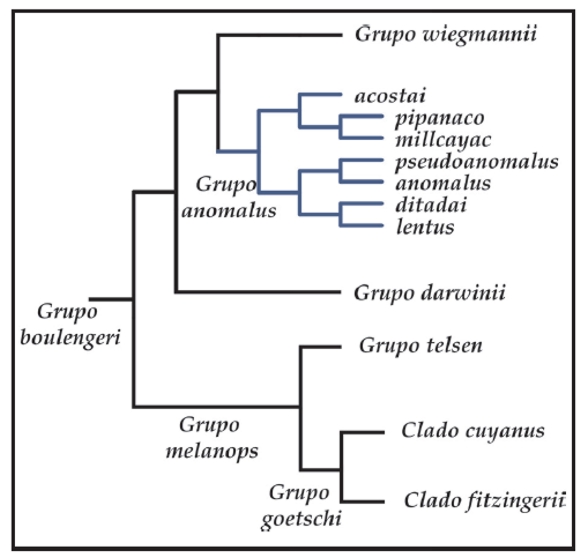
Figura 19. Árbol que exhibe las relaciones filogenéticas del grupo de L. anomalus, tratando los caracteres bajo el método de pesos implicados con K=1 o 2. Esta hipótesis presenta al grupo de L. anomalus como hermano del grupo de L. wiegmannii, no recuperándose los grupos de L. chacoensis y L. laurenti propuestos por Abdala (2007). Longitud: 103.80256 CI: 0.232 IR: 0.510.
Figura 20. Árbol que exhibe las relaciones filogenéticas del grupo de L. anomalus, tratando los caracteres bajo el método de pesos implicados con K=3 a 20. Esta hipótesis también presenta al grupo de L. anomalus como hermano del grupo de L. wiegmannii, no recuperándose los grupos de L. chacoensis y L. laurenti propuestos por Abdala (2007). Longitud: 85.57064 CI: 0.238 IR: 0.525. El clado de L. anomalus muestra los valores obtenidos usando como prueba de soporte el remuestreo simétrico.
De la misma forma las relaciones internas son muy estables para el grupo de Liolaemus anomalus, ya que la relación entre las tres especies que se describen en este trabajo forman el clado (L. acostai (L. millcayac + L. pipanaco)) en todos las topologías obtenidas, independientemente del valor asignado para K, incluso esta relación también se obtiene para PI (pesos iguales). Las sinapomorfías que soportan este clado son: el desarrollo leve del pliegue ante humeral (carácter 51), cuello definido menos ancho que la cabeza (carácter 60), hembras con algunas escamas celestes o blancas en los lados del cuerpo (carácter 121) y de la cola (carácter 122).
Otra relación muy estable dentro del grupo de Liolaemus anomalus es la formada por (L. anomalus + L. pseudoanomalus), la cual es recuperada en todas las corridas que se utilizó pesos implicados. Las sinapomorfías que unen esta relación son el número de escamas dorsales desde el occipucio hasta los miembros posteriores (carácter 12) y el cuello igual o más ancho que la cabeza (carácter 60), las supralabiales e infralabiales con manchas claras y oscuras de manera alternada (carácter 65), anillos en el cuarto dedo de la mano evidentes (carácter 69), machos sin mancha postescapular (carácter 75), hembras con manchas paravertebrales anteriores (carácter 111) y posteriores (carácter 112) en forma de línea transversal al cuerpo o con forma de medialuna con escotadura posterior.
Las diferencias entre las distintas hipótesis obtenidas se exhiben en la ubicación de Liolaemus lentus y L. ditadai, que en la hipótesis con pesos iguales aparecen como especies hermanas formando un clado con (L. acostai (L. millcayac + L. pipanaco)) (Fig. 18). Cuando se le asignó el valor de 1 o 2 a K se formó el clado ((L. lentus + L. ditadai) + (L. anomalus + L. pseudoanomalus)) (Fig. 19), mientras que con K= 3 a 20 el clado entre L. ditadai y L. lentus no se recuperó, formando el clado (L. lentus (L. ditadai (L. pseudoanomalus + L. anomalus) + (L. acostai (L. millcayac + L. pipanaco)))) (Fig. 20).
Por estar presente en la mayor cantidad de árboles obtenidos y mayor número de congruencias con los demás autores, la hipótesis con valores de 3 a 20 para "K" es la hipótesis filogenética que proponemos para el grupo de Liolaemus anomalus (Fig. 20).
En base a la hipótesis filogenética presentada en este trabajo (K= 3 a 20), 26 sinapomorfías soportan la monofilia del grupo de Liolaemus anomalus: el número de escamas supraoculares (carácter 3), el número de escamas supralabiales (carácter 0), el número de escamas alrededor del cuerpo (carácter 8), el número de escamas gulares (carácter 11), el número de escamas dorsales en el cuerpo desde el occipucio hasta los miembros posteriores (carácter 12), el número de poros en los machos (carácter 17), el número de poros en las hembras (carácter 18), la relación entre el ancho de la cabeza y el largo hocico - cloaca (carácter 20), la relación entre el ancho y el largo de la cabeza (carácter 21), la relación entre el largo de la cola y el largo hocico - cloaca (carácter 23), la relación entre el alto y ancho del oído (carácter 31), la relación entre el alto de la cabeza y el alto del oído (carácter 32), la ausencia del proceso tibial en forma de navaja (carácter 37), la ausencia de la escama diagonal superior en el oído (carácter 44), la presencia del pliegue supraescapular (carácter 52), con escamas del dorso del cuerpo sin quilla (carácter 57), con escamas del dorso del cuerpo yuxtapuestas (carácter 58), el diseño de las escamas dorsales con un punteado uniforme y definido (carácter 59), los machos y hembras sin línea oscura entre el ojo y hombro (caracteres 72 y 105), con machos y hembras sin manchas por debajo de la línea media lateral del cuerpo (caracteres 88 y 120), machos y hembras con diseño de la cola anillado (caracteres 99 y 133) y con ausencia de dicromatismo sexual (carácter 141).
DISCUSIÓN
Taxonomía
Debido a las características morfológicas y biología particular que presentan las especies del grupo de Liolaemus anomalus, es entendible que se haya propuesto como un género diferente relacionado a Liolaemus por muchos autores (Müller, 1928; Nuñez y Yañez, 1984; Laurent, 1984). Asimismo la propuesta de Cei (1986, 1993) de designar al grupo de L. anomalus como un subgénero (Ceiolaemus) de Liolaemus es comprensible teniendo en cuenta la metodología y el sistema de clasificación que desarrollaban en aquella época. Sin embargo ninguna de estas propuesta prosperó cuando la metodología cladista comenzó a implementarse para analizar las relaciones filogenéticas de los Liolaemus. Inicialmente fue Etheridge (1995), en un análisis no formal pero con base en las sinapomorfías de los grupos, quien sinonimizó y desestimó los géneros y subgéneros propuestos (Abas, Ceiolaemus, Phrynosaura) para el grupo de L. anomalus. En base a los resultados presentados en este trabajo podemos coincidir con la propuesta de Etheridge (1995), ya que la validación como género o subgénero del grupo de L. anomalus dejaría como parafilético al género Liolaemus. Sin embargo Etheridge (1995) propone al grupo de Liolaemus anomalus basal al grupo signifer o "argentino" por la presencia de la hipertrofia del músculo puboisquitibialis y la ausencia del "parche femoral" (carácter que une a las especies del grupo de L. boulengeri). Esta hipótesis es opuesta a los resultados expuestos en este trabajo, ya que el grupo de L. anomalus es claramente un grupo monofilético anidado dentro del grupo de L. boulengeri y, al igual que Abdala (2007a), afirmamos la presencia del parche femoral en todas las especies relacionadas al grupo de L. anomalus.
Los resultados obtenidos indican, con la descripción de los nuevos taxa, que el grupo de Liolaemus anomalus es más diverso de lo que se conocía en oposición a la hipótesis de Cei (1979, 1986, 1993). Las resoluciones de los problemas taxonómicos planteados en este trabajo, muy complejos en algunos casos (debido a las breves descripciones de especies, sinonimias erróneamente establecidas y la determinación inexacta de la localidad tipo), son congruentes con las hipótesis de Abdala (2007a) que eleva al rango de especie a Liolaemus ditadai y revalida como especie a Liolaemus lentus. Asimismo en contraposición a las hipótesis de Cei (1979, 1986), Avíla et al. (2003) y Abdala (2007a) las poblaciones que habitan en Ischigualasto si bien presentan algunos caracteres particulares son consideradas como L. anomalus. Igualmente consideramos que la población que habita en Colonia Mackinlay, Departamento Rivadavia, provincia de Santiago del Estero, Argentina, y que fuera asignada a Liolaemus ditadai por Abdala (2007b), deberá ser revisada con mayor detalle si se logra capturar más ejemplares a fin de determinar su verdadera identidad, ya que con solo un ejemplar (mal fijado) es difícil establecer su posición taxonómica.
Relaciones filogenéticas
El análisis cladístico desarrollado en este estudio en base a caracteres morfológicos recuperó, en todas las corridas realizadas, al grupo de Liolaemus anomalus dentro del grupo L. boulengeri. Este resultado es congruente con los análisis filogenéticos en base a caracteres moleculares propuestos por Schulte et al. (2000), Morando (2004), Espinoza et al. (2004), por los análisis filogeográficos de Avila et al. (2006, 2008, 2010) y por el análisis de Evidencia Total de Abdala (2007a).
En todos los análisis desarrollados (Figs. 18-20) el grupo de Liolaemus anomalus aparece como grupo hermano del grupo de L. wiegmannii, este punto es incongruente con la hipótesis de Abdala (2007a), en el cual el grupo de L. anomalus es grupo hermano del grupo de L. chacoensis (grupo propuesto en dicho trabajo y que equivale al grupo de L. boulengeri de Etheridge, 1995). De esta manera el grupo de L. chacoensis (Abdala, 2007a) es desestimado por los resultados aquí expuestos, ya que sería parafilético. Asimismo la hipótesis presentada en este trabajo deja como parafilético al grupo de L. laurenti (Abdala, 2007a), conformado por los grupos de L. wiegmannii y de L. darwinii, ya que el grupo de L. wiegmannii se une al grupo de L. anomalus (Fig. 20). Incluso en los análisis realizados con pesos implicados con valor de 3 a 20 para la constante de concavidad (Fig. 20), el grupo de L. darwinii se une como grupo hermano del grupo de L. melanops (Abdala, 2007a) y no con el grupo de L. wiegmannii como propuso Abdala (2007a). Sólo cuando se utiliza el valor de 1 y 2 para "K" (Fig. 19), el clado formado por los grupos de L. anomalus y de L. wiegmannii, se une al grupo de L. darwinii.
A pesar que el clado formado por los grupos de Liolaemus anomalus y de L. wiegmannii son incongruentes con los resultados de Abdala (2007a), se muestran notoriamente congruentes con los análisis filogenéticos moleculares desarrollados por Schulte et al. (2000), Morando (2004), Espinoza et al. (2004), Avila et al. (2006, 2008, 2010) y los análisis de morfología interna de Lobo y Abdala (2002). Esta congruencia en los análisis, con diferentes tipos de caracteres (morfológicos externos, internos y moleculares) y tratados con diferentes criterios de optimalidad (criterio de parsimonia, máxima verosimilitud y método bayesiano), proporcionan peso importante para esta hipótesis de relaciones.
Con respecto a las relaciones internas dentro del grupo de Liolaemus anomalus presentadas en este trabajo, solo pueden ser comparados con los resultados de Abdala (2007a), ya que es el único trabajo previo en el cual se utilizan casi todas las especies válidas para el grupo (no utilizó a L. millcayac). En las dos hipótesis filogeneticas obtenidas en este trabajo se repiten las relaciones formadas por (L. anomalus + L. pseudoanoamlus) y (L. acostai (L. millcayac+ L. pipanaco)), variando la posición de L. lentus y L. ditadai. La única congruencia con la hipótesis de Abdala (2007a) se da en el árbol con K= 1 o 2 con la relación (L. lentus + L. ditadai).
De los resultados obtenidos en los análisis desarrollados en este trabajo se observa que Liolaemus lentus, en las hipótesis propuestas, no está relacionado directamente con L. anomalus, presentando más evidencia a la hipótesis de Abdala (2007a) que invalida la sinonimia planteada por Donoso Barros (1969), de L. lentus con L. anomalus. De la misma manera L. ditadai tampoco aparece filogenéticamente cercana a L. anomalus, por lo tanto también se afianza la propuesta por Abdala (2007a) en la cual se eleva a L. ditadai al rango específico.
Las descripciones de los nuevos taxa desarrollados en este trabajo están firmemente apoyados en el análisis filogenético presentado. Liolaemus acostai, fue mencionada reiteradamente como una población de L. pseudoanomalus (Villavicencio et al., 2001, 2002, 2003, 2006, 2007), y L. millcayac fue considerada una población de L. anomalus (Cei 1986; Avila et al., 1998, 2003), incluso Cei (1979) designó un "neotipo" de L. anomalus correspondiente a un ejemplar de L. millcayac. Asimismo Liolaemus pipanaco fue considerado por Avila et al. (2003) como una población de L. pseudoanomalus. Sin embargo las nuevas especies están filogenéticamente relacionadas, formando entre sí, en todos los árboles obtenidos, el clado (L. acostai (L. millcayac + L. pipanaco)) (Figs. 18-20), posicionandose distantes de aquellas con las que estuvieron confundidas.
Otro punto a destacar es la incongruencia con la propuesta de Espinoza et al. (2004), Pincheira et al. (2008) y Lobo et al. (2010) en la cual consideran a Liolaemus duellmani como integrante del grupo de L. anomalus. Los resultados presentados infieren que L. duellmani es un representante del grupo de L. montanus, coincidiendo con la idea de Cei (1978).
Los análisis estadísticos realizados en este trabajo, proporcionan información valiosa, ya que los 3 caracteres que muestras diferencias significativas entre las especies analizadas, son capaces de diferenciar a L. anomalus y L. pseudoanomalus. En los caracteres utilizados no hay ninguno exclusivo para L. acostai y que permita diferenciarlo de L. anomomalus y L. pseudoanomalus. Esto amerita la elección, para futuros análisis estadísticos, de otros caracteres que puedan llegar a ser específicos de L. acostai.
AGRADECIMIENTOS
Agradecemos a M. Halloy y G. Scrocchi por su lectura crítica y valioso aporte en la elaboración del manuscrito. A J. Abdala, J. C. Acosta, S. Barrionuevo, P. Corrales, A. Laspiur, E. Malovini, J. Nori, C. Pamelin, S. Quinteros, L. Quiroga, E. Sanabria, G. Scrocchi, Romina Semhan, por su invalorable ayuda en el campo. A Pablo Schliserman por su importante colaboración para los análisis estadísticos. A Christian Fulvio Perez por el préstamo de la foto de L. lentus. A Esteban Lavilla, Eduardo Sanabria, Lorena Quiroga, Alejandro Laspiur y Juan Carlos Acosta, por el envío y prestamo de material. Especialmente a Jorge Williams y Hugo Carrizo por el préstamo de ejemplares holotipos y paratipos. A las Direcciones de Fauna de Catamarca, Mendoza, San Juan, La Rioja y La Pampa por permitir realizar los trabajos y colectas de campo. Esta investigación comenzó en el año 2001 y fue posible realizarla gracias a la financiación de los siguientes proyectos: CIUNSA N°915/00, 1236/04, CIUNT Nº G 218, G315, G430, PIP-CONICET N° 2668, 6287, 5982, 5780, 2422 y PICT 01205.
Apéndice I
Liolaemus ditadai (6): MLP 701 (Holotipo). Salinas Grandes, a 4 Km. del límite. Córdoba - Catamarca, Departamento de Tulumba, Provincia de Córdoba. MLP 702-05. (Paratipos), iguales datos que Holotipo. FML 23616; 23624. Salinas Grandes, a 4 Km. del límite Córdoba - Catamarca, Departamento de Tulumba, 29°51'10.7 S 64° 40'09.0" O, Provincia de Córdoba.
Liolaemus lentus (3): MACN 22071 (Holotipo) Cochicó, Altos de Cochicó. Departamento Puelén, Provincia de La Pampa. MACN 22072-73. Iguales datos que Holotipo.
Liolaemus anomalus (17): MLP. S. 785. 786 (Material Tipo). Provincia de La Rioja. MLP 1270. Los Baldecitos, Departamento de Valle Fértil, Provincia de San Juan. MLP 017-018. Chilecito, Departamento de Chilecito, Provincia de La Rioja. MACN 1965. Guallapa, Patquía, Provincia de La Rioja. MACN 12975-76. Ischigualasto, Departamento de Valle Fértil, Provincia de San Juan. MACN 199. Provincia de La Rioja. FML 309. Agua de La Peña, Ischigualasto, Departamento Valle Fertil, Provincia de San Juan. FML 02740. Talampaya, departamento Lavalle, provincia de La Rioja. FML 00318. Guayapa, Departamento Independencia, Provincia de La Rioja. MACN 558. Nonogasta, Provincia de La Rioja. FML 19007-008. Ischigualasto, Departamento de Valle Fértil, Provincia de San Juan. FML 24136- 37. Ischigualasto, departamento Valle Fertil, provincia de San Juan.
Liolaemus pseudoanomalus (38): MACN 1489. Aimogasta, departamento de Arauco, provincia de La Rioja. FML 9447. Acceso al Señor de la Peña, departamento de Castro Barros, provincia de La Rioja. MACN 8496. 8515. Provincia de La Rioja. MLP 992. Provincia de La Rioja. FML 19009. Aimogasta, Departamento Arauco, Provincia de La Rioja. FML 018121-22. Medanitos, Departamento Tinogasta (1650 m), Provincia de Catamarca. FML 19023- 027. Anillaco, Departamento Castro Barros, Provincia de La Rioja. FML 19017-022. Anillaco, Departamento Castro Barros, Provincia de La Rioja. FML 19029-32. Anillaco, departamento Castro Barros, provincia de La Rioja. SAUR 03004. 8.2 km. al norte de Fiambalá, sobre Ruta Provincial 45, Departamento Tinogasta, Provincia de Catamarca. FML 020871-5. Campo de Loma Larga (1100 m), Antinaco, Departamento de Famatina, Provincia de La Rioja. FML 1983. Aimogasta, Provincia de La Rioja. FML 19028. Anillaco, Departamento Castro Barros, Provincia de La Rioja. FML 19010-16. Anillaco, Departamento Castro Barros, Provincia de La Rioja.
Liolaemus acostai (37): FML 690. 20 Km. de Caucete, Departamento de Caucete, Provincia de San Juan. FML 3734. Posta el Balde, Km. 241, Ruta Nacional 40, Departamento Jachal, Provincia de San Juan. FML 24127. Departamento Zonda, Lecho Rio San Juan, Provincia de San Juan. FML 24128. Barreal de Matagusano, Provincia de San Juan. FML 16336-337. La Laja, Departamento Albardón, Provincia de San Juan. FML 18398. La Laja, Departamento Albardón, Provincia de San Juan. FML 24118-26. La Laja, Departamento Albardón, Provincia de San Juan. FML 22437-443. Baños de Salado, La Laja, 20 Km. al norte de San Juan capital, Departamento Albardón, Provincia de San Juan. JV 3-8-13-14-21-22-24-33-40-47-56-58-61-64-67-84-87, 20 Km al noroeste de San Juan, Provincia de San Juan.
Liolaemus pipanaco (7): FML 15011-12. Salar de Pipanaco, Provincia de Catamarca. FML 19225. Salar de Pipanaco, Entrando por puesto Rio Blanco 36 Km al sur de Andalgalá, (737 m.s.n.m). Departamento Andalgalá, Provincia de Catamarca. FML 18396. Pasando por puesto Rio Blanco, al oeste para llegar a Salar de Pipanaco, 27°49'23.9" S 65°14'38.2"O, Departamento Andalgalá, Provincia de Catamarca. FML 1480. Salar de Pipanaco, Departamento Andalgalá, Porvincia de Catamarca.
Lioalemus millcayac (16): FML 22434-436. 46 Km al noroeste de El Encón, Ruta Nacional 141, Departamento 25 de Mayo, Provincia de San Juan. FML 24129-32. Sur de Macasin, Provincia de La Rioja. FML 3338-41. Norte de Mendoza a un Km de la RN 40, desvio hacia las Lagunas del Rosario, Departamento de Lavalle, Provincia de Mendoza. MLP 088-90. Alto Pencoso, Departamento de Belgrano, Provincia de San Luis. MLP 086-87. Sierra del Gigante, Departamento de Belgrano, Provincia de San Luis.
Apéndice II
Liolaemus calchaqui: FML 17013 Hualinchay, 3000 s.n.m., Departamento de Trancas, Provincia de Tucumán.
Liolaemus camarones: FML 13965. 18 Km. al sur de Camarones, Departamento Florentino Ameghino, Provincia de Chubut. FML 23301-08. Camarones, Departamento Florentino Ameghino, Provincia de Chubut.
Liolaemus canqueli: FML 13966 Al oeste de Meseta El Sombrero, 1.2 Km. al norte de intersección entre Ruta Provincial 53 y Ruta Provincial 90, mojón 1778, Departamento Paso de Indios, Provincia de Chubut. FML 22212: 11 Km. este de Pocitos de Quichaura, Departamento de Languiñeo, Provincia de Chubut.
Liolaemus casamiquelai (Liolaemus sp. 8 en Abdala 2007a): FML 22240 768-74. 9.1 Km. al este de Maquinchao, mojón 330, de Ruta Provincial 23, Departamento 25 de Mayo, Provincia de Río Negro.
Liolaemus fitzingerii: FML 2130. Islote Galfrasconi, frente a Puerto Melo, a 35 Km. al suroeste de Camarones, Departamento de Florentino Ameghino, Provincia de Chubut. FML 02428. Punta Maqueda, 36 Km. al sur de Comodoro Rivadavia, Departamento de Puerto Deseado, Provincia de Santa Cruz. MACN 31288-89. Río Fénix, Lago Buenos Aires, Departamento de Lago Buenos Aires, Provincia de Santa Cruz.
Liolaemus goetschi: FML 21501-511. Laguna Playa - 16,9 km al norte de intersección de Rutas Nacional 22 y provincial 6 - por Ruta Provincial 6 - al norte de General Roca, Departamento de General Roca, Provincia de Río Negro.
Liolaemus morenoi: FML 22206-210 20 Km. al suroeste de Zapala, Ruta Nacional 40, Departamento de Zapala, Provincia de Neuquén. FML 17020-028 2 Km. al norte de Cerrito piñón, Departamento de Collón Curá, Provincia de Neuquén.
Liolaemus telsen: FML 116266-67 80 Km. al oeste de Telsen, Departamento de Telsen, Provincia de Chubut. FML 16935-39 Meseta de Telsen, 80 Km. al oeste de Telsen, Departamento de Telsen, Provincia de Chubut.
Liolaemus xanthoviridis: FML 21259-265: 22 Km. al noroeste de Dos Pozos por Ruta Provincial 75, Provincia de Chubut.
Lista de Acrónimos utilizados en los Apéndices I y II:
FML: Fundación Miguel Lillo, Tucumán, Argentina.
MLP: Museo de La Plata, Buenos Aires, Argentina.
MACN: MuseoArgentino de Ciencias Naturales, Buenos Aires, Argentina.
MCN: Museo de Ciencias Naturales, Provincia de Salta, Argentina.
IBA: Instituto de biología Animal, Mendoza, Argentina.
JV: José Villavicencio.
LITERATURA CITADA
1. Abdala, C.S. 2007a. Phylogeny of the boulengeri group (Iguania: Liolaemidae, Liolaemus) based on morphological and molecular characters. Zootaxa 1538: 1-84. [ Links ]
2. Abdala, C.S. 2007b. Estatus de Liolaemus ditadai, Cei 1983, (Iguania: Liolaemidae) y ampliación de su rango de distribución. Cuadernos de Herpetologia 21: 59-63. [ Links ]
3. Abdala, V.; Abdala C.S. & Tulli, M.J. 2006. Three traditional muscular characters in the phylogeny of Liolaemus (Squamata: Tropiduridae), a reappraisal. Zootaxa 1205: 55-68 [ Links ]
4. Abdala, C.S.; Acosta, J. L.; Acosta, J. C.; Alvarez, B.; Arias, F.; Avila, L.; Blanco, G.; Bonino, M.; Boretto, J. M.; Brancatelli, G.; Breitman, M. F.; Cabrera, M. R.; Cairo, S.; Corbalán, V.; Hernando, A.; Ibargüengoytía , N. R.; Kacoliris, F.; Laspiur, A.; Montero, R.; Morando, M.; Pelegrin, N,; Perez, C. H. F.; Quinteros, S.; Semhan, R.; Tedesco, M. E.; Vega, L.; Zalba, S. M. . 2012. Categorización del estado de conservación de las lagartijas y anfisbenas de la República Argentina. Cuadernos de Herpetología 26: 215-248. [ Links ]
5. Acosta, J.C. & Murúa, A.F. 1997. Geographic Distribution. Liolaemus pseudoanomalus. Herpetological Review 28: 51. [ Links ]
6. Acosta, J.C. & Murúa, F. 1998. Lista preliminar y estado de conservación de los reptiles del Parque Natural Ischigualasto, San Juan - Argentina. Multequina 7:49-59. [ Links ]
7. Aparicio, J. & Ocampo, M. 2010. Liolaemus grupo montanus Etehridge, 1995 (Iguania: Liolaemidae). Cuadernos de Herpetología 24: 133-135. [ Links ]
8. Avila, L.J.; Acosta, J.C. & Murúa, A.F. 1998. Herpetofauna de la provincia de San Juan, Argentina: lista comentada de distribución geográfica. Cuadernos de Herpetologia 12: 11-29. [ Links ]
9. Avila. L.J.; Montero, R.; Morando, M.; Tedesco, M.E.; Acosta, J.C.; Federico, L.; Lobo, F. & Vega, L. 2000. Categorización de las lagartijas y anfisbenas de Argentina. En: Lavilla, E.O.; Richard, E. & Scrocchi, G. (eds.). Categorización de los anfibios y reptiles de la República Argentina. Asociación Herpetológica Argentina, Tucumán. [ Links ]
10. Avila, L.J.; Morando, M. & Belver, L. 2003. Natural history and geographic distribution of the desert lizard Liolaemus pseudoanomalus (Squamata: Iguania: Liolaemidae) from the northern monte region, Argentina. Bulletin of Chicago Herpetology Society 38: 52-53. [ Links ]
11. Avila, L.J.; Morando, M. & Sites Jr., J.W. 2006. Congeneric phylogeography: hypothesizing species limits and evolutionary processes in Patagonian lizard of the Liolaemus boulengeri group (Squamata: Liolaemini). Biological Journal of the Linnean Society 89: 241-275. [ Links ]
12. Avila, L.J.; Morando, M. & Sites Jr., J.W. 2008. New species of the iguanian lizard genus Liolaemus (Squamata, Iguania, Liolaemini) from central Patagonia, Argentina. Journal of Herpetology 42:186-196. [ Links ]
13. Avila, L.J.; Perez, F.C.H.; Morando, M. & Sites Jr., J.W. 2010. A new species of Liolaemus (Reptilia: Squamata) from southwestern Rio Negro province, northern Patagonia, Argentina. Zootaxa 2434: 47-59. [ Links ]
14. Belver, L. & Avila, L.J. 1999. Ecología trofica de Liolaemus pseudoanomalus (Tropiduridae: Liolaeminae). V Congreso Latinoamericano de Hepetología. Montevideo. Publicación Extra 50: 36. [ Links ]
15. Breitman, M.F.; Avila, L.J.; Sites, J.W. & Morando, M. 2011. Lizards from the end of the world: Phylogenetic relationships of the Liolaemus lineomaculatus section (Squamata: Iguania: Liolaemini). Molecular Phylogenetics and Evolution 59: 364-376. [ Links ]
16. Burmeister, H. 1861. Reise durch die La Plata Staaten mit besonderer Rücksicht auf die physische Beschaffenheit und den Culturzustand der Argentinische Republik. Ausgeführt in den Jahren 1857, 1858, 1859 und 1860. Halle: H. W. Schmidt. [ Links ]
17. Cabrera, A.L. & Willink, A. 1980. Biogeografía de América Latina. Segunda edición. Monografía 13, serie biología. Programa Regional de Desarrollo Científico y Tecnológico, Organización de Organización de los Estados Americanos. [ Links ]
18. Cánovas, M.G.; Acosta, J.; Villavicencio, J.C. & Marinero, J.A. 2006. Dimorfismo sexual y morfometría de una población de Liolaemus olongasta (iguania: Liolaeminae) en La Laja, Albardón, San Juan, República Argentina. Cuadernos de Herpetologia 19: 57-61. [ Links ]
19. Cei, J.M. 1971. Further comments on the holotype of Liolaemus ruizleali Donoso Barros and Cei, 1971, from northern Patagonia, Argentina (Reptilia, Iguanidae). Museo Regionale Scienze Naturali 8: 45-52. [ Links ]
20. Cei, J.M. 1974. Two new species of Ctenoblepharis (Reptilia, Iguanidae) from the arid environments of the central Argentina (Mendoza Province). Journal of Herpetology 8: 71-75. [ Links ]
21. Cei, J.M. 1978. Un grupo de reptiles excepcionlmente adaptados a condiciones de vida en ambientes arenosos y eremicos: los iguanidos tropidurinos argentines del grupo "Ortolaemus" (Liolaemus multimaculatus- L. rabinoi). Publicación Ocasional Instituto de Biología Animal Universidad Nacional de Cuyo 7: 1-4. [ Links ]
22. Cei, J.M. 1979. Remarks on the South American lizard Liolaemus anomalus Koslowsky, and the synonomy of Phrynosaura werneri Müller (Reptilia, Lacertilia, Iguanidae). Journal of Herpetology 13: 183-186. [ Links ]
23. Cei, J.M. 1980. Remarks on taxonomic status and specific characters of Liolaemus marmoratus (Burmeister). Herpetologica 14: 192-193. [ Links ]
24. Cei, J.M. 1981. Liolaemus pseudoanomalus a substitute name for Liolaemus marmoratus (Burmeister 1861). Journal of Herpetology 15: 253-254. [ Links ]
25. Cei, J.M. 1983. Una nueva subespecie de Liolaemus anomalus de la región árida halófila de Salinas Grandes (Provincia de Córdoba, Argentina). Deserta (Mendoza) 7: 172-178. [ Links ]
26. Cei, J.M. 1986. Reptiles del centro, centro-oeste y sur de la Argentina. Herpetofauna de las zonas áridas y semiáridas. Museo Regionale di Scienze Naturali, Torino, Monografie 4: 1-527. [ Links ]
27. Cei, J.M. 1993. Reptiles del Noroeste, Nordeste y Este de la Argentina. Herpetofauna de las selvas subtropicales, Puna y Pampas. Museo Regionale di Scienze Naturali Torino Monographie 14: 1-929. [ Links ]
28. Donoso-Barros, R. 1969. Consideraciones nomenclaturales sobre dos lagartijas Argentinas. Boletín Sociedad de Biología de Concepción 41: 93-94. [ Links ]
29. Donoso-Barros, R. 1971. The genera Ctenoblepharis and Phrynosaura. Herpetological Review 3: 85. [ Links ]
30. Duméril, A.M.C. & Bibron, G. 1837. Erpétologie générale ou histoire naturelle complete des reptiles. Ed. Roret, Paris. [ Links ]
31. Espinoza, R.E.; Wiens, J.J. & Tracy, C.R. 2004. Recurrent evolution of herbivory in small, cold-climate lizards: breaking the ecophysiological rules of reptilian herbivory. Proceedings of the National Academy of Sciences, USA 101:16819-16824. [ Links ]
32. Etheridge, R.E. 1993. Lizards of the Liolaemus darwinii complex (Squamata: Iguania: Tropiduridae) in northern Argentina. Bollettino del Museo Regionale di Scienze Naturali, Torino 11: 137-199. [ Links ]
33. Etheridge, R.E. 1995. Redescription of Ctenoblepharys adspersa Tschudi, 1845, and the taxonomy of Liolaeminae (Reptilia: Squamata: Tropiduridae). American Museum Novitates 3142: 1-34. [ Links ]
34. Etheridge, R.E. 2000. A review of the Liolaemus wiegmannii group (Squamata, Iguania, Tropiduridae), and a history of morphological change in the sand-dwelling species. Herpetological Monographs 14: 293-352. [ Links ]
35. Etheridge, R. & Espinoza, R.E. 2000. Taxonomy of the Liolaeminae (Squamata: Iguania: Tropiduridae) and a semi-annotated bibliography. Smithsonian Herpetological Information Service 126: 1-64. [ Links ]
36. Ferraro, D.P. & Williams, J.D. 2006. Material tipo de la colección herpetologica del Museo de La Plata, Buenos Aires, Argentina. Cuadernos de Herpetología 19: 19-36. [ Links ]
37. Frost, D.R. & Etheridge, R.E. 1989. A phylogenetic analysis and taxonomy of iguanian lizards (Reptilia: Squamata). University of Kansas Museum of Natural History Museum, Miscellaneous Publication 81: 1-65. [ Links ]
38. Gallardo, J.M. 1966. Liolaemus lentus nov. sp. (Iguanidae) de La Pampa y algunas observaciones sobre los saurios de dicha provincia Argentina y del oeste de Buenos Aires. Neotrópica (Buenos Aires) 12: 15-29. [ Links ]
39. Girard, C. 1858. Descriptions of some new reptiles collected by the U.S. Exploring Expedition, under the command of Capt. Charles Wilkes, U.S.N., Fourth part. - Including the species of Saurians, exotic to North America. Proceedings of the Academy of Natural Sciences of Philadelphia 9: 195-199. [ Links ]
40. Goloboff, P. 1993. Estimating character weights during tree search. Cladistics 9: 83-91. [ Links ]
41. Goloboff, P.; Farris, J. & Nixon, K. 2003. T.N.T.: Tree Analisis Using New Technology. Disponible en: <www.zmuc.dk/public/phylogeny>. Ultimo acceso: 12 de diciembre de 2012. [ Links ]
42. Goloboff, P.A.; Mattoni, C.I. & Quinteros, A.S. 2006. Continuous characters analyzed as such. Cladistics 22: 589-601. [ Links ]
43. Gravenhorst, J.L.C. 1838. (Sauria-Liolaeminae). Tesis de Magister, Universidad de Chile. [ Links ]
44. Halloy, M. & Castillo, M. 2006. Forelimb wave displays in lizard species of the genus Liolaemus (Iguania: Liolaemidae). Herpetological Natural History 9: 127-133. [ Links ]
45. Koslowsky, J. 1895. Batracios y reptiles de La Rioja y Catamarca (República Argentina) recogidos durante los meses de Febrero a Mayo de 1895 (expedición del Director del Museo). Revista Museo La Plata 6: 357-370. [ Links ]
46. Koslowsky, J. 1896. Sobre algunos reptiles de patagonia y otras regiones argentinas. Revista Museo La Plata 7: 447-457. [ Links ]
47. Lavilla, E.O.; Richard, E. & Scrocchi, G. 2000. Categorización de los anfibios y reptiles de la Republica Argentina. Asociación Herpetológica Argentina, Tucumán. [ Links ]
48. Laurent, R.F. 1982. Description de trois espèces nouvelles du genre Liolaemus (Sauria, Iguanidae). Spixiana 5: 139-147. [ Links ]
49. Laurent, R.F. 1983. Contribución al conocimiento de la estructura taxonómica del género Liolaemus Wiegmann (Iguanidae). Boletín de la Asociación Herpetológica Argentina 1:15-18. [ Links ]
50. Laurent, R.F. 1984. On some iguanid genera related to or previously confused with Liolaemus Wiegmann. Journal of Herpetology 18: 357-373. [ Links ]
51. Laurent, R.F. 1985. Segunda contribución al conocimiento de la estructura taxonómica del género Liolaemus Wiegmann (Iguanidae). Cuadernos de Herpetologia 1: 1-37. [ Links ]
52. Laurent, R.F. 1992. On some overlooked species of the genus Liolaemus Wiegmann (Reptilia Tropiduridae) from Peru. Breviora 494: 1-33. [ Links ]
53. Lesson, R.P. 1830. Observations générales sur les reptiles recueillis dan la voyage de la corvette La Coquille. Pp. 1-66, En: Duperry M. (ed.), Voyage autour du monde exécuté par ordre du Roi sur la corvette de Sa Majesté La Coquille pendant les années 1822, 1823, 1824 et 1825.par M.L.I. Duperrey Zoologie. Tom. II, partie 1, (chapter 9). Arthus Bertrand, Paris. [ Links ]
54. Lobo, F. 2001. A phylogenetic analysis of lizards of the Liolaemus chilensis group (Iguania: Tropiduridae). Journal of Herpetology 11:137-150. [ Links ]
55. Lobo, F. 2005. Las relaciones filogenéticas dentro del grupo chiliensis (Iguania: Liolaemidae: Liolaemus): sumando nuevos caracteres y taxones. Acta Zoológica Lilloana 49: 67-89. [ Links ]
56. Lobo, F. & Abdala, C. 2001. Variación morfológica en el esqueleto de Liolaemus (Iguania: Liolaemidae). Búsqueda y descripción de caracteres. Cuadernos de Herpetologia 15: 119-135. [ Links ]
57. Lobo, F. & Abdala, C. 2002. La información cladística de un set de datos morfológicos en lagartos del género Liolaemus (Iguania: Liolaemidae). Cuadernos de Herpetologia 16: 137-150. [ Links ]
58. Lobo F.; Espinoza, R.E. & Quinteros, S. 2010. A critical review and systematic discussion of recent classification proposals for liolaemid lizards. Zootaxa 2549: 1-30. [ Links ]
59. Marquez, J. 1999. Las áreas protegidas de la provincial de San Juan. Multequina 8: 1-10. [ Links ]
60. Morando, M. 2004. Sistemática y filogenia de grupos de especies de los géneros Phymaturus y Liolaemus (Squamata: Tropiduridae: Liolaemidae). Tesis Doctoral Universidad Nacional de Tucumán. [ Links ]
61. Morando, M.; Avila, L.J. & Sites Jr., J.W. 2003. Sampling strategies for delimiting species: genes, individuals, and populations in the Liolaemus elongatus-kriegi complex (Squamata: Liolaemidae) in Andean-Patagonian South America. Systematic Biology 52: 159-185. [ Links ]
62. Müller, L. 1928. Herpetologische Mitteilungen. Zoologischer Anzeiger 77: 61-84. [ Links ]
63. Núñez, H. & Yánez, J. 1984. Abas y Velosaura nuevos géneros de lagartos Iguanidae y proposiciones sistemáticas respecto de los grupos aliados (Reptilia: Squamata). Boletín del Museo Nacional de Historia Natural Chile 40: 97-105. [ Links ]
64. Ortiz, J.C. 1981. Révision taxonomique et biologie des Liolaemus du groupe nigromaculatus (Squamata, Iguanidae). Thèse de Doctorat d'Étatès Sciences Naturelles, Université Paris VII. [ Links ]
65. Pérez, C.H.F. & Avila, L.J. 2011. First record of Liolaemus lentus Gallardo, 1966 (Squamata, Iguania, Liolaemini) in Río Negro province, Argentina. Herpetology Notes 4: 191-193. [ Links ]
66. Peters, J.A. & Donoso-Barros, R. 1970. Catalogue of the Neotropical Squamata: Part II. Lizards and amphisbaenians. Bulletin of the United States National Museum 297: 1-293. [ Links ]
67. Pincheira-Donoso, D.; Scolaro, J.A. & Sura, P. 2008 A monographic catalogue on the systematics and phylogeny of the South American iguanian lizard family Liolaemidae (Squamata, Iguania). Zootaxa 1800: 1-85. [ Links ]
68. Schulte, J.A.; Macey, J.R.; Espinoza, R.E. & Larson, A. 2000. Phylogenetic relationships in the iguanid lizard genus Liolaemus: multiple origins of viviparous reproduction and evidence for recurring andean vicariance and dispersal. Biological Journal of the Linnean Society 69: 75-102. [ Links ]
69. Statsoft, Inc. 2000. Statistica for Windows. Tulsa OX, Estados Unidos de Norteamérica. [ Links ]
70. Villavicencio, H.J.; Acosta, J.C.; Cánovas, M.G. & Marinero, A. 2002. Patrones de actividad temporal diaria y estacional de Liolaemus pseudoanomalus (Squamata: Tropiduridae) en el centro-oeste de Argentina. Multiquenia 11: 51-60. [ Links ]
71. Villavicencio, H.J.; Acosta, J.C; Cánovas, M.G. & Marinero, A. 2003. Dimorfismo sexual de Liolaemus pseudoanomalus (Squamata: Tropiduridae), en el centro-oeste de Argentina. Revista Española de Herpetologia 17: 87-92. [ Links ]
72. Villavicencio, H.J.; Acosta, J.C.; Cánovas, M.G. & Marinero, A. 2006. Space use in Liolaemus pseudoanomalus (Iguania: Liolaeminae) in central-western Argentina. Bulletin of the Maryland Herpetological Society 42: 1-7. [ Links ]
73. Villavicencio, H.J.; Acosta, J.C.; Marinero, A. & Cánovas, M.G. 2007. Thermal ecology of a population of the lizard, Liolaemus pseudoanomalus in western Argentina. Amphibia- Reptilia 28: 163-165. [ Links ]
74. Wiens, J.J. 1995. Polymorphic characters in phylogenetic systematics. Systematic Biology 44: 482-500. [ Links ]
75. Williams, J.D. & Cei, J.M. 1983. Redescubrimiento de los sintipos de Liolaemus anomalus Koslowsky, 1896: comentarios sobre la distribución de la especie. Boletín de la Asociación Herpetologica Argentina 1: 7-8. [ Links ]
