










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkCuadernos de herpetología
versão On-line ISSN 1852-5768
Cuad. herpetol. vol.29 no.1 San Salvador de Jujuy maio 2015
TRABAJO
Reptiles escamosos (Squamata) del Pleistoceno Medio del Norte de la ciudad de Mar del Plata (provincia de Buenos Aires, Argentina)
Santiago Brizuela1, Marcos M. Cenizo2,3, Daniel A. Tassara4
1 CONICET, Departamento de Biología, Universidad Nacional de Mar del Plata, Funes 3250, B7602AYL. Mar del Plata, Argentina.
2 Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151 (6300), Santa Rosa, La Pampa, Argentina.
3 Área Paleontología, Fundación de Historia Natural "Félix de Azara", Departamento de Ciencias Naturales y Antropología, CEBBAD - Universidad Maimónides, Hidalgo 775 7º piso (1405), Buenos Aires, Argentina.
4 Museo Municipal de Ciencias Naturales "Pachamama", Niza 1065 (7609), Santa Clara del Mar, Buenos Aires, Argentina.
Recibido: 06/06/14
Revisado: 25/08/14
Aceptado: 18/09/14
ABSTRACT
In this study, we describe two fossil remains of squamate reptiles found in Middle Pleistocene outcrops at the northern marine cliffs of the city of Mar del Plata (Buenos Aires province). The specimens were found forming a taphocenosis with remains of other microvertebrates (amphibians, mammals and birds). The reptiles recognized in the association are represented by remains of an undetermined colubrid, and the anguid Ophiodes sp. This latter finding represents the first fossil record for the family Anguidae exhumed in Argentina.
Key words: Fossil; Pleistocene; Ophiodes; `Colubridae´; Argentina.
RESUMEN
En este trabajo se describen restos fósiles de dos reptiles escamosos hallados en afloramientos correspondientes al Pleistoceno Medio de los acantilados marinos ubicados al Norte de la ciudad de Mar del Plata (provincia de Buenos Aires). Los especimenes fueron encontrados conformando una tafocenosis junto a restos de otros microvertebrados (anfibios, mamíferos y aves). Los reptiles reconocidos en la asociación están representados por restos de un colúbrido indeterminado y del ánguido Ophiodes sp. Este último hallazgo constituye el primer registro fósil para la familia Anguidae recuperado en en Argentina.
Palabras clave: Fósil; Pleistoceno; Ophiodes; `Colubridae´; Argentina.
INTRODUCCION
En los últimos años se ha incrementado el registro de ensambles de escamosos fósiles procedentes del Neógeno y Cuaternario de Argentina (Albino, 2005, 2008; Albino y Montalvo, 2006; Albino et al., 2013, Agnolín et al., 2011, Brizuela y Cruz, 2013). Esto ha permitido comenzar a profundizar el conocimiento sobre la evolución y diversidad pasada de este grupo de reptiles. No obstante, el registro argentino de lagartos disponible se encuentra restringido principalmente a las familias Teiidae e Iguanidae (sensu Schulte et al., 2003), en tanto otras familias ampliamente distribuidas en la actualidad muestran hasta el momento un registro nulo (Anguidae, Gymnophthalmidae) o dudoso (Scincidae) (Rodriguez Loredo, 1998; Brizuela y Tassara, 2012).
Recientemente, una rica asociación de microvertebrados ha sido recuperada de los depósitos sedimentarios que constituyen los acantilados ubicados al norte de la ciudad de Mar del Plata. Previamente, entre los taxones exhumados se reconoció preliminarmente la presencia de especímenes fósiles de reptiles escamosos, correspondientes tanto serpientes, como a la de un ánguido (Brizuela y Tassara, 2011, 2012). Este registro de ánguido presenta relevancia ya que constituiría el primero para este particular grupo de lagartos, actualmente representados en América del Sur por sólo dos géneros de diploglossinos (Diploglossinae): Diploglossus (D. fasciatus, D. lessonae, D. monotropis) y Ophiodes (O. intermedius, O. striatus, O. vertebralis, O. yacupoi) (Ávila-Pires, 1995; Savage et al. 2008). En esta contribución presentamos la descripción y asignación taxonómica formal de dichos restos. Estos nuevos registros permiten contar con nuevas evidencias acerca de la composición de la herpetofauna durante el Pleistoceno de la región pampeana.
Contexto del hallazgo
Los materiales aquí descriptos fueron colectados en las exposiciones sedimentarias del frente acantilado ubicado entre las calles "P. Mugaburu" y "A. Storni" (37°57'46.53"S - 57°32'24.33"W) al Norte de la ciudad de Mar del Plata (Partido de General Pueyrredón, provincia de Buenos Aires, Fig. 1A). El área de hallazgo fue referida con anterioridad como sector "Constitución" (Pardiñas et al., 2004; Verzi et al., 2004; Taglioretti et al., 2009).
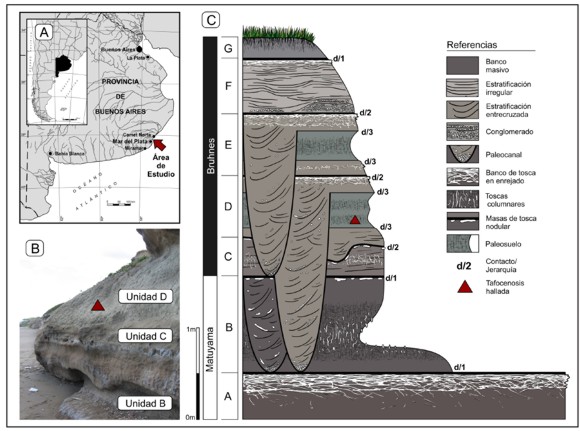
Figura 1. Ubicación geográfica y posición estratigráfica de los ejemplares descriptos. A. Localidad del hallazgo; B. Imagen de la sucesión estratigráfica local. C. Perfil estratigráfico generalizado del área de Constitución (véase texto).
La prospección de la localidad del hallazgo y sectores lindantes permitió el reconocimiento de las entidades estratigráficas seguidamente descriptas (Fig. 1B y C). La unidad de base (A) se encuentra compuesta por un banco tabular marrón rojizo de limos areno-arcillosos masivos, el cual puede presentar concreciones aisladas de carbonatos. Con una potencia de 0,4 a 1,20 m y base no visible, dicha unidad exhibe característicamente una fuerte compactación y un conspicuo banco de tosca en enrejado coronando su techo. Sobre una marcada discordancia erosiva descansa la unidad B correspondiente a limos arcillo-arenosos masivos de hasta dos metros de potencia y coloración marrón rojizo oscura. Esta unidad presenta una superficie cuarteada y en su sector medio desarrolla un notable paleosuelo con nódulos de tosca columnares, donde la geometría tabular del depósito es seccionada por numerosos procesos erosivos. El contacto con la unidad suprayacente (C) es neto, presentando algunas masas nodulares de tosca. La unidad C está representada por un banco de espesor muy variable (entre 0,30 y 1,30 m), compuesto por limos arenosos marrón grisáceos de estructura general masiva, pero que hacia el NE puede incluir bancos pseudoestratificados con tosquillas columnares débiles. Sobre una discordancia erosiva con masas nodulares de tosca se encuentra depositada la unidad D, correspondientes a un conjunto heterogéneo de facies limo areno-arcillosas grisáceo amarillentas y estratificación irregular que desarrollan en su sector medio facies areno-arcillosas verde grisáceas de superficie cuarteada y estructura en media caña correspondiente a dos paleosuelos superpuestos. Hacia el tope este depósito exhibe profundos canales seccionando las unidades infrayacentes hasta el contacto con la unidad A, su techo es coronado por un banco de tosca en enrejado y/o masiva. Una discordancia erosiva la separa de la unidad E cuyas características repiten aquellas observadas en la unidad precedente. Sobre una discordancia erosiva descansa la unidad F compuesta por arenas limosas gris amarillentas con marcada estratificación irregular y bancos conglomerádicos de tosquilla en su base. La sucesión estratigráfica culmina con suelos actuales vegetados incluidos en la unidad G. Específicamente, en el área del hallazgo se encuentran representadas la sucesión de unidades B-G, solo en sectores de playa lindantes al norte puede reconocerse la unidad A.
Ameghino (1908) fue el primero en reconocer una marcada discordancia erosiva regional ubicada a unos 7 m del nivel de base, la cual permitiría segregar localmente sus pisos Ensenadense y Bonaerense. Las acumulaciones modernas de arenas de playa han producido la obliteración de los niveles basales expuestos a principios del siglo XX, por lo que es posible que la discordancia señalada por Ameghino se encuentre hoy representada en la base de los acantilados y corresponda a los contactos entre las unidades B-C aquí descriptas. Posteriormente, Kraglievich (1952, 1953, 1959) identificó en el área un conjunto de unidades originalmente descriptas para el tramo de acantilados expuestos entre Mar del Plata y Miramar. El reconocimiento de estas entidades es una tarea difícil, sin embargo, las descripciones brindadas por dicho autor se corresponden con las aquí definidas, reconociéndose en el área de estudio la posible presencia de sus "Formaciones" Vorohué (unidad A), San Andrés (unidad B), Miramar (unidades C-D?) y Arroyo Seco (unidad E-F?). Los estudios paleomagnéticos disponibles para la región indican que el límite entre los crones Brunhes-Matuyama se encontraría posiblemente ubicado entre las unidades B y C (Unidad 4 e inferiores de Bidegain et al., 2005; Fig 1. C), por lo que la edad de la unidad C y suprayacentes sería menor a 780 ka AP. En tanto, Verzi et al. (2004; véase también Taglioretti et al., 2009) informaron el hallazgo en niveles medios de Constitución de restos asignados a Ctenomys kraglievichi, fósil guía de la biozona homónima propuesta como base del Piso-Edad Bonaerense y correspondiente al Pleistoceno Medio (circa 400 ka; Verzi et al., 2004, Cione y Tonni, 2005). Las características y posición estratigráfica de la unidad portadora de C. kraglievichi en la localidad de Constitución descriptas por estos autores son congruentes con las unidades regionales U3 y U2 descriptas por Bidegain et al. (2005, véase Verzi et al., 2004; Fig. 5) y unidades D y E de nuestro perfil.
El material de reptiles escamosos aquí descripto formaba parte de una tafocenosis de microvertebrados integrada por anfibios (Anura), roedores (Cricetidae), junto a aves acuáticas (Podicipedidae, Charadriidae, Anatidae) y terrestres (Strigidae, Tinamidae, Passeriformes) actualmente bajo estudio. La asociación se encontraba diseminada en la base del paleosuelo inferior desarrollado sobre la unidad D, presentando una dispersión horizontal aproximada de un metro. De acuerdo con sus atributos tafonómicos, composición taxonómica y contexto sedimentario, el conjunto faunístico ha sido interpretado preliminarmente como correspondiente a egagrópilas o restos de presas de aves rapaces. Este tipo de agregados óseos ya han sido previamente reportados para el área y parecen constituir un modo tafonómico frecuente en las sucesiones sedimentarias locales (Vucetich et al., 1997; Tonni et al., 1998, Verzi et al., 2002; Pardiñas, 2004; Pardiñas et al., 2004; Cenizo y de los Reyes, 2008; Taglioretti et al., 2009). Depósitos similares al nivel portador de los ejemplares aquí descriptos, han sido interpretados como cuerpos de agua pedogenizados al disminuir las condiciones de humedad ambiental (Osterrieth y Martínez, 1990).
MATERIALES AND METODOS
El material estudiado se encuentra depositado, con el resto de los fósiles del ensamble herpetológico (Brizuela y Tassara, 2011), en la colección de la Sección Paleontología del Museo Municipal de Ciencias Naturales "Lorenzo Scaglia" (MMP), bajo el número MMP 5194. Las vértebras fósiles han sido diferenciadas numéricamente como MMP 5194-1 y MMP 5194-2 de los restantes elementos fósiles.
Para las comparaciones osteológicas se utilizaron ejemplares actuales depositados en las colecciones de la Universidad Nacional de Mar del Plata (UNMdP, Colección Herpetológica; UNMdP-O, Colección Herpetológica, sección Osteología) y de la Universidad Nacional Nordeste (UNNE) (Apéndice). Las distintas vertebras se fotografiaron con el microscopio electrónico de barrido del Laboratorio de Microscopia de la Universidad Nacional de Mar del Plata. La terminología y medidas empleadas en las comparaciones corresponden principalmente a Hoffstetter y Gasc (1969) y Albino (2011).
La sistemática de Serpentes y por lo tanto la de Colubroidea se encuentra actualmente en un estado de fluctuación, con discrepancias entre distintas topologías moleculares y morfológicas (Lawson et al., 2005; Zaher et al., 2009; Pyron et al., 2011). En este trabajo, por su enfoque morfológico, se opta por seguir a Zaher (1999). Así, entre los colubroideos, solo Viperidae, Elapidae y Atractaspididae presentan sinapomorfías no ambiguas, los restantes colubroideos se agrupan en `Colubridae´.
Paleontología sistemática
Squamata Oppel, 1811
Scleroglossa Estes et al., 1988
Autarchoglossa Wagler, 1830
Anguimorpha Fürbringer, 1900
Anguidae Gray, 1825
Diploglossinae Bocourt, 1873
Ophiodes Wagler, 1828
Ophiodes sp.
(Fig. 2)
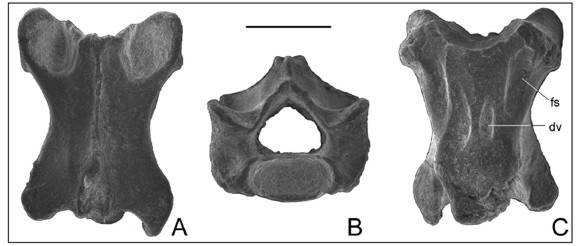
Figura 2. Vértebra presacra (MMP-5194-1) asignada a Ophiodes sp. procedente del Pleistoceno Medio de la costa atlántica de la provincia de Buenos Aires (Argentina). A, vista dorsal; B, vista anterior C, vista ventral. Abreviaturas: fs, fosa subcentral; dv, depresión ventral. Escala 1 mm.
Material referido. MMP 5194-1, vértebra presacra posterior.
Procedencia geográfica estratigráfica. Sitio "Constitución", Mar del Plata, provincia de Buenos Aires. Unidad D (Fig. 1C), Piso-Edad Bonaerense, Biozona de C. kraglievichi, Pleistoceno Medio.
Descripción. Vértebra procélica pequeña (longitud del centro vertebral = 2,08 mm), grácil y relativamente alargada con una leve constricción entre las pre y postzigapófisis (Fig. 2A). La espina neural se extiende a lo largo de todo el arco neural, en vista anterior es delgada y baja, mientras que posteriormente se eleva y se engrosa considerablemente a nivel de las postzigapófisis. La espina neural se encuentra fragmentada distalmente, sin embargo, es evidente que se extendía posteriormente más allá del límite posterior del arco neural. El límite anteromedial del arco neural se prolonga hasta la mitad de la longitud anterior de las prezigapófisis. La parte anterior del arco neural es estrecha e inclinada a ambos lados de la espina neural, careciendo de carillas de articulación zigosfénicas. En vista anterior (Fig. 2B), el arco neural es amplio, más alto que el cótilo y de contorno triangular. Las prezigapófisis se originan a media altura del arco neural, encontrándose inclinadas 45º (aprox.) con respecto al plano horizontal y orientadas anterolateralmente en vista dorsal. Sus carillas de articulación son ligeramente ovales. En vista dorsal, el contorno de la postzigapófisis derecha se extiende posteriormente, acompañando la carilla de articulación de la postzigapófisis, la cual es oval, anteroposteriormente orientada e inclinada con respecto al plano horizontal. En vista lateral, las zigapófisis están unidas por una conspicua cresta interzigapofisial (= margo lateralis), la cual muestra ventralmente una menos marcada cresta subcentral (= margo ventralis). Entre ambas crestas se observa una leve depresión, anteriormente a la cual se encuentran los procesos transversos. Estos procesos están poco desarrollados y presentan sinapófisis ovales verticalmente dispuestas. En la superficie anterior del proceso transverso izquierdo se observa un pequeño foramen. El centro vertebral se encuentra dorsoventralmente comprimido. Tanto el cótilo y cóndilo son ovales y se encuentran verticalmente orientados. El cótilo es profundo, mientras que el cóndilo no se encuentra diferenciado del centro vertebral (i.e. carece de constricción precondilar). El centro vertebral a simple vista es plano (Fig. 2B), aunque con mayor detalle se diferencian dos fosas subcentrales en la parte anterior, lateralmente a la cresta sagital que presenta una ligera constricción entre el cótilo y el cóndilo. A la altura de dicha constricción, la cresta sagital presenta un par de profundas depresiones ventrales (Fig. 2C).
Comentarios. Blain et al. (2010) sostienen que, entre los reptiles escamosos, vértebras con el centro vertebral plano pueden reconocerse en Amphisbaenia, Anguidae, Varanidae y Helodermatidae. La ausencia de una marcada constricción precondilar permite excluir a MMP 5194-1 de los varánidos en los cuales esta constricción está presente (Hoffstetter y Gasc, 1969). El material descripto difiere de los Helodermatidae, que pueden presentar una sutil constricción precondilar, ya que la espina neural de los helodermátidos es delgada, alta y posterodorsalmente prolongada en un ángulo agudo (Blain et al., 2010), distinto a lo observado en el fósil. MMP 5194-1 difiere de Amphisbaenia tanto por la ausencia de procesos prezigapofisarios como de una proyección anteromedial del arco neural (Hoffstetter y Gasc, 1969). Adicionalmente, el material fósil exhibe un desarrollo considerable de la espina neural, mientras que esta condición no está presente en los anfisbénidos (ver comentario al respecto en Augé, 2012).
Entre los ánguidos, los Anguinae presentan un centro vertebral plano de crestas subcentrales paralelas (Anguis) o algo divergentes anteriormente (Ophisaurus) y ligeramente cóncavas (Meszoely, 1970; Rage y Augé, 2010). Aunque generalmente mayores, las vértebras de los Glyptosaurinae son muy similares a las de los Anguinae, sin embargo a diferencia de estos últimos no presentan un centro vertebral marcadamente plano (Bolet y Evans, 2013). Una situación similar ocurre en los restantes Anguidae (i.e., Diploglossinae y Gerrhonotinae) en donde el centro vertebral es levemente convexo (Augé, 1992). A diferencia de los Gerrhonotinae (Gerrhonotus), los Diploglossinae (Diploglossus y Ophiodes) presentan fosas subcentrales (Meszoely, 1970: Fig. 5; este trabajo). Dicho carácter se observa también en MMP 5194-1 lo que permite diferenciarlo de los Gerrhonotinae y asignarlo a Diploglossinae. Más aún, la morfología general del material procedente del Pleistoceno de Mar del Plata resulta indiferenciable de los elementos homólogos observados en los representantes actuales del género Ophiodes (único Anguidae con distribución en Argentina, Abdala et al., 2012).
En particular, cuando analizamos la variación en la morfología ventral del centro vertebral a lo largo de la columna vertebral en Ophiodes observamos sutiles diferencias en las vértebras presacras anteriores de las dos especies actuales evaluadas (O. vertebralis y O. intermedius). Mientras que en O. vertebralis (UNMdP 162; UNMdP-O 66) la cresta sagital es plana, sin depresiones ventrales, en O. intermedius (UNNE 4842, 4848) la cresta sagital presenta una ligera, pero nítida, concavidad a lo largo de la línea media del centro vertebral (Fig. 3A y C). En ambos casos se observan forámenes subcentrales a ambos lado de la cresta sagital. Las vértebras presacras posteriores presentan mayor variabilidad morfológica de la cara ventral del centro vertebral. En esta zona se observa que la cresta sagital se hace ligeramente cóncava y puede o no mostrar las depresiones ventrales (Fig. 3B y D). No se observó un patrón distintivo en ninguna de las especies en cuanto a las vértebras presacras posteriores.

Figura 3. Vértebras de ejemplares actuales del género Ophiodes. A, vértebra presacra anterior de O. vertebralis (UNMdP-O 66); B, vértebra presacra posterior de O. vertebralis (UNMdP 162); C, vértebra presacra anterior de O. intermedius (UNNE 4848); D, vértebra presacra posterior de O. vertebralis (UNNE 4848). Escala 1 mm.
Por lo antes expuesto, MMP 5194-1 es considerada una vértebra presacra posterior de Ophiodes, ya que cuenta con depresiones ventrales en la cresta sagital. Por el momento sólo se ha apreciado un patrón en la variación en las vértebras presacras anteriores entre las dos especies actuales disponibles. Dado que la vértebra fósil es una dorsal posterior no es posible una asignación a nivel específico.
Serpentes Linnaeus, 1758
Alethinophidia Nopcsa, 1923
Caenophidia Hoffstetter, 1939
Colubroidea Oppel, 1811
`Colubridae´ Oppel, 1811
genero y especie indet.
(Fig. 4)

Figura 4. Vertebras precloacales de distintos colúbridos. A-E, vértebra precloacal (MMP-5194-2) asignada a `Colubridae´ indet. procedente del Pleistoceno Medio de la costa atlántica de la provincia de Buenos Aires (Argentina) en vistas anterior (A), lateral (B), posterior (C), dorsal (D) y ventral (E); F, vértebra precloacal de Philodryas patagoniensis (UNMdP-O 56) en vista frontal; G,vértebra precloacal de Paraphimophis rustica (UNMdP-O 65) en vista lateral; H, vértebra precloacal de Erythrolamprus poecilogyrus (UNMdP-O 62) en vista dorsal. Escala 1 mm.
Material referido. MMP 5194-2, vértebra precloacal.
Procedencia geográfica estratigráfica. Sitio "Constitución", Mar del Plata, provincia de Buenos Aires. Unidad D (Fig. 1C), Piso-Edad Bonaerense, Biozona de C. kraglievichi, Pleistoceno Medio.
Medidas (en mm). Ana, ancho mínimo del arco neural = 1,5; anap, ancho del arco neural inmediatamente posterior a las prezigapófisis = 1,99; cl, longitud del centro vertebral desde el borde ventral del cótilo hasta el extremo más distal del cóndilo =1,76; cona, ancho máximo del cóndilo = 0,88; conh, altura máxima del cóndilo = 0,75; cta, ancho máximo del cótilo = 0,92; cth, altura máxima del cótilo = 0,75; enh, altura de la espina neural en vista anterior = 0,43; h, altura de la vértebra desde la base del cóndilo hasta el extremo dorsal de la espina neural = 2,14; po-po, distancia entre los extremos de las postzigapófisis = 2,74; pr-pr, distancia entre los extremos de las prezigapófisis = 2,65; pr-po, distancia interzigapofisaria, entre el extremo más anterior de la prezigapófisis y el más posterior de la postzigapófisis del mismo lado = 2,18; zga, ancho máximo del zigósfeno = 1,12; zgh, altura del zigósfeno en su parte media = 0,13.
Descripción. Vértebra delicada, pequeña (cl = 1,76 mm) completa a excepción del extremo distal anterior de la espina neural y extremos distales del proceso trasverso y prezigapófisis izquierda. El arco neural es largo y deprimido; dorsalmente, el contorno de las postzigapófisis delimita una marcada muesca. La espina neural es baja, extendiéndose caudalmente por detrás del zigósfeno hasta una corta distancia más allá del contorno de las postzigapófisis. La cresta interzigapofisial (= margo lateralis) es marcada, presentando contornos cóncavos en vista dorsal (i.e. la vértebra se encuentra restringida en su parte media). En vista ventral, el centro vertebral es alargado, con un contorno ligeramente triangular por la divergencia anterior de las levemente desarrolladas crestas subcentrales (= margo ventralia). La superficie ventral se encuentra surcada por una carena hemal plana de contorno espatulado (sensu Auffenberg, 1963). Posterior a la carena hemal, el cóndilo se observa diferenciado por una marcada constricción precondilar. Se observan dos pequeños forámenes subcentrales. En vista lateral, la sinapófisis se diferencia en una diapófisis convexa dorsoposterior a la parapófisis ligeramente cóncava. Ventralmente a la sinapófisis se proyectan cortos procesos parapófisiales, que sobrepasan ligeramente el borde ventral del cótilo. Posterior a la diapófisis, sobre la cara lateral del arco neural se observa el foramen lateral. En vista anterior se observa un amplio canal neural de contorno subcuadrangular, ligeramente trilobulado, y de dimensiones similares al cótilo. El techo anterior del canal neural corresponde al zigósfeno. Este último es delgado, levemente convexo en vista anterior y sutilmente sinusoidal o crenado (sensu Auffenberg, 1963) en vista dorsal. Asimismo, el zigósfeno posee dos largas (en sentido anteroposterior) carillas de articulación levemente inclinadas lateralmente. Estas se diferencian claramente de las carillas de articulación de las prezigapófisis. En vista posterior se observa sobre la postzigapófisis izquierda un foramen parazigantral. Las prezigapófisis de contorno oval, se encuentran anterolateralmente orientadas y poco inclinadas con respecto a la horizontal. En posición ventral con respecto a estas articulaciones se encuentran los procesos prezigapofisarios. El derecho se orienta lateralmente, es poco desarrollado y de contorno obtuso (sensu Auffenberg, 1963). El cótilo es de contorno tenuemente oval y se encuentra orientado ligeramente anteroventralmente. A ambos lados del cótilo y próximo a su borde, se verifica la presencia de forámenes paracotilares. El cóndilo es de contorno circular en vista posterior y se encuentra inclinado en sentido anterior.
Comentarios. La presencia de procesos prezigapofisiales y articulación zigósfeno/zigantro son caracteres que indican que se trata de una vértebra referible a una serpiente (Hoffstetter y Gasc, 1969). Por otra parte, la siguiente combinación de características permiten asignar el material MMP 5194-2 a Colubroidea (Albino y Montalvo, 2006; Hsiou y Albino, 2010): 1, constitución delicada; 2, zigósfeno delgado en vista frontal; 3, espina neural delgada; 4, diapófisis y parapófisis diferenciadas; 5, forámenes paracotilares y procesos paracotilares desarrollados. Entre los colubroideos, las hipapófisis están bien desarrolladas en las vértebras precloacales de Elapidae, Viperidae, y algunos `Colubridae´ (`Homalopsinae´, Natricinae y ciertos Colubrinae) (Hoffstetter y Gasc, 1969), permitiendo excluir al fósil MMP 5194-2 de estas formas. Así, esta vértebra puede ser asignada a los `Colubridae´. El escaso conocimiento disponible acerca de la variabilidad morfológica en las vértebras de las formas actuales de colúbridos, la similitud en las formas presentes en la provincia de Buenos Aires (Albino y Quintana, 1992; Scanferla, 2006), junto a la ausencia de claras sinapomorfías vertebrales que permitan definir categorías taxonómicas inferiores a la familiar (e.g. Lee y Scanlon, 2002) impiden una asignación sistemática más precisa. No obstante, se incluyen aquí algunas apreciaciones con respecto a los taxones previamente reconocidos en el registro fósil de la región pampeana (i.e., Boiruna, Clelia, Philodryas y Xenodon) (Albino y Quintana 1992; Albino, 1999; Scanferla, 2006; Scanferla et al., 2005, 2009). Las vértebras de Philodryas (P. patagoniensis y P. baroni) son más anchas y deprimidas (Albino, 2000) de lo que se observa en el material estudiado, presentando además procesos prezigapofisarios más largos (Fig. 4F). Por otra parte, mientras las espinas neurales de Boiruna y Paraphimophis son altas y cortas (Albino, 1999; Scanferla et al., 2005, 2009) (Fig. 4G), las exhibidas por Xenodon y Erythrolamprus son bajas y largas (Albino y Quintana, 1992; Scanferla, et al., 2005) recordando la condición observada en MMP 5194-2. Sin embargo, Xenodon (i.e., X. dorbignyi) difiere del material de Mar del Plata en la presencia de procesos prezigapofisarios más desarrollados y robustos, de contorno cuadrangular en vista dorsal (Scanferla et al., 2005: Fig. 3D). En tanto que Erythrolamprus (i.e., E. poecilogyrus) presenta el contorno anterior del zigósfeno marcadamente sinusoidal, procesos prezigapofisiales más agudos y, en vista dorsal, la superficie de articulación de las postzigapófisis con características distintas a las observadas en MMP 5194-2 (Fig. 4H). Estas diferencias podrían atribuirse a problemas de preservación del fósil, pero la confirmación de esta hipótesis requiere el hallazgo de material adicional.
DISCUSION Y CONCLUSIONES
De acuerdo con Conrad (2008) y Conrad y Norell (2008) la familia Anguidae incluye un clado extinto (Glyptosaurinae) y tres clados con representantes actuales (Anguinae, Diploglossinae y Gerrhonotinae). Los análisis filogenéticos efectuados por estos autores indican un origen para el grupo previo al Cretácico Tardío, momento en el cual se registra Odaxosaurus piger (Glyptosaurinae) en América del Norte, encontrándose ya diferenciados en esta edad los Diploglossinae de los restantes ánguidos. La temprana diferenciación del grupo contrasta con el extremadamente escaso registro fósil que estos lagartos muestran en América del Sur. Hasta el momento, el mismo se encuentra restringido a la vertebra aquí descripta (MMP 5194-1) y un dentario referido a Diploglossus cf. D. fasciatus procedente del Pleistoceno Tardío-Holoceno del sudeste de Brasil (Camolez y Zaher, 2010). Ambos registros se hallan incluidos dentro de los límites de la distribución geográfica actual registrada para los respectivos géneros (Ophiodes y Diploglossus) (Cei, 1993; Peters y Donoso-Barrios, 1970) por lo que se infiere que dicha distribución estaría presente desde al menos el Pleistoceno Medio. Estes (1983) considera incierto el momento en el cual los diploglossinos arribaron a América del Sur; no obstante, Strahm y Schawart (1977) proponen la existencia al menos desde el Mioceno, de un antecesor común para los dos género actuales presentes en América del Sur.
Los restos más antiguos de colúbridos hallados en la provincia de Buenos Aires se encuentran representados por formas indeterminadas procedentes de unidades pliocénicas de la costa atlántica (Albino y Quintana, 1992). Los géneros actuales de colúbridos Boiruna y Paraphimophis tienen sus primeros registros a partir del Ensenadense (Pleistoceno Temprano-Medio) (Paraphimophis dado como Clelia en Scanferla, 2006; Scanferla et al., 2009); en tanto, la presencia de Xenodon ha sido informada para unidades correspondientes al Bonaerense (Pleistoceno Medio-Tardío) (Scanferla et al., 2005). Por último, ejemplares asignados a las especies Paraphimophis rustica y Philodryas patagoniensis han sido identificados en el sitio arqueológico Cueva Tixi (Pleistoceno Tardío-Holoceno) (Albino, 1999). En este contexto y dada la imposibilidad de una asignación genérica, MMP 5194-2 sólo amplía la distribución de los colúbridos durante el Pleistoceno Medio. Con excepción de Boiruna, los restantes taxones mencionados presentan áreas de distribución que en la actualidad incluyen la provincia de Buenos Aires (Giraudo et al., 2012).
Los escamosos descriptos en este estudio fueron hallados conformando una tafocenosis junto a distintos restos de anfibios entre los cuales se pueden reconocer vértebras y tibiofíbulas de afinidades inciertas, y un fragmento craneal robusto cuya ornamentación indica una vinculación con Ceratophryidae (Brizuela y Tassara, 2011). Por su parte Ophiodes es un lagarto mesófilos, y frecuentemente se lo encuentra asociado con cuerpos de agua (Cei, 1993). La presencia en la asociación exhumada de fósiles referidos a taxones con hábitos acuáticos y/o anfibios, es congruente con las características sedimentarias señaladas para la unidad portadora (ver arriba unidad D), permitiendo inferir la existencia local de cuerpos de agua relativamente estables durante el Pleistoceno Medio.
AGRADECIMIENTOS
Queremos agradecer a A. Dondas y F. Scaglia del Museo Municipal de Ciencias Naturales "Lorenzo Scaglia". A J. F. Vilá y M. Oppedisano del Laboratorio de Microscopía Universidad Nacional de Mar del Plata por sus respectivas colaboraciones. L. Vega (Universidad Nacional de Mar del Plata) y S. Palomas (Universidad Nacional del Nordeste) por facilitar el material de referencia a su cuidado. A dos revisores anónimos y al editor asociado M. Cabrera por sus valiosos comentarios y sugerencias sobre el manuscrito. SB agradece particularmente a A.M. Albino (Universidad Nacional de Mar del Plata).
Bothrops alternatus UNMdP-O; 78Paraphimophis rustica UNMdP-O 65; Lygophis anomalus UNMdP-O 61; Erythrolamprus poecilogyrus UNMdP-O 62, 86; Ophiodes intermedius UNNE 4842, 4848; Ophiodes vertebralis: UNMdP 162; UNMdP-O 66; Philodryas baroni UNMdP-O 38; Philodryas patagoniensis UNMdP-O 55, 56, 64; Xenodon dorbignyi UNMdP-O 59, 60, 63, 79.
LITERATURA CITADA
1. Abdala, C.S.; Acosta, J.L.; Acosta, J.C.; Álvarez, B.B.; Arias, F.; Avila, L.J.; Blanco, G.M.; Bonino, M.; Boretto, J.M.; Brancatlli, G.; Breitman, M.F.; Cabrera, M.R.; Cairo, S.; Corbalán, V.; Hernando, A.; Ibargüengoytía, N.R.; Kacoliris, F.P.; Laspiur, A.; Montero, R.; Morando, M.; Pelegrin, N.; Fluvio Pérez, C.H.; Quinteros, A.S.; Semhan, R.V.; Tedesco, M.E.; Vega, L.E. & Zabala, S.M. 2012. Categorización del estado de conservación de las lagartijas y anfisbenas de la República Argentina. Cuadernos de Herpetología 26: 215-247. [ Links ]
2. Agnolín, F.L. & Jofré, G. 2011. Nuevos registros de Squamata (Reptilia) para el Pleistoceno superior del Norte de la provincia de Buenos Aires, Argentina. Papéis Avulsos de Zoologia, Museu de Zoologia da Universidade de São Paulo 51: 49-58. [ Links ]Albino, A.M. 1999. Serpientes del sitio arqueológico Cueva Tixi (Pleistoceno tardío-Holoceno), provincia de Buenos Aires, Argentina. Ameghiniana 36: 269-273.
3. Albino, A.M. 2005. A late Quaternary lizard assemblage from the southern Pampean Region of Argentina. Journal of Vertebrate Paleontology 25: 185-191.
4. Albino, A.M. 2008. Lagartos iguanios del Colhuehuapense (Mioceno temprano) de Gaiman (provincia del Chubut, Argentina). Ameghiniana 45: 775-782.
5. Albino, A. M. 2011. Morfología vertebral de Boa constrictor (Serpentes: Boidae) y la validez del género Mioceno Pseudoepicrates Auffenberg, 1923. Ameghiniana 48: 53-62.
6. Albino, A.M. & Quintana, C.A. 1992. Los Colubridae (Reptilia: Serpentes) del Chapadmalalense y Uquiense (Plioceno tardío - Pleistoceno temprano?) de la provincia de Buenos Aires. Ameghiniana 29: 125-133.
7. Albino, A.M. & Montalvo, C.I. 2006. Snakes from the Cerro Azul Formation (Upper Miocene), central Argentina, with a review of fossil viperids from South America. Journal of Vertebrate Paleontology 26: 581-587.
8. Albino, A.M.; Montalvo, C.I. & Brizuela, S. 2013. New records of squamates from the Upper Miocene of South America. Journal of Herpetology 47: 590-598.
9. Ameghino, F. 1908. Las formaciones sedimentarias de la región litoral de Mar del Plata Chapadmalán. Anales del Museo Nacional de Buenos Aires 10: 343-428.
10. Auffenberg, W. 1963. The fossil snakes of Florida. Tulane Studies in Zoology 10: 131-216.
11. Augé, M.L. 1992. Une espéce nouvelle ďOphisaurus (Lacertilia, Anguidae) de ľOligocène des phosphorites du Quercy. révision de la sous-famille des Anguinae. Paläontologische Zeitschrift 66: 159-175.
12. Augé, M.L. 2012. Amphisbaenians from the European Eocene: a biogeographical review. Palaeobiodiversity and Palaeoenvironments 92: 425-443.
13. Ávila-Pires, T.C.S. 1995. Lizards of Brazilian Amazonia. Zoologische Verhandelingen 299: 1-706
14. Bidegain, J.C.; Osterrieth, M.L.; Van Velzen, A.J. & Rico, Y. 2005. Geología y registros magnéticos entre arroyo La Tapera y Santa Clara del Mar, Mar del Plata. Revista de la Asociación Geológica Argentina 60: 599-604.
15. Blain, H.-A.; Canudo, J.I.; Cuenca-Bescós, G. & López-Martínez, N. 2010. Amphibians and squamate reptiles from the latest Maastrichtian (Upper Cretaceous) of Blasi 2 (Huesca, Spain). Cretaceous Research 31: 433-446.
16. Bolet, A. & Evans, S. E. 2013. Lizards and amphisbaenians (Reptilia, Squamata) from the late Eocene of Sossís (Catalonia, Spain). Palaeontologia Electronica 16: 23.
17. Brizuela, S. & Tassara, D. 2011. Ensamble herpetológico del Pleistoceno Medio-Superior de Mar del Plata (Argentina). VI Reunión de Biólogos En Red (Mar del Plata). Resúmenes: 20 [ISSN 1853-9998].
18. Brizuela, S. & Tassara, D. 2012. Primer registro fósil de un Anguidae en Argentina (Pleistoceno medio-superior). XIII Congreso Argentino de Herpetología, Libro de Resúmenes: 49.
19. Brizuela, S. & Cruz, L.E. 2013. Escamosos (Squamata) de la Formación Brochero, Plioceno, de la provincia de Córdoba (Argentina). Ameghiniana 50: 9-10.
20. Camolez, T. & Zaher, H. 2010. Levantamento, identificação e descrição da fauna de Squamata do Quaternário brasileiro (Lepidosauria). Arquivos de Zoologia, Museu de Zoologia da Universidade de Sao Paulo 41: 1-96.
21. Cei, J.M. 1993. Reptiles del noroeste, nordeste y este de la Argentina. Museo Regionale di Scienze Naturale, Torino, Monografie 14: 1-949.
22. Cenizo, M.M. & de los Reyes, L.M. 2008. Primeros registros de Tyto alba (Scopoli, 1769) (Strigiformes, Aves) en el Pleistoceno Medio-Tardío de la provincia de Buenos Aires (Argentina) y sus implicancias tafonómicas. Revista del Museo Argentino de Ciencias Naturales 10: 199-209.
23. Cione, A.L. & Tonni, E.P. 2005. Bioestratigrafía basada en mamíferos del Cenozoico superior de la provincia de Buenos Aires, Argentina. En: de Barrio, R.E., Etcheverry, R.O., Caballé, M.F., y Llambías, E. (eds.), Relatorio del XVI Congreso Geológico Argentino 11: 183-200.
24. Conrad, J.L. 2008. Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bulletin American Museum of Natural History 310: 1-182.
25. Conrad, J.L. & Norell, M.A. 2008. The braincases of two glyptosaurines (Anguidae, Squamata) and anguid phylogeny. American Museum Novitates 2613: 1-24.
26. Estes, R. 1983. Handbunch der Palaoherpetologíe. Gustav Fischer Verlag, New York.
27. Giraudo, A.R.; Arzamendia, V.; Bellini, G.P.; Bessa, C.A.; Calamante, C.; Cardozo, G.; Chiaraviglio, M.; Costanzo, M.B.; Etchepare, E.G.; Di Cola, V.; Di Pietro, D.O.; Kretzschmar, S.; Palomas, S.; Nenda, S.J.; Rivera, P.C.; Rodríguez, M.E.; Scrocchi, G.J. & Williams, J.D. 2012. Categorización del estado de conservación de las Serpientes de la República Argentina. Cuadernos de Herpetología 26: 303-326.
28. Hoffstetter, R. & Gasc, J P. 1969. Vertebrae and ribs of modern reptiles: 201-310. En: Gans, C.; Bellairs, A. & Parsons, T.S. (eds.), Biology of the Reptilia, Vol. 1. Academic Press, London and New York.
29. Hsiou, A. & Albino, A. M. 2010. New snake remains from the Miocene of northern South America. Herpetological Journal 20: 249-259.
30. Kraglievich, J.L. 1952. El perfil geológico de Chapadmalal y Miramar, provincia de Buenos Aires. Revista del Museo de Mar del Plata 1: 8-37.
31. Kraglievich, J.L. 1953. La llanura bonaerense a través de un perfil geológico. Revista Mundo Atómico 9: 88-99.
32. Kraglievich, J.L. 1959. Nota acerca de la geología costera en la desembocadura del arroyo Malacara (prov. de Buenos Aires). Revista del Museo Argentina de Ciencias Naturales 1: 3-9.
33. Lawson, R., Slowinski, J.B., Crother, B.I. & Burbrink, F.T. 2005. Phylogeny of the Colubroidea (Serpentes): New evidence from mitochondrial and nuclear genes. Molecular Phylogenetics and Evolution 37: 581-601.
34. Lee, M.S.Y. & Scanlon, J.D. 2002. Snake phylogeny based on osteology, soft anatomy and ecology. Biological Reviews 77: 333-401.
35. Meszoely, C.A.M. 1970. North America fossil anguid lizards. Bulletin of The Museum of Comparative Zoology 139: 87-149.
36.Osterrietch, M.L. & Martínez,G.A. 1990. Paleosuelos en secuencias loessicas de la vertiente nororiental de las sierras septentrionales (Buenos Aires, Argentina): 131-137. En: M. Zárate (ed.), Características, Cronología y Significado Paleoclimático del Loess, Simposio Internacional sobre Loess, Mar del Plata.
37. Pardiñas, U.F.J. 2004. Reodores sigmodontinos (Mammalia: Rodentia: Cricetidae) y otros micromamíferos como indicadores de ambientes hacia el Ensenadense cuspidal en el sudeste de la provincia de Buenos Aires (Argentina). Ameghiniana 41: 437-450.
38. Pardiñas, U.F.J.; Cione, A.L.; San Cristóbal, J.; Verzi, D. & Tonni, E.P. 2004. A new Last Interglacial continental vertebrate assemblage in central-eastern Argentina. Current Research in the Pleistocene 21: 111-112.
39. Peters, J.A. & Donoso-Barros, R. 1970. Catalogue of the Neotropical Squamata. Part II. Lizards and amphisbaenians. Smithsonian Institution Press, Washington, D.C. & London.
40. Pyron, R.A., Burbrink, F.T., Colli, G.R., de Oca, A.N.M., Vitt, L.J., Kuczynski, C.A. & Wiens, J.J. 2011. The phylogeny of advanced snakes (Colubroidea), with discovery of a new subfamily and comparison of support methods for likelihood trees. Molecular Phylogenetics and Evolution 58: 329-342.
41. Rage, J.-C. & Augé, M. 2010. Squamate reptiles from the middle Eocene of Lissieu (France). A landmark in the middle Eocene of Europe. Geobios 43: 253-268.
42. Rodríguez Loredo, C. 1998. Estudio arqueozoológico del sitio Inca Potrero Chaquiago, Barrios La Solana y Retambay, Andalgalá provincia de Catamarca, Argentina. Relaciones de la Sociedad Argentina de Antropología 22-23: 203-245.
43. Savage, J.M.; Lips, K.R. & Ibáñez D.R. 2008. A new species of Celestus from west-central Panama, with consideration of the status of the genera of the Anguidae: Diploglossinae (Squamata). Revista de Biología Tropical 56: 845-859.
44. Schulte II, J. A.; Valladares, J. P. & Larson, A. 2003. Phylogenetic relationships within Iguanidae inferred using molecular and morphological data and a phylogenetic taxonomy of iguanian lizards. Herpetologica 59: 399-419.
45. Scanferla, C.A. 2006. The oldest record of Clelia (Serpentes-Colubridae) in South America. Comptes Rendus Palevol 5: 721-724.
46. Scanferla, C.A.; Cenizo, M.M. & de los Reyes, L.M. 2005. Sobre el primer registro fósil del género Lystrophis Cope, 1885 (Serpentes-Colubridae-Xenodontinae). Studia Geologica Salmanticensia 41: 93-101.
47. Scanferla, C.A.; Agnolín, F.L. & Voglino, D. 2009. Boiruna cf. B. maculata (Ophidia: Colubroides) from the early to middle Pleistocene of South America, and the effects of Pleistocene extinctions on South American reptiles. South American Journal of Herpetology 4: 259-267.
48. Strahm, M.H. & Schwartz, A. 1977. Osteoderms in the anguid lizard subfamily Diploglossinae and their taxonomic importance. Biotropica 9: 58-72.
49. Taglioretti, M.L.; Scaglia, F.A. & Isla, F.I. 2009. Tafonomía y estratigrafía del yacimiento paleontológico "Constitución", Pleistoceno medio-superior, Mar del Plata, Argentina. Ameghiniana, 46: 94.
50. Tonni, E.P.; Pardiñas, U.F.J.; Verzi, D.H.; Noriega, J.I.; Scaglia, O. & Dondas, A. 1998. Microvertebrados pleistocénicos del sudeste de la provincia de Buenos Aires (Argentina): Bioestratigrafía y Paleoambientes. V Jornadas Geológicas y Geofísicas Bonaerenses 1: 73-83.
51. Verzi, D.H.; Tonni, E.P.; Scaglia, O.A. & San Cristobal, J.O. 2002. The fossil record of the desert adapted South American rodent Tympanoctomys (Rodentia, Octodontidae). Paleoenvironmental and biogeographic significance. Palaeogeography, Palaeoclimatology, Palaeoecology 179: 149-158.
52. Verzi, D.H.; Deschamps, C.M. & Tonni, E.P. 2004. Biostratigraphic and paleoclimatic meaning of the Middle Pleistocene South American rodent Ctenomys kraglievichi (Caviomorpha, Octodontidae). Palaeogeography, Palaeoclimatology, Palaeocology 212: 315-329.
53. Vucetich, M.G.; Verzi, D.H.; & Tonni, E.P. 1997. Paleoclimatic implications of the presence of Clyomys (Rodentia, Echimyidae) in the Pleistocene of central Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 128: 207-214.
54. Zaher, H. 1999. Hemipenial morphology of the South American xenodontine snakes: with a proposal for a monophyletic Xenodontinae and a reappraisal of colubroid hemipenes. Bulletin of the American Museum of Natural History 240: 1-168.
55. Zaher, H., Grazziotin, F.G., Cadle, J.E., Murphy, R.W., de Moura-Leite, J.C. & Bonatto, S. L. 2009. Molecular phylogeny of advanced snakes (Serpentes, Caenophidia) with an emphasis on South American Xenodontines: a revised classification and descriptions of new taxa. Papéis Avulsos de Zoologia 49: 115-153.

