










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkKurtziana
On-line version ISSN 1852-5962
Kurtziana vol.32 no.1-2 Córdoba Jan./Dec. 2006
ANATOMÍA Y MORFOLOGÍA
Morfología y anatomía floral en especies de Apocynaceae-Asclepiadoidea
Leonardo Galetto*
* "Grupo de Investigación en Ecología Reproductiva de Plantas". Instituto Multidisciplinario de Biología Vegetal (IMBIV). CONICET-Universidad Nacional de Córdoba y Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Casilla de Correo 495, 5000 Córdoba, Argentina. E-mail: leo@imbiv.unc.edu.ar. Miembro de la Carrera del Investigador Científico. CONICET.
Resumen
Se estudió la morfología y anatomía floral de cuatro especies de Apocynaceae - Asclepiadoideae: Asclepias flava, Funastrum gracile, Morrenia odorata y Philibertia gilliesii, prestando especial atención al nectario. Las flores son pentámeras, con gineceo bicarpelar, dos estilos y una cabeza estigmática adnata al androceo que en conjunto forman el ginostegio. Se observan cinco zonas estigmáticas por debajo de la cabeza estigmática, cada una dentro de una cámara estigmática que se localiza entre dos anteras adyacentes. El androceo está formado por cinco estambres con anteras biloculares y filamentos insertos en la base de la corola gamopétala formando un tubo. La corona está asociada al tubo estaminal y rodea al ginostegio. Puede formar un tubo como en Morrenia odorata, o bien dividirse en cinco apéndices individuales como en Asclepias flava. El polen se encuentra aglutinado en polinios, los que se fijan por pares a un "corpúsculo" que se ubica en la zona apical del ginostegio. Todas las especies tienen cinco zonas nectaríferas en la flor, que se relacionan con los márgenes de la base de cada antera. Cada zona secretora se ubica dentro de una cámara estigmática y consiste en una epidermis uniestratificada. En Asclepias flava y Funastrum gracile el néctar se acumula en cada apéndice de la corona, mientras que en Morrenia odorata y Philibertia gilliesii se acumula en la base de la corona que es simple.
Palabras clave: Morfología floral; Anatomía; Nectarios; Asclepiadoidea.
Abstract
Flower morphology and anatomy in species of Apocynaceae-Asclepiadoidea.
Flower morpho-anatomy and nectary structure of four species of Apocynaceae - Asclepiadoideae: Asclepias flava, Funastrum gracile, Morrenia odorata y Philibertia gilliesii, were studied. Flowers are 5-merous except of gynoecium of two carpels, with two styles and a stigma-head adnate to the androecium forming the gynostegium. Five stigmatic zones enclosed within the stigmatic chambers between adjacent anthers under the stigma head can be observed. The androecium consists of five stamens, with fused filaments forming a tube, and 2-locular anthers. The corona is related to the staminal tube and is located around the gynostegium. It can be tubular as in Morrenia odorata or it can show five-free lobes as in Asclepias flava. Pollen is gathered into pollinia which are attached by pairs to a corpuscle on the stigma head. All species have five nectariferous zones which are related to the flanks of the anther base. Each secretory region is located within a stigmatic chamber and consists of one layer of ephitelial cells. Nectar accumulates in the five cuculli in Asclepias flava and Funastrum gracile), or at the corona base in Morrenia odorata and Philibertia gilliesii.
Key words: Floral morphology; Anatomy; Nectaries; Asclepiadoidea.
Introducción
La propuesta de incluir a las Asclepiadaceae (subfamilia Asclepiadoideae) dentro de Apocynaceae se sustenta, fundamentalmente, porque los resultados de recientes análisis cladísticos con datos morfológicos y moleculares no apoyan la separación de Asclepiadaceae como grupo monofilético e independiente de las Apocynaceae (e.g., Endress and Bruyns, 2000; Liede-Schumann et al., 2005). Si se considera que se trata de una familia con muchísimos géneros y especies (ca. 355/3700 respectivamente), es poco lo que se conoce sobre la anatomía floral en relación con sus características de polinización, particularmente en plantas de nuestro país. Las particularidades anatómicas de las flores descriptas para este grupo de plantas y su relación con la polinización, resultan entre las más complejas dentro de las Angiospermas y desde Sprengel (1793) han interesado a los botánicos, siendo Asclepias L. el género que ha recibido más atención (e.g. Galil and Zeroni, 1965; Bookman, 1981; Pant et al., 1982; Kevan et al., 1989; Sage et al., 1990; Kunze, 1991, 1995, 1997). Asimismo, hay algunos antecedentes para Cynanchum L. (Christ and Schnepf, 1985) y para Funastrum E. Fourn. y Philibertia Kunth (Kunze and Liede, 1991; descriptos allí dentro de Sarcostemma). Sin embargo, otros géneros aún se encuentran inexplorados en estos aspectos, entre otros Morrenia Lindl., que posee sólo 10 especies restringidas a Sudamérica (Goyder, 2003). En Argentina crecen aproximadamente 145 especies de unos 30 géneros de Asclepiadoideae (Ezcurra, 1999). El objetivo de esta contribución es aportar nuevos datos sobre la morfología floral de esta subfamilia y considerar especialmente las particularidades anatómicas de los nectarios en cuatro especies -Asclepias flava Lillo (endémica del NO de Argentina), y tres especies nativas que crecen en Córdoba, Funastrum gracile (Decae.) Schltdl., Morrenia odorata (Hook. et Arn.) Lindl. y Philibertia gilliesii Hook. et Arn.- que se relacionan con las interacciones de polinización de estas especies.
Materiales y métodos
Todas las muestras se colectaron en Argentina y los correspondientes ejemplares de herbario se encuentran depositados en CORD. Los datos de procedencia de las mismas son los siguientes:
Asclepias flava Lillo. ARGENTINA. Prov. Salta. Dpto. Chicoana: Quebrada de Escoipe, Galetto et al. 194, 5-XII-1989 (CORD).
Funastrum gracile (Decae.) Schltdl. ARGENTINA. Prov. Córdoba. Dpto. Ischilín: Ca. Quilino, Bernardello y Galetto 724, 8-I-1990 (CORD).
Morrenia odorata (Hook. et Arn.) Lindl. ARGENTINA. Prov. Córdoba. Dpto. Ischilín: Ca. Quilino, Bernardello y Galetto 725, 8-I-1990 (CORD).
Philibertia gilliesii Hook. et Arn. ARGENTINA. Prov. Córdoba. Dpto. Santa María: Ca. Yocsina, Galetto 105, 10-XI-1989 (CORD).
El análisis de la anatomía y la estructura de los nectarios se realizó sobre flores fijadas en F.A.A. El material fue sometido a una serie de deshidrataciones con alcohol etílico-xilol y luego incluido en parafina. Los cortes longitudinales y transversales seriados de 9-12 μm de espesor se tiñeron con la triple coloración de hematoxilina, safranina y verde permanente (Conn et al., 1960). Las fotomicrografías se tomaron con película Kodak T-Max en un fotomicroscopio Zeiss II. Los esquemas florales se realizaron con la ayuda de una cámara clara. Ocasionalmente se efectuaron observaciones sobre los visitantes florales mientras se efectuaba la recolección de materiales en el campo.
Resultados
Morfología y anatomía de las flores
Las flores en estas cuatro especies son perfectas, actinomorfas y pentámeras, excepto el gineceo que es bicarpelar (Fig. 1). El cáliz es gamosépalo con tubo muy corto, profundamente partido. Asclepias flava presenta coléteres en el interior del cáliz (Fig. 1 A). Su estructura consiste en una columna rodeada por células elongadas en sentido radial; estas últimas forman una epidermis uniestratificada de aspecto secretor y poseen, externamente, una delgada cutícula. La parte basal de cada coléter presenta un adelgazamiento ya que las células epidérmicas no se encuentran totalmente desarrolladas. La corola es gamopétala (Fig. 1). El gineceo posee ovario súpero, con dos estilos y una cabeza estigmática engrosada adnata al androceo (Fig. 1 A, F). El androceo y la parte superior del gineceo constituyen un cuerpo central o ginostegio (Figs. 1 y 2). Se observan cinco zonas estigmáticas en la base de la cabeza estigmática, cada una dentro de una cámara estigmática que se localiza entre dos anteras adyacentes (Figs. 1 D y H; 2). El androceo está formado por cinco estambres con anteras biloculares (Figs. 1 C y G; 2 A, D y G) y filamentos insertos en la base de la corola (Fig. 1 A y F). Los filamentos se encuentran fusionados formando un tubo que rodea los estilos. La corona está asociada al androceo, rodea al ginostegio y presenta longitud variable, pudiendo formar un tubo como en Morrenia odorata (Fig. 1 F), o bien dividirse en cinco apéndices individuales ("cucullus") constituidos por la "capucha" y el "cuerno", como en el caso de Asclepias flava (Figs. 1 A; 2 A) o Funastrum gracile. Los márgenes laterales de cada teca se encuentran expandidos, lo cual les da aspecto de alas (Fig. 2 A, D y G), y son córneos. El polen se encuentra aglutinado en polinios, los que son sostenidos por los "brazos de transporte" que se fijan por pares a un "corpúsculo" -estas dos últimas estructuras forman el "aparato de traslación"-, y todo junto forman un "polinario" (terminología según Bookman, 1981). Los cinco corpúsculos se ubican en la zona apical del ginostegio, sobre la cabeza estigmática, por encima de cada hendidura que comunica con una cámara estigmática.
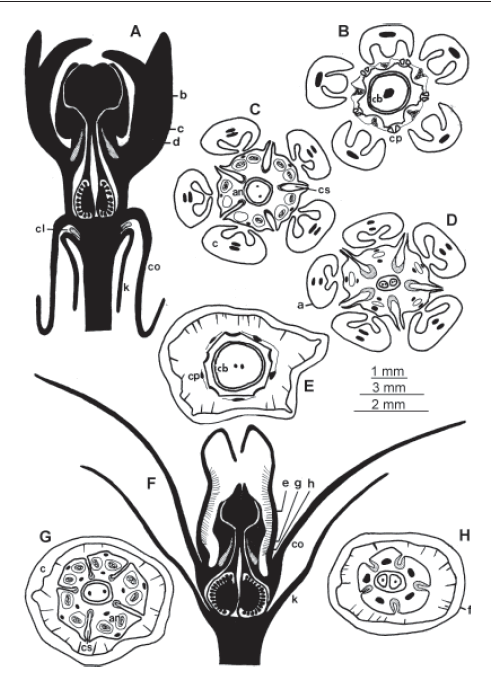
Fig. 1.- Morfo-anatomía floral en Asclepias flava y Morrenia odorata. A-D: Asclepias flava; A: esquema de corte longitudinal por flor según plano indicado en D; B-D: esquemas de cortes transversales por flor a distintas alturas según planos indicados en A, B: parte media del ginostegio, C: base del ginostegio donde se ve la región apical del epitelio nectarífero, D: base de las anteras donde se ve la zona media del tejido nectarífero. E-H: Morrenia odorata; F: esquema de corte longitudinal por flor según plano indicado en H; E, G y H: esquemas de cortes transversales por flor a distintas alturas según planos indicados en F, E: parte apical del ginostegio, G: base del ginostegio donde se ve la región apical del epitelio nectarífero, H: base de las anteras donde se encuentra la zona media del tejido nectarífero. La escala de 1 mm vale para A-D, la de 3 mm para F y la de 2 mm para el resto. Abreviaturas: an, antera; cb, cabeza estigmática; cs, cámara estigmática; co, corola; c, corona; cl, coléter; cp, ápice antera; k, cáliz. Símbolos: el grisado indica tejido nectarífero.
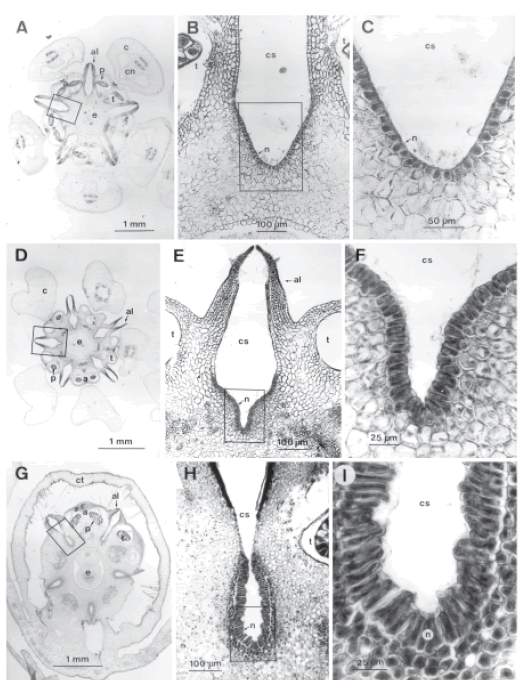
Fig. 2.- Fotomicrografías que ilustran la estructura del nectario en especies de Asclepiadoideae. A-C: Asclepias flava, A: corte transversal por flor a la altura de la corona y el ginostegio, B: detalle de una de las zonas secretoras indicada en A, C: detalle del epitelio nectarífero indicado en B; D-F: Funastrum gracile, D: corte transversal por flor a la altura de la corona y el ginostegio, E: detalle de una de las zonas secretoras indicada en D, F: detalle del epitelio nectarífero indicado en D; G-I: Morrenia odorata, G: corte transversal por flor a la altura de la corona tubular y el ginostegio, H: detalle de una de las zonas secretoras indicada en G, I: detalle del epitelio nectarífero indicado en G. Abreviaturas: a, antera; al, ala de la antera; c, corona; cn, cuerno; cs, cámara estigmática; ct, corona tubular simple; e, región estilar; n, nectario; p, polinio; t, teca.
Estructura del nectario
Todas las especies tienen cinco zonas nectaríferas en la flor, que se relacionan con los márgenes de la base de cada antera (Figs. 1 D, G, H; 2 A, D, G). Cada zona secretora se ubica dentro de una cámara estigmática (Fig. 2). El área secretora se restringe a la zona interna de esta cámara (Fig. 2 A, B, D, E, G, H) y consiste en una epidermis uniestratrificada de células bien teñidas, sin espacios intercelulares y con abundante contenido celular, grandes núcleos y numerosas vacuolas (Fig. 2 C, F, I). El tejido secretor se extiende, en sentido longitudinal, desde un poco más abajo de la zona receptiva del estigma, debajo de la cabeza estigmática, hasta el extremo inferior de la cámara estigmática (Fig. 1 A, F). El epitelio secretor se diferencia claramente del parénquima subyacente y del epitelio que recubre la parte más externa de la cámara (Fig. 2 B, E, H). En Asclepias flava y Funastrum gracile el néctar se acumula en la "capucha" de cada "cucullus" de la corona, mientras que en Morrenia odorata y Philibertia gilliesii se acumula en la base de la corona que es simple (Fig. 1 F).
Visitantes florales
Fue posible observar mariposas diurnas visitando las flores de A. flava y distintas especies de avispas en el caso de Morrenia odorata y Philibertia gilliesii. Es necesario aumentar el tiempo de observación para determinar el elenco de polinizadores que efectivamente interactúan con estas plantas.
Discusión
Existen diferentes opiniones para categorizar los distintos tipos de corona en la familia, como así también para interpretar el desarrollo ontogenético y evolutivo de esa estructura floral (e.g., Kunze and Liede, 1993; Endress and Bruyns, 2000). En cambio, si bien los nectarios en Apocynaceae s. l. se encuentran en distintas regiones de la flor (e.g., Kunze, 1991; Endress and Bruyns, 2000), no hay mayores discrepancias entre los autores en cuanto a su descripción, ubicación dentro de la flor, o relación con distintos verticilos florales. Se considera que, evolutivamente, el nectario habría migrado desde la base de la flor hacia la zona apical del filamento de los estambres (e.g., Endress and Bruyns, 2000); los primeros son típicos de Apocynoideae (i.e., 2-5 nectarios libres o bien un disco anular, siempre rodeando la base del ovario), mientras que los descriptos aquí son ejemplo de nectarios asociados al androceo característicos de Asclepiadoideae.
En las Asclepiadoideae el néctar tiene una doble función, como recompensa para los polinizadores y también como medio para favorecer la germinación del polen dentro de la cámara estigmática (e.g., Eisikovitch, 1986). El néctar es secretado dentro de la cámara estigmática por el tejido secretor. A través del seguimiento de soluciones azucaradas teñidas, fue posible determinar el camino que sigue el néctar desde la región secretora hasta la zona donde se acumula y es buscado por los polinizadores (e.g., Galil and Zeroni, 1965). En los géneros que poseen "cucullus", el néctar corre por capilaridad desde el nectario hasta esa estructura, donde se acumula (e.g., Galil and Zeroni, 1965; Kevan et al., 1989; Kunze, 1995, 1997). En el caso de Asclepias, por ejemplo, los visitantes florales buscan el néctar dentro de cada capucha de la corona, desplazándose dentro de cada flor posados sobre el ginostegio. De esta manera, las patas generalmente quedan cerca de los corpúsculos y entonces pueden pegarse polinarios a los animales y eventualmente depositar polinios dentro de cámaras estigmáticas de la misma u otra flor. En aquellos géneros sin "cucullus", el néctar también se secreta en las cámaras estigmáticas y se acumula en la base de la corona tubular.
Algunas especies de Asclepias del hemisferio norte han sido bastante estudiadas relacionando algunos aspectos de la morfo-anatomía floral, características del néctar con aspectos de la polinización (e.g., Robertson, 1886; Macior, 1965; Willson and Bertin, 1979; Bookman, 1981; Fritz and Morse, 1981; Kevan et al., 1989; Meve and Liede, 1994; Wyatt and Broyles, 1994). Sin embargo, poco se conoce sobre otros géneros de la subfamilia Apocynoideae, por lo que resultaría particularmente interesante desarrollar estudios sobre las interacciones planta-animal en este grupo de plantas con flores muy especializadas.
Agradecimientos
Agradezco a dos revisores anónimos por las constructivas sugerencias que permitieron mejorar versiones anteriores de este manuscrito, a la Secretaría de Ciencia y Técnología de la Universidad Nacional de Córdoba, a la Secretaría de Promoción Científica y Tecnológica (FONCyT) y al Consejo Nacional de Investigaciones Científicas y Técnicas por el financiamiento otorgado.
Referencias bibliográficas
1. Bookman, S. S. 1981. The floral morphology of Asclepias speciosa (Asclepiadaceae) in relation to pollination and a clarification in terminology for the genus. Amer. J. Bot. 68: 675-679. [ Links ]
2. Christ, P. & E. Schnepf. 1985. The nectarines of Cynanchum vincetoxicum (Asclepiadaceae). Israel J. Bot. 34: 79-90. [ Links ]
3. Conn, H. J., M. A. Darrow & V. M. Emmel. 1960. Staining procedures. Williams and Wilkins Co., Baltimore. [ Links ]
4. Eisikovitch, D. 1986. Morpho-ecological aspects on the pollination of Calotropis procera (Asclepiadaceae) in Israel. Pl. Syst. Evol. 152: 185-194. [ Links ]
5. Endress, M. E. & P. V. Bruyns. 2000. A revised classification of the Apocynaceae s.l. Bot. Rev. 66: 1-56. [ Links ]
6. Ezcurra, C. 1999. Asclepiadaceae, en Zuloaga, F. O. & O. Morrone (Eds.), Catálogo de las Plantas Vasculares de la República Argentina II, Syst. Bot. Missouri Bot. Gard. 74: 78-98. [ Links ]
7. Fritz, R. S. & D. H. Morse. 1981. Nectar parasitism of Asclepias syriaca by ants: Effect on nectar levels, pollinia insertion, pollinaria removal and pod production. Oecologia 50: 316-319. [ Links ]
8. Galil, J. & M. Zeroni. 1965. Nectar system of Asclepias curassavica. Bot. Gaz. 126: 144-148. [ Links ]
9. Goyder, D. J. 2003. A synopsis of Morrenia Lindl. (Apocynaceae subfam. Asclepiadoideae). Kew Bull. 58: 713-721. [ Links ]
10. Kevan, P. G., D. Eisikowitch & B. Rathwell. 1989. The role of nectar in the germination of pollen in Aclepias syriaca L. Bot. Gaz. 150: 266-270. [ Links ]
11. Kunze, H. 1991. Structure and function in asclepiad pollination. Pl. Syst. Evol. 176: 227-253. [ Links ]
12. Kunze, H. 1995. Floral morphology of some Gonolobeae (Asclepiadaceae). Bot. Jahrb. Syst. 117: 211- 238. [ Links ]
13. Kunze, H. 1997. Corona and nectar system in Asclepiadinae (Asclepiadaceae). Flora 192: 175-183. [ Links ]
14. Kunze, H. & S. Liede. 1991. Observations on pollination in Sarcostemma (Asclepiadaceae). Pl. Syst. Evol. 178: 95-105. [ Links ]
15. Kunze, H. & S. Liede. 1993. A descriptive system for corona analysis in Asclepiadaceae and Periplocaceae. Pl. Syst. Evol. 185: 275-284. [ Links ]
16. Liede-Schumann, S., A. Rapini, D. Goyder & M. W. Chase. 2005. Phylogenetics of New Word subtribes of Asclepiadaceae (Apocynaceae-Asclepiadoideae): Metastelmatinae, Oxypetalinae, and Gonolobinae. Syst. Bot. 30: 184-200. [ Links ]
17. Macior, L. Z. 1965. Insect adaptation and behavior in Asclepias pollination. Bull. Torrey Bot. Club 92: 114-126. [ Links ]
18. Meve, U. & S. Liede. 1994. Floral biology and pollination in stapeliads - New results and a literature review. Pl. Syst. Evol. 192: 99-116. [ Links ]
19. Pant, D. D., D. D. Nautiyal & S. K. Chaturvedi. 1982. Pollination ecology of some Indian asclepiads. Phytomorphology 32: 302-313. [ Links ]
20. Robertson, C. 1886. Notes on the mode of pollination of Asclepias. Bot. Gaz. 11: 262-269. [ Links ]
21. Sage, T. L., S. B. Broyles & R. Wyatt. 1990. The relationship between the five stigmatic chambers and two ovaries of milkweed (Asclepias amplexicaulis Sm.) flowers: a three-dimensional assessment. Israel J. Bot. 39: 187-196. [ Links ]
22. Sprengel, C. K. 1793. Discovery of the secret of nature in the structure and fertilization of flowers. Translated by Peter Haase, in Lloyd, G. D. & S. C. H. Barrett (eds.), Floral Biology. Studies on floral evolution in animal-pollinated plants, pp. 3-43, 1996, Chapman & Hall, New York. [ Links ]
23. Willson, M. F. & R. I. Bertin. 1979. Flower-visitors, nectar production, and inflorescence size of Asclepias syriaca. Canad. J. Bot. 57: 1380-1388. [ Links ]
24. Wyatt, R. & S. B. Broyles. 1994. Ecology and evolution of reproduction in milkweeds. Annu. Rev. Ecol. Syst. 25: 423-441. [ Links ]
Original recibido el 4 de julio de 2005;
Aceptado el 30 de agosto de 2006.

