










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkKurtziana
versión On-line ISSN 1852-5962
Kurtziana vol.32 no.1-2 Córdoba ene./dic. 2006
ECOLOGÍA
La vegetación rupícola del Parque Provincial Copahue. Neuquén, Argentina
Ricardo Gandullo * & A. Maria Faggi **
* Departamento de Biología Aplicada, Facultad Ciencias Agrarias, CC85, CP (8303), Cinco Saltos, Río Negro, Argentina, e-mail: rgandullo@ciudad.com.ar
** Museo Argentino de Ciencias Naturales Bernardino Rivadavia, A. Gallardo 470 (1405 DJR) Buenos Aires. Argentina. Universidad de Flores, Fac. de Ingeniería en Ecología, Nazca 274, 1406 Buenos Aires, Argentina.
Resumen
Se presenta un estudio florístico y ecológico de las comunidades rupícolas del Parque Provincial Copahue, ubicado en el noroeste de la Provincia de Neuquén. La vegetación fue estudiada de acuerdo a la metodología de la escuela fitosociológica de Zürich-Montpellier. Los datos de cobertura de la matriz inicial fueron clasificados con técnicas jerárquicas aglomerativas. Se reconoce la existencia de cuatro tipos de comunidades rupícolas: 1) de glerófitos en canchales estabilizados del piso bioclimático supramediterráneo; 2) de casmófítos del piso montano inferior 3) de comófitos del piso montano superior y 4) de casmófitos orófilos del piso subandino. Se presenta su ubicación sintaxonómica y se discuten aspectos estructurales, ecológicos, biogeográficos y florísticos.
Palabras clave: Fitosociología; Dendrograma; Vegetación saxícola; Parque Provincial Copahue.
Abstract
The rupicolous vegetation of the Copahue Provincial Park.
Floristical and ecological relationships of rupicolous communities were analyzed in the Copahue Provincial Park, located at the northwest of Neuquén Province. The vegetation was studied according to the methodology of the Zürich-Montpellier phytosociological school. The relevéspecies matrix was classified by means of clustering hierarchical techniques. Four rupicolous communities have been recognized at different bioclimatic belts: 1) Glerophytic on stabilized stones, located in the supramediterranean belt; 2) Chasmophytic in the lower montane one; 3) Chomophytic in the upper montane and 4) Chasmophytic orophilous in orotemperate or subandean belt. The syntaxonomical location of these communities is presented. Also structural, ecological, biogeographical and floristical features are discussed.
Key words: Phytosociology; Cluster; Saxicolous vegetation; Copahue Provincial Park.
Introducción
Es notable la dificultad inherente al estudio fitosociológico de comunidades rupícolas, especialmente para la zona altoandina. Este hecho se halla principalmente relacionado con la baja sociabilidad de las especies (Meier & Braun- Blanquet, 1934; Davies, 1951; Rivas Martínez, 1960; Houle, 1990; Mota et al., 1991; Acosta et al., 1991; Escudero & Pajarón, 1994; Funes & Cabido, 1995; Escudero, 1996; Wiser et al., 1996; Ferreyra et al., 1998) y con los problemas asociados a la identificación del óptimo ecológico de las mismas. Estas comunidades se caracterizan por desarrollarse en espacios reducidos, donde varían gradualmente las condiciones ecológicas. Por ende, otra dificultad para identificar los límites de las comunidades se debe a las condiciones de microhábitats y al tamaño del parche que ocupan, los cuales pueden variar ampliamente. Los frágiles ecosistemas rupícolas muestran variaciones discontinuas ligada a los patrones geomorfológicos. Esto implica que si el tamaño de la unidad de muestreo supera al parche de la comunidad rupícola, la identificación de la comunidad puede resultar errónea. Por lo tanto, son difíciles de muestrear porque pueden confundirse sus límites con las comunidades lindantes.
Algunos autores como Cabrera, 1976; Cabrera et al., 1977; Ower & Pérez, 1983; Roig, 1998; Ferreyra et al., 1998; Chiapella & Ezcurra, 1999a, han realizado aportes sobre comunidades rupícolas para el Distrito Altoandino Austral. Los mismos utilizan una nomenclatura muy variada; las denominaciones van desde pedregal, saxícola, rupícola, vegetación sobre laderas rocosas, del escorial y grandes bloques de basalto. Esta heterogeneidad de nombres señala la necesidad de precisar la nomenclatura a usar.
Por otro lado y hasta la fecha, no se cuenta con estudios florísticos de las comunidades del norte neuquino, desde una perspectiva fitosociológica. Este aspecto permite profundizar el conocimiento de los requerimientos ambientales de las especies, información básica que colabora en la estrategia de conservación de la biodiversidad.
El presente trabajo constituye un aporte al conocimiento de la composición florística de la vegetación rupícola del Parque Provincial Copahue. El mismo incorpora mayor información a la ya existente, ampliando la corología de los sintaxa de la Patagonia andina, precisando la nomenclatura a usar y estableciendo relaciones con los pisos bioclimáticos.
Materiales y Métodos
El estudio se llevó a cabo en el Parque Provincial Copahue, ubicado al noroeste de la Provincia de Neuquén (Departamento Ñorquin), Argentina, encuadrado entre las coordenadas 37º 47' y 37º 55' S y 70º 55' y 71º 10' O, con una superficie de 28.300 hectáreas (Fig. 1). La región de Caviahue y Copahue constituye un gran bajo intermontano de unos 250 km2 de superficie, de relieve plano a suavemente ondulado, limitado por abruptos paredones rocosos. Los principales factores modeladores del relieve han sido los procesos tectónicos holocénicos y pliocénicos con formación de extensos mantos lávicos y derrames de fondo de valle formados por basaltos olivínicos (Ferrer et al., 1990). Durante el cuaternario, la acción glaciaria modificó las geoformas de origen endógeno, erosionando las formas volcánicas y dando lugar a valles, cubetas, planicies de deposición, morenas, sin excluir depósitos coluviales pedregosos que se observan al pie del talud de las Sierras de Palos Parados y Trolón.
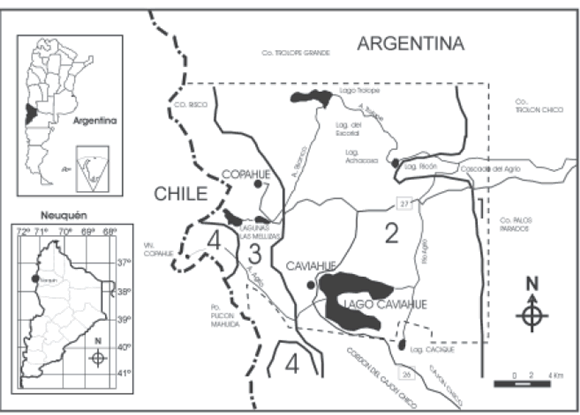
Fig. 1.- Ubicación geográfica Parque Provincial Copahue. Referencias: 1= piso supramediterráneo; 2= piso montano inferior; 3= piso montano superior; 4=piso orotemplado, (- - -) límite del Parque.
La falta de datos climáticos del área llevó a determinar los pisos bioclimáticos en un trabajo previo (Peña & Gandullo, 2003). Los mismos se establecieron de acuerdo a la propuesta de Rivas-Martínez (1993). Quedaron definidos así, dos macroclimas y tres pisos: el supramediterráneo del macroclima mediterráneo; el supratemplado o montano (inferior y superior) y el orotemplado o subandino del macroclima templado.
El trabajo de campo fue llevado a cabo durante los veranos de los a ños 1999-2002, siguiendo la metodología de la escuela fitosociológica de Zürich Montpellier (Braun- Blanquet, 1979). Se determinó un área mínima de 5 m2 para las muestras de la vegetación rupícola de acuerdo a Knapp (1984). Se realizaron un total de 37 relevamientos en unidades de muestreo florístico y geomorfológico (grietas, afloramientos rocosos verticales y/o horizontales, repisas, rocas sueltas y estabilizadas, "canchales", etc.) en rocas químicamente homogéneas. Para cada relevamiento se estimaron altitud, orientación, cobertura total de la vegetación y pendiente. Se definió además una escala cualitativa para el tipo de sustrato (basaltos olivínicos de carácter básico) de acuerdo al estado de disgregación de la roca: R1. Afloramientos rocosos con cubiertas > del 90% del área (roca continua); R2. Fragmentos rocosos > de 60 cm de diámetro (bloques); R3. Fragmentos > 25 a 60 cm de diámetro (piedras); R4. Fragmentos de 0,2 a 25 cm de diámetro (gravas + guijarros); R5. Gravilla y arena < a 0,2 cm.
La tabla fitosociol ógica inicial fue tratada con técnicas de agrupamiento jerárquico mediante el método de la mínima varianza (Ward, 1963; Orloci, 1967; 1978; Orloci & Kenkel, 1985), usando la distancia euclidiana como parámetro de disimilitud (Seber, 1984; Cuadras 1981; Anderberg, 1973). Para ello se utilizó el programa PCORD (Mc Cune & Mefford, 1999).
El análisis de inventarios florísticos (Tabla 1), en los cuales cada especie fue valorada con grados de presencia, permitió delimitar las comunidades. La flora rupícola se clasificó de acuerdo a Alcaraz Ariza (1999) en:
Tabla I: VEGETACIÓN RUPÍCOLA DEL PARQUE PROVINCIAL COPAHUE


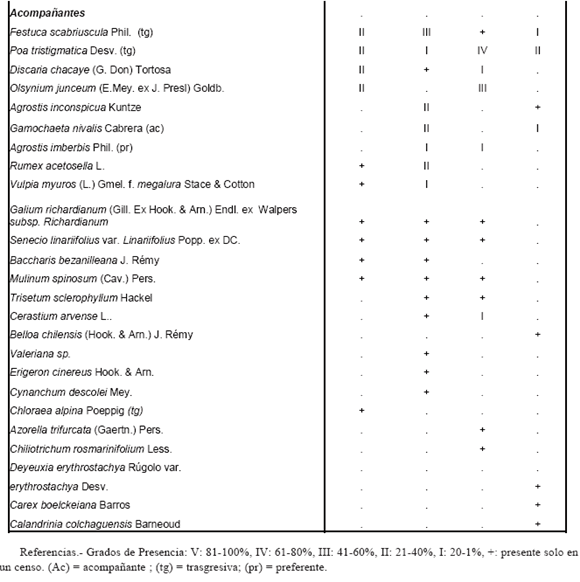
Casmófitos: plantas de los roquedales, en cuyas finas grietas hincan sus raíces.
Comófitos: plantas cuyas raíces crecen en lugares con acumulación de material suelto dentro de fisuras o grietas de rocas más o menos anchas.
Glerófitos: plantas que crecen en canchales, pedreras y gleras.
La identificación de los taxa se realizó mediante la Flora Patagónica (Correa, 1969, 1971, 1978, 1984a, 1984b, 1988, 1998, 1999) y la nomenclatura fue revisada de acuerdo con Zuloaga et al., (1994) y Zuloaga & Morrone (1996, 1999).
Resultados y Discusión
Se registraron 92 especies: 65 dicotiledóneas, 21 monocotiledóneas, 5 pteridófitas y 1 gimnosperma.
Entre las plantas con frecuencia superior al 80 % se destacan: Fabiana imbricata, Haplopappus prunelloides var. prunelloides, Gaultheria pumilia, Senecio poeppigii var. poeppigii; Rhodophiala andicola, Hypochaeris tenuifolia var. tenuifolia.
El dendrograma de la Fig. 2 permite diferenciar cuatro grupos de relevamientos con similitud decreciente. Cada agrupamiento posee una composición florística particular representada en la Tabla 1 por las comunidades de 1) glerófitos, 2) casmófitos 3) comófitos y 4) casmófitos orófilos, las cuales sirvieron de base para la interpretación sintaxonómica. Estas comunidades responden a un claro gradiente altitudinal y climático donde la comunidad 1 se ubica en el piso bioclimático supramediterráneo, la 2 en el piso montano inferior, la 3 en el piso montano superior mientras que la comunidad 4 se desarrolla en el piso orotemplado o subandino. Las comunidades 3 y 4 mantienen una relación florística a través de Gaultheria pumila, la cual es un elemento común con alta plasticidad de adaptación a ambos pisos bioclimáticos. Mientras que las comunidades 1 y 2 comparten elementos florísticos con óptimo ecológico por debajo de los 1900 m.s.m., como son Berberis empetrifolia y Baccharis magellanica.

Fig. 2.- Dendrograma de la vegetación rupícola del Parque (37 relevamientos). 1. comunidad de glerófitos con Fabiana imbricata 2. comunidad de casmófitos con Haplopappus prunelloides 3. comunidad de comófitos con Rhodophiala andicola y 4. comunidad de casmófitos orófilos con Senecio poeppigii.
Descripción de las comunidades
Comunidad de glerófitos (1): representa la comunidad rupícola del piso supramediterráneo, ubicada en la base del Cerro Palos Parados. Se caracteriza por la presencia de Fabiana imbricata en sitios con grandes rocas (canchal) semimóviles a estabilizadas, de dimensiones variables, al pie de paredes basálticas con abundante acumulación de sedimentos entre los espacios interbloques. Ocupa el rango altitudinal más bajo del Parque desde 1350 hasta 1500 m.s.m., en laderas con exposición E, NW y N, en sitios siempre protegidos de umbrías, sobre pendientes moderadas a pronunciadas, que oscilan entre 5º a 50º. La comunidad presenta sitios con edafogénesis incipiente, de origen eólico y/o coluvial. Esto permite una mayor estabilidad térmica e hídrica del hábitat, con el establecimiento de numerosas gramíneas y arbustos rastreros (Tabla 1).
Comunidad de casmófitos (2): se caracteriza por la presencia del caméfito rastrero Haplopappus prunelloides var. prunelloides. El grupo de especies que la acompañan (Tabla 1) crece entre las fisuras de las rocas macizas y con grietas de ancho variable (0,5 a 3 cm). Ocupa preferentemente los sitios de solana en el rango altitudinal de 1550 a 1750 m.s.m. El agrupamiento de estos 14 relevamientos, constituye la comunidad de casmófitos del piso montano inferior, donde el estado de disgregación de la roca es casi nulo, representado por afloramientos rocosos con más del 90% de roca masiva. Este grupo se subdivide en dos subgrupos en función de la pendiente del microhábitat. Por una parte, los relevamientos 6, 10, 11, 14, 12, 15, 18 y 13 nuclean los casmófitos que crecen entre las grandes grietas (2-3 cm) originadas por las rocas diaclasadas en llanos de muy poca pendiente (0-5º). Esto permite una relativa acumulación material suelto y fino en las fisuras. La otra situación, representada por los relevamientos 7, 8, 9, 16, 17 y 19 se da sobre repisas de acumulación de la zona vertical de las paredes rocosas y grietas (0,5-1 cm) originadas por los grandes desprendimientos de bloques basálticos columnar.
En ambos casos, colonizan casmófitos ubicuos como: Haplopappus prunelloides y Berberis empetrifolia. Las plantas que acompañan presentan particularidades morfológicas que les permiten permanecer en estos ambientes, como ser rizomas: Cheilanthes glauca, Adiantum chilense var. subsulfureum, Polystichum andinum. Otras especies poseen hojas resinosas: Haplopappus prunelloides, Baccharis magellanica, hojas reducidas y suculentas: Berberis empetrifolia y gramíneas xerófilas: Agrostis incospicua y A. imberbis, con el ingreso de elementos florísticos de la estepa circundante como son Festuca scabriuscula y Poa tristigmatica. Éstas dos últimas poseen un comportamiento transgresivo y sólo aparecen con la acumulación de material fino suelto entre las fisuras rocosas. La comunidad de casmófitos no pudo separarse según la fisura de las rocas y posición horizontal o vertical de las mismas. Otros factores, relativos a las condiciones climáticas del piso ubicado entre los 1550 a 1750 m.s.m., deben jugar un papel importante.
Comunidad de comófitos (3): es la comunidad herbácea del piso montano superior. Crece sobre las formaciones basálticas (coladas de lava, lahares y rocas meteorizadas) en pequeñas terrazas escalonadas a modo de artesas irregulares entre los roquedales macizos. Allí se acumulan materiales finos meteorizados y depósitos de ceniza volcánica, garantizando un mejor almacenaje de humedad y de materia orgánica. Estas características permiten el desarrollo de un amplio espectro de especies forrajeras y saxícolas altoandinas (1900-2300 m.s.m.) que enriquecen notablemente esta comunidad con una cobertura relativamente alta. Sin embargo, también se observan materiales más gruesos (guijarros y/o pedregales, lajas y lapilli) con escaso material suelto, más someros y de mayor drenaje. La menor disponibilidad de agua puede condicionar algunas diferencias florísticas con la situación anterior, no fáciles de dilucidar. Estos últimos ambientes son colonizados por comófitos más especializados con una cobertura bajísima, como por ejemplo Nassauvia revoluta, N. digitata y Viola cotyledon entre otras.
Por lo tanto, la comunidad de comofíticos es florísticamente más heterogénea y de una gran riqueza específica. Se caracteriza por la abundancia de Rhodophiala andicola secundada, en la mayoría de los relevamientos, por Berberis empetrifolia, Baccharis magellanica, Discaria nana, Gaultheria pumilia e Hypochaeris tenuifolia, entre otras. Tal como se puede observar en la Tabla 1, Gaultheria pumilia y, en menor medida, Senecio poeppigii var. poeppigii, representan los elementos florísticos de transición hacia la comunidad casmofítica y orófila.
Comunidad de casmófitos orófilos (4): esta comunidad, de gran homogeneidad florística y ecológica, se ubica en el piso orotemplado o subandino sobre rocas basálticas agrietadas por la meteorización, constituyendo el límite superior de la vegetación entre los 2300 hasta, aproximadamente, los 2750 m.s.m. Está conformada por micro-nanofanerófitos esparcidos y de escasa cobertura. Las plantas están adaptadas a condiciones de temperaturas extremas y con ciclo vegetativo corto en lugares donde la nieve permanece por largos períodos. Dominan Gaultheria pumila, Senecio poeppigii var. poeppigii, S. aspericaulis y Rytidosperma picta. La comunidad casmofítica orófila, prefiere laderas de solana, aunque cuando se presenta en umbría, está al reparo entre los grandes bloques basálticos. Ambas situaciones presentan pendientes pronunciadas.
Riqueza de especies y Formas de vida por pisos bioclimáticos
Funes & Cabido (1995) y Frangi & Bottino (1995), observan que el gradiente altitudinal es una variable que juega un rol importante en la composición florística y riqueza total de las comunidades rupícolas. Similares conclusiones presentan Chiapella & Ezcurra (1999b) para el Parque Tromen en Neuquén. Estas conclusiones son compatibles con nuestros resultados al analizar el rango altitudinal (1350 a 2700 m.s.m.) que ocupan las comunidades estudiadas. En la Tabla 1 puede apreciarse que por encima de los 2300 m, la comunidad de casmófitos orófilos del piso orotemplado, es la de menor riqueza con 23 especies conformadas por taxa del Distrito Altoandino-Austral (Cabrera, 1976; Roig, 1998) muchas de ellas citadas por otros autores para este tipo de ambiente (Cabrera et al., 1977; Ferreyra et al., 1998; Chiapella & Ezcurra, 1999, Gandullo et al., 2001, 2004). En el extremo inferior, entre los 1350-1500 m se ubica el piso bioclimático supramediterráneo con unas 30 especies representadas por la comunidad de glerófitos.
La excepción entre los 1500 a 2300 m, la constituyen las comunidades de casmófitos y de comófitos de los pisos montanos superior e inferior con riquezas similares, de 47 y 52 especies respectivamente. En estas comunidades se asocian especies mesotérmicas de la Provincia Subantártica y Patagónica (Cabrera, 1976; Roig, 1998). Las diferencias de riqueza y composición florística encontradas entre los pisos bioclimáticos, posiblemente, puedan atribuirse al carácter transicional del área desde un punto de vista fitogeográfico y orográfico. En segundo lugar, las condiciones ambientales presentes por encima de los 2300 m.s.m., con temperaturas extremas y alta radiación (Scarpati, 1999; Scarpati & Capriolo, 1999; Gandullo et al., 2001).
Las formas de vida predominantes en las cuatro comunidades rupícolas son arbustos bajos o microfanerófitos y caméfitos reptantes o en espaldera, datos comparables con Ruthsatz (1974,1978) para el noroeste argentino. Los hemicriptófitos y geófitos son relevantes cuando las condiciones del roquedal permiten la acumulación de material suelto fino para la instalación de ellos, como es el caso de la comunidad de comófitos, ubicada en el piso montano superior. Mientras que los nanofanerófitos comienzan a ser evidentes en ambientes más benignos, en nuestro caso, en el piso bioclimático supramediterráneo. Similares resultados describen Karagiannakidou et al. (1999) en montañas de Grecia.
Fitosociología
Al analizar la Tabla 1, se observa que tres comunidades rupícolas pertenecen al macroclima templado de los pisos supratemplado (piso montano superior e inferior) y orotemplado o subandino y una al macroclima mediterráneo del piso supramediterráneo. Las mismas presentan especies como Empetrum rubrum, Perezia pilifera y Tristagma nivale con mayor plasticidad de adaptación a lo largo de todo el gradiente climático. La presencia de estos taxa permite ubicarlas en la clase Empetro-Bolacetea (Roig et al., 1985). Esta clase posee un sólo orden el Berberidio-Empetralia y la alianza Berberidio- Empetrion (Roig et al., 1985) descriptos para el sur argentino.
Si bien el orden presenta la alianza Berberidio-Empetrion (Roig et al., 1985) con las especies características y diferenciales del orden, las comunidades rupícolas del Parque, además del nanofanerófito Berberis empetrifolia, exhiben alta presencia de otra taxa diferencial el caméfito en espaldera Baccharis magellanica (Tabla 1). Posiblemente, éste sea un elemento a tener en cuenta en futuras investigaciones al ampliar el conocimiento y rango geográfico de este tipo de vegetación con la presencia y dominancia de Berberis empetrifolia y codominancia de Baccharis magellanica. Estas podrían constituir las especies características que permitan separar las comunidades rupícolas de alta montaña del norte patagónico de la alianza Berberidio-Empetrion (Roig et al., 1985) del sur argentino.
Variabilidad: por el momento, sin rango de asociación, reconocemos las comunidades de glerófitos con Fabiana imbricata; comunidad de casmófitos montanos con Haplopappus prunelloides var. prunelloides; comunidad de comófitos con Rhodophiala andicola y comunidad de casmófitos orófilos con Senecio poeppigii var. poeppigii.
Conclusiones
1. El patrón microgeomorfológico permitió distinguir claramente las comunidades rupícolas compuestas por glerófitos, casmófitos y comófitos, los cuales son elementos florísticos especializados.
2. La microgeomorfología, altitud, orientación, pendiente y disgregación de los roquedales influyen en la variabilidad observada de las comunidades rupícolas.
3. Las especies Fabiana imbricata, Haploppapus prunelloides, Rhodophiala andicola y Senecio poeppiggi son buenos bioindicadores de los pisos bioclimáticos de las comunidades rupícolas estudiadas, sin obviar la existencia de condiciones locales significativas.
4. Las comunidades rupícolas pertenecen, mayoritariamente, al macroclima templado de los pisos bioclimáticos supratemplado (piso montano superior e inferior) y orotemplado o subandino, a excepción de la comunidad de glerófitos que se encuentran en el macroclima mediterráneo del piso supramediterráneo.
5. Las comunidades rupícolas del piso supratemplado superior y orotemplado están estrechamente relacionadas ecológicamente. En todas las comunidades se puede reconocer un claro gradiente altitudinal donde existen cambios florísticos graduales.
Referencias bibliográficas
1. Acosta, A., S. Díaz & M. Cabido. 1991. Patch structure in natural grasslands of Cordoba mountains (Argentine) in relation to different rock substrates. Coenoses 6: 21-27. [ Links ]
2. Alcaraz Ariza, F. J. 1999. Manual de teoría y Práctica Geobotánica. Universidad de Murcia. [ Links ]
3. Anderberg, M. 1973. Cluster Analysis for Applications. Academic Press, New York. [ Links ]
4. Braun-Blanquet, J. 1979. Fitosociología. Bases para el estudio de las comunidades vegetales. Ed. Blume, Madrid. [ Links ]
5. Cabrera, A. L. 1976. Regiones Fitogeográficas Argentinas, en R. L. Parodi, Enciclopedia Argentina de Agricultura y Ganadería (ed. 2), 2 (1): 1-85. ACME. Buenos Aires. [ Links ]
6. Cabrera, A. L., R. Kiesling & E. Zardini. 1977. Notas sobre algunas comunidades altoandinas del NW de la Patagonia. Obra Centenario Museo de La Plata III (Bot.): 213-227. [ Links ]
7. Correa, M. N. 1969. Fl. patagónica 8 (2). Colecc. Cient. I.N.T.A., Buenos Aires. [ Links ]
8. Correa, M. N. 1971. Fl. patagónica 8 (7). Colecc. Cient. I.N.T.A., Buenos Aires. [ Links ]
9. Correa, M. N. 1978. Fl. patagónica 8 (3). Colecc. Cient. I.N.T.A., Buenos Aires. [ Links ]
10. Correa, M. N. 1984a. Fl. patagónica 8 (4a). Colecc. Cient. I.N.T.A., Buenos Aires. [ Links ]
11. Correa, M. N. 1984b. Fl. patagónica 8 (4b). Colecc. Cient. I.N.T.A., Buenos Aires. [ Links ]
12. Correa, M. N. 1988. Fl. patagónica 8 (5). Colecc. Cient. I.N.T.A., Buenos Aires. [ Links ]
13. Correa, M. N. 1998. Fl. patagónica 8 (1). Colecc. Cient. I.N.T.A., Buenos Aires. [ Links ]
14. Correa, M. N. 1999. Fl. patagónica 8 (6). Colecc. Cient. I.N.T.A., Buenos Aires. [ Links ]
15. Chiapella, L. & C. Ezcurra. 1999a. Relaciones de la flora altoandina de dos Parques Provinciales de Neuquén. Argentina. Resúmenes. XXV Jornadas Argentinas de Botánica. Mendoza. [ Links ]
16. Chiapella, L. & C. Ezcurra. . 1999b. La flora del Parque Provincial Tromen, provincia de Neuquén, Argentina. Multequina 8: 51-60. [ Links ]
17. Cuadras, C. 1981. Métodos de análisis multivariante. Eunibar Barcelona. [ Links ]
18. Davies, O. H. 1951. Cliff vegetation in the eastern mediterranean. J. Ecol. 39:63-72. [ Links ]
19. Escudero, A. & S. Pajaron. 1994. Numerical syntaxanomy of the Asplenietalia petrarchae in the Iberian Peninsula. J. Veg. Sci. 5: 205-214. [ Links ]
20. Escudero, A. & S. Pajaron. 1996. Community patterns on exposed cliffs in a Mediterranean calcareous mountain. Vegetatio 125: 99-110. [ Links ]
21. Ferrer, J. A., J. M. Mendía & J. Irisarri. 1990. Estudio Regional de Suelos de la Provincia de Neuquén, 1. CFI. Buenos Aires. [ Links ]
22. Ferreyra, M., S. Clayton & C. Ezcurra. 1998. La flora altoandina de los sectores este y oeste del Parque Nacional Nahuel Huapi, Argentina. Darwiniana 36: 65-79. [ Links ]
23. Frangi, J. L. & O. J. Bottino. 1995. Comunidades vegetales de la Sierra de la Ventana, Provincia de Buenos Aires, Argentina. Revista Fac. Agron. 71: 93-133. [ Links ]
24. Funes, G. & M. Cabido. 1995. Variabilidad local y regional de la vegetación rupícola de las Sierras Grandes de Córdoba, Argentina. Kurztiana 24: 173-188. [ Links ]
25. Gandullo, R., J. Irizarri, A. M. Faggi & O. Scarpati. 2001. Tipos de vegetación, suelos y temperatura del aire del Parque Provincial Copahue (Neuquén, Argentina). Revista Museo Argent. Ci. Nat. 3: 13-20. [ Links ]
26. O. Scarpati, E. Maletti & A. M. Faggi. 2004. Diversidad florística del Parque Provincial Copahue, Neuquén, Argentina. Bol. Soc. Argent. 39 (3-4): 265-281. [ Links ]
27. Houle, G. 1990. Species-area relationship during primary succession in granite outcrop plant communities. Amer. J. Bot. 77: 1433-1439. [ Links ]
28. Karagiannakidou, V., M. Konstantinou & K. Papademetriou. 1999. Analysis of the upper montane and the subalpine grassland flora of East Macedonia, Greece. Candollea 54: 453-472. [ Links ]
29. Knapp, R. 1984. Considerations on quantitative parameters and qualitative attributes in vegetation analysis and in phytosociological relevés, en R. Knapp (ed.), Sampling methods and taxon analysis in vegetation science. Junk Publishers, La Haya. [ Links ]
30. Mc Cune, B. & M. J. Mefford. 1999. PC-ORD Multivariate analysis of ecological data, (version 4) Oregón. [ Links ]
31. Meier, H. & J. Braun-Blanquet. 1934. Prodrome des groupements vegetaux. 2. (Clase Asplenietales rupestres Groupements rupicoles). Montpellier. [ Links ]
32. Mota, J., F. Gómez & F. Valle. 1991. Rupicolous vegetation of beltic ranges (south Spain). Vegetatio 94: 101-113. [ Links ]
33. Orloci, L. 1967. An agglomerative method for the classification of plant communities. J. Ecol. 55: 193-206. [ Links ]
34. Orloci, L. 1978. Multivariate analysis in vegetation research.. Junk Publishers. The Hague. [ Links ]
35. Orloci, L. & N. Kenkel. 1985. Introduction to Data Analysis. International Cooperative Publishing House. Maryland. [ Links ]
36. Ower, G. & C. Pérez. 1983. Estudio de la Vegetación Natural. Relevamiento 3: 1-115. [ Links ]
37. Peña, O. & R. Gandullo. 2003. Aplicación de una metodología bioclimática mediante el uso de información satelital y SIG. Parque Provincial Copahue. Boletín Geográfico 23: 51-71. [ Links ]
38. Rivas-Martínez, S. 1960. Roca. Clima y comunidades rupícolas. Sinopsis de las alianzas hispanas de Asplenietea rupestris. Anales R. Acad. Farmacia 26: 153-168. [ Links ]
39. Rivas-Martínez, S. 1993. Bases para una nueva clasificación climática de la Tierra. Folia Bot. Matritensis 10: 1-23. [ Links ]
40. Roig, F. A., J. Anchorena, O. Dollenz, A. M. Faggi & E. Méndez. 1985. Las comunidades vegetales de la Transecta Botánica de la Patagonia Austral, en O. Boelcke, D. M. Moore & F. A. Roig (eds.), Transecta Botánica de la Patagonia Austral, pp. 350-456. CONICET-Instituto de la Patagonia-Royal Society, Buenos Aires. [ Links ]
41. Roig, F. A., J. Anchorena, O. Dollenz, A. M. Faggi & E. Méndez. 1998. La Vegetación de la Patagonia, en M. N. Correa, Fl. patagónica 8 (1): 48-166. INTA, Buenos Aires. [ Links ]
42. Ruthsatz, B. 1974. Los arbustos de las estepas andinas del noroeste argentino y su uso actual. Bol. Soc. Argent. 16: 27- 45. [ Links ]
43. Ruthsatz, B. 1978. Las plantas en cojín de los semidesiertos andinos del noroeste Argentino. Darwiniana 21: 491-539. [ Links ]
44. Scarpati, O. E. 1999. Radiación global estimada en Cerrillos (Salta) y Parque Provincial Caviahue- Copahue (Neuquén). Revista Fac. Agron. 19: 257-266. [ Links ]
45. Scarpati, O. E., A. Capriolo. 1999. Cartas de temperatura en el Parque Provincial Caviahue-Copahue (Neuquén, Argentina). Diversidad y Ambiente 1: 73-77. [ Links ]
46. Seber, G. 1984. Multivariate Observations. J. Wiley & Sons, New York. [ Links ]
47. Ward, J. H. 1963. Hierarchical grouping to optimize an objective function. Amer. Statist. Ass. J. 58: 236-244. [ Links ]
48. Wiser, S. K., R. K. Peet & P. S. White. 1996. Highelevation rock outcrop vegetation of the Southern Appalachian Mountains. J. Veg. Sci. 7: 703-722. [ Links ]
49. Zuloaga, F. O., E. G. Nicora, Z. Rúgolo de Agrasar, O. Morrone, J. Pensiero & A. M. Cialdella. 1994. Catálogo de la Familia Poaceae en la República Argentina. Monogr. Syst. Bot. Missouri Bot. Gard. 47: 1-178. [ Links ]
50. Zuloaga, F. O. & O. Morrone. 1996. Catálogo de las Plantas Vasculares de la República Argentina. I. Monogr. Syst. Bot. Missouri Bot. Gard. 60: 1-323. [ Links ]
51. Zuloaga, F. O. & O. Morrone. 1999. Catálogo de las Plantas Vasculares de la República Argentina. II. Monogr. Syst. Bot. Missouri Bot. Gard. 74: 1-1269. [ Links ]
Original recibido 31 de agosto de 2005;
Aceptado el 6 de abril de 2006.

