










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkKurtziana
versión On-line ISSN 1852-5962
Kurtziana v.35 n.1 Córdoba ene./jun. 2010
ARTÍCULOS ORIGINALES
Prosopis flexuosa DC. (Fabaceae, Mimosoideae)
Juan A. Alvarez 1, 3 & Pablo E. Villagra 1, 2
1 Departamento de Dendrocronología e Historia Ambiental (IANIGLA-CCT Mendoza-CONICET), Casilla de correo 330, 5500, Mendoza, Argentina.
2 Facultad de Ciencias Agrarias, Universidad Nacional de Cuyo. Alte. Brown S/N. Chacras de Coria. 5505. Mendoza, Argentina.
3 Autor para correspondencia: jalvarez@mendoza-conicet.gov.ar
Dedicado a la memoria de Don Fidel Antonio Roig, quien inició los estudios de esta especie en el desierto de Mendoza.
Resumen
Prosopis flexuosa DC. (Fabaceae, Mimosoideae).
El objetivo de este trabajo fue elaborar una descripción detallada de Prosopis flexuosa DC. (algarrobo dulce), árbol característico de las regiones áridas argentinas. Para su elaboración se consultaron trabajos clásicos sobre esta especie como así también las últimas investigaciones publicadas tanto en artículos nacionales como internacionales. La información obtenida abarca los siguientes temas: descripción de la especie, distribución geográfica, hábitat y comunidades, hábito de crecimiento y morfología, características citogenéticas, crecimiento y fenología, ecofisiología e interacciones biológicas. Este trabajo finaliza con la relación entre los pobladores que habitan los bosques y el algarrobo, por medio del manejo de bosques de Prosopis y los usos etnobotánicos que brinda esta especie desde épocas prehispánicas.
Abstract
Prosopis flexuosa DC. (Fabaceae, Mimosoideae).
The goal of this work was to provide a detailed description of Prosopis flexuosa DC. (algarrobo dulce), a characteristic tree of the Argentine arid regions. To do so we consulted the classic literature about this species as well as the last published papers in national and international journals. The obtained information comprises the following subjects: description of the species, geographic distribution, habitat and communities, growth habit and morphology, cytogenetic characteristics, growth and phenology, ecophysiology and biological interactions. The work ends with the relation of the woods inhabitants and the algarrobo, through the management of the Prosopis woods and the ethnobotanic uses that this species has afforded since prehispanic times.
Introducción
El género Prosopis pertenece a la familia Fabaceae (subfamilia: Mimosoideae). El nombre Prosopis fue seleccionado por Linnaeus para describir P. spicigera en la India en 1767. Para Burkart (1976), esta es la especie tipo del género, denominada en la actualidad P. cineraria. El origen de la palabra Prosopis es del griego antiguo y podría significar según Perry (1988), "hacia la abundancia" (pros = hacia y Opis = diosa de la abundancia y la agricultura). Las especies de este género se encuentran ampliamente distribuidas en regiones áridas y semiáridas de las Américas, África y Asia occidental (Burkart, 1976).
Prosopis flexuosa es llamada vulgarmente «algarrobo dulce», «algarrobo negro», «árbol negro» y «lámaro», entre otros (Roig, 1987a). Fue descripta por primera vez por De Candolle en 1825. Roig (1987a) revisó la taxonomía de la especie y, posteriormente, Palacios y Brizuela (2005) lo hacen para todas las especies del género presentes en Argentina. En estos trabajos se describen tres variedades para Prosopis flexuosa: una arbórea (var. flexuosa) y dos arbustivas (var. depressa y var. fruticosa). Entre la variedad arbórea, los que poseen espinas corresponden a la forma flexuosa y los inermes o con pocas espinas corresponden a la forma subinermis (Roig, 1987a; Palacios y Brizuela, 2005). Entre los arbustos, la variedad fruticosa presenta numerosas ramas basales, erguidas y divergentes que alcanzan los 3-5 m de alto. La variedad depressa presenta ramas basales enterradas, semienterradas o reptantes y puede alcanzar los 2,5 m de altura (Roig, 1987a). La mayor parte de los trabajos sobre la biología de esta especie han sido realizados sobre P. flexuosa var. flexuosa (árboles de porte erecto hasta 10 m de altura), por lo que la información que discutiremos en este trabajo se limitará a esta variedad.
Distribución geográfica
Prosopis flexuosa se distribuye en las regiones áridas de Sudamérica (oeste árido de Argentina y centro-norte de Chile) (Roig, 1987a). En Argentina, se la encuentra en la diagonal árida al este de la cordillera de Los Andes, abarcando parte de las provincias de Salta, Tucumán, Catamarca, La Rioja, San Juan, Mendoza, San Luis, el oeste de Córdoba, la provincia de La Pampa, el suroeste de Buenos Aires y norte Río Negro (Roig, 1993a). En Chile, habita en las regiones de Copiapó (Roig, 1993a), Coquimbo y en el valle de Elqui (Burkart, 1976) (Fig. 1). Habita las provincias biogeográficas del Monte (especie arbórea característica), el oeste del Chaco seco y el sur del Espinal (Morello, 1958; Roig, 1987a; Roig, 1993a). Las poblaciones se distribuyen desde el nivel del mar (sur de Buenos Aires) hasta los 2200 m.s.m. en valles de la cordillera de los Andes (ej. Jagüe, provincia de La Rioja) (Burkart, 1976). Existen trabajos que señalan poblaciones de P. flexuosa en Bolivia, aunque no cuentan con especificación de los lugares exactos de procedencias (Antezana et al., 2000; Larrea-Alcázar et al., 2005; López, 2003). Por el contrario, otros autores la ubican solo en Argentina y Chile (Burkart, 1976; Correa, 1984; Roig, 1987a; Steibel & Troiani, 1999).
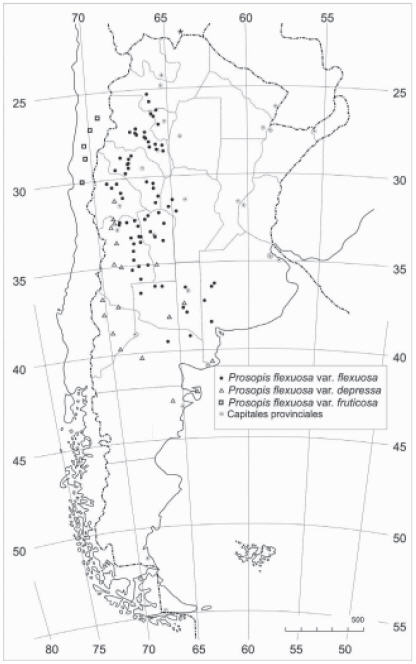
Fig. 1. Mapa de distribución de Prosopis flexuosa D.C. Variedad arbórea: Prosopis flexuosa var. flexuosa (![]() ) y variedades arbustivas: Prosopis flexuosa var. depressa (
) y variedades arbustivas: Prosopis flexuosa var. depressa (![]() ) y Prosopis flexuosa var. fruticosa (
) y Prosopis flexuosa var. fruticosa (![]() ). (Burkart 1952, Roig 1987a, b y Roig 1993b). Se ha citado esta especie para Bolivia (*) pero no se cuentan con especificación de los lugares exactos de procedencias (Antezana et al., 2000; Larrea-Alcázar et al., 2005; López, 2003).
). (Burkart 1952, Roig 1987a, b y Roig 1993b). Se ha citado esta especie para Bolivia (*) pero no se cuentan con especificación de los lugares exactos de procedencias (Antezana et al., 2000; Larrea-Alcázar et al., 2005; López, 2003).
Hábitat y comunidades
Prosopis flexuosa habita principalmente en dos tipos de hábitat: bosques en galería de los ríos y bosques de llanura (Roig et al., 1992). El rango de distribución con respecto a la precipitación anual media va desde los 50 mm (centro-sur de San Juan) hasta los 500 mm en el oeste de Córdoba. La temperatura de estas regiones varía entre los 48ºC de máxima absoluta hasta los -12ºC de mínima absoluta.
Considerando que en zonas áridas el agua es el factor que controla la productividad primaria (Noy-Meir, 1973), los bosques de algarrobo existentes en la transición entre el Chaco seco y el Monte presentan diferentes características estructurales en función de la humedad disponible. En este sentido, los representantes arbóreos se vuelven cada vez menos frecuentes hacia la zona más seca (Cabido et al., 1993). En la zona de mayor humedad de este gradiente (Reserva Chancaní, 31º 23'S y 65º 27'O), el estrato arbóreo superior está dominado por Aspidosperma quebracho-blanco seguido en menor importancia por P. flexuosa, los arbustos Larrea divaricata, Mimozyganthus carinatus y Acacia furcatispina presentan los valores más altos de abundancia-cobertura (Bonino & Araujo, 2005; Cabrera, 1976). En la zona de mayor aridez (Reserva Telteca, 32º 24'S y 68º 4'O), el bosque abierto de P. flexuosa es la formación boscosa más representativa acompañado por Geoffroea decorticans, Capparis atamisquea, Atriplex lampa y Suaeda divaricata entre otras (Alvarez et al., 2006).
Morello (1958) propone que los árboles de esta especie no pueden vivir independientemente de un suplemento de agua en zonas donde las precipitaciones anuales son menores a 350 mm. El análisis de los hábitats ocupados por esta especie fue una de las primeras evidencias que sustentan esta hipótesis. En el Monte, es la especie característica de áreas aledañas a los ríos permanentes, como el Bermejo o el Desaguadero. En las travesías, forma bosques abiertos donde existe disponibilidad hídrica por niveles freáticos próximos a la superficie (entre 6 y 12 m), comportándose como freatófita (Roig et al., 1992; Villagra et al., 2004). En suelos limosos muy profundos (con nivel continuo de agua subterránea a los 80 m de profundidad), P. flexuosa se comporta como vadosófita (bosque de Ñacuñán) (Torres & Zambrano, 2000). Una gran variedad de suelos profundos pueden brindarle condiciones aptas para su desarrollo, los mismos pueden poseer distintas granulometrías de arenas, limos o alto contenido de arcilla (Alvarez, 2002).
Donde el hombre ha transformado las áreas ocupadas por bosques, la acción combinada de agentes naturales y antrópicos ha dado lugar a procesos degradatorios del ecosistema. Esta dinámica ha conducido al bosque a diferentes grados de desertificación en diversas regiones del Monte. Los efectos más observados son cambios en la composición de las especies y la diversidad, en la dinámica del agua y las condiciones del suelo, en la estructura trófica y en la productividad de estos bosques (Villagra et al., 2009).
Hábito de crecimiento y morfología
El tamaño y la forma del árbol varían considerablemente entre poblaciones e individuos, debido a la variabilidad genética e influencias ambientales (Cony & Trione, 1996a; Duff et al., 1994). Las poblaciones norteñas (Bolsón de Fiambalá y Salar de Pipanaco en Catamarca) poseen mayor densidad de árboles con diámetros basales grandes (>35 cm), los individuos son más altos (6-12 m) y de mayor diámetro de copa (8-12 m), la cobertura arbórea, el diámetro y altura de fuste, la mayoría son erectos y unifustales y la biomasa forestal del bosque puede llegar a 42 Tn ha-1 (Fig. 2A). Las poblaciones del Monte central (Telteca, Ñacuñán y General Alvear en Mendoza) poseen menor densidad de individuos maderables, los individuos son más bajos (3-7 m), el diámetro de copa es menor (3-6 m) (< cobertura), la mayor proporción de los individuos son decumbentes y multifustales, los fustes son de menor tamaño y la biomasa varía entre 9 y 12 Tn ha-1 (Fig. 2B) (Villagra & Alvarez, 2006). En el Chaco árido, donde esta especie acompaña a Aspidosperma quebracho-blanco, la biomasa de P. flexuosa varía entre 1,6 y 5,2 Tn ha-1 (Bonino & Araujo, 2005). En un bosque con densidad promedio de 120 árboles ha-1 (Balde Viejo, Departamento de Pocho en Córdoba), Martijena y colaboradores (1988) estimaron 32 Tn ha-1 para la biomasa aérea de P. flexuosa. Este árbol es altamente heliófilo, las ramas interiores se secan formando la leña campana (Roig, 1985). Según Burkart (1976), P. flexuosa se caracteriza por sus ramas arqueadas o semipéndulas, flexuosas y nudosas, siendo las espinas geminadas y fuertes (largas o cortas). Las especies del género Prosopis poseen un tejido leñoso complejo en el cual la determinación de sus anillos de crecimiento no siempre es simple y sencilla (Castro, 1994; Villalba & Boninsegna, 1989). Como en otras especies del género, los caracteres más frecuentemente asociados con la delimitación de los anillos de crecimiento anual son la presencia de parénquima terminal y de vasos de mayor diámetro al comienzo de las bandas de crecimiento (Villalba, 1985). Para P. flexuosa la porosidad semicircular es la distribución de vasos más frecuente, siendo difusa en los estadios de la adultez (observado en el leño de los últimos años de individuos mayores de 100 años del Monte central) (observ. pers.).

Fig. 2. Prosopis flexuosa D.C. A) Individuo adulto unifustal, bosque de Fiambalá (Catamarca) (Foto de P. Villagra); B) Individuo adulto con varios fustes, bosque de Telteca (Mendoza) (Foto de J. Alvarez); C) Inflorescencias (Foto de P. Villagra) y D) Infrutescencia con frutos en desarrollo (Foto de A. Giantomasi).
Las hojas son bipinadas, con 1 a 3 yugas, glabras o muy poco pubérulas; las pinas de 5 a 13 cm de largo; los foliolos lineares de 12 a 29 por pina, opuestos, distanciados en más del ancho por el raquis, de 4 a 15 mm de largo por 1 a 2 mm de ancho, subcoriáceos (Burkart, 1976). En estudios sobre individuos renovales procedentes de Catamarca, San Juan y Mendoza realizados en huerto común, se marca la similitud morfológica en los individuos de Catamarca y San Juan con respecto a Mendoza en la mayor longitud de la hoja, del pecíolo, de la pina primaria, del folíolo, del fruto y del ápice del fruto (Brizuela et al., 2000), lo anterior indica que estas diferencias responderían a la variación genética heredable.
Las inflorescencias se agrupan en racimos densifloros de 4 a 14 cm de largo con aproximadamente 200 piezas florales cada uno, presentando cáliz y corola pubérulos. Siendo los pétalos de 3,5 mm de largo con estambres de 5 a 6 mm de longitud (Burkart, 1976) (Fig. 2C). El azúcar total por inflorescencia es 9,66 mg y el polen total por inflorescencia es 19,85 mg (Simpson et al., 1977).
El fruto es una vaina recta subfalcada de 5 a 28 cm de largo por 0,7 a 1,2 cm de ancho, subcomprimida, submoniliforme (márgenes ondulados) color amarillo apagado con manchas violeta o violeta oscuro. Las semillas son de forma ovalada, duras e impermeables, requieren altas temperaturas para germinar (Roig, 1987b) (Fig. 2D). La alta variabilidad anual en la producción de semillas puede responder a múltiples factores, se han observado pérdidas de inflorescencias en alta proporción durante eventos de lluvia y granizo (observ. pers.). Según Solbrig y Cantino (1975) menos del 3% de la producción total de inflorescencias inician el desarrollo de frutos y solo la mitad o un tercio lo completan. Dalmasso y Anconetani (1993) observaron que la producción de semillas varía en esta región entre 80.000 y 800.000 semillas por hectárea para tres años de medición. Las semillas de esta especie poseen de 32 a 36% de proteínas, 17% de grasas, 40% de hidrato de carbono, 5% de ceniza y 6% de fibra (Burkart, 1952; Burkart, 1976). Estudios realizados sobre goma extraída de las semillas presentó entre 66,1 y 72,5% de polisacáridos, estos hidrocoloides son sustancias hidrofílicas (altamente solubles o dispersables en agua) (Ibáñez & Ferrero, 2003).
Características citogenéticas
Como la mayoría de las especies de este género, P. flexuosa es diploide, con un número de cromosomas somáticos de 2n = 28 (Hunziker et al., 1975; Trenchard et al., 2008). La alta variabilidad genética encontrada en las diferentes poblaciones del Monte (también a nivel de familia), en rasgos alométricos evaluados (altura y diámetro basal), y los altos valores de heredabilidad a nivel individual, promueven su utilización en programas de mejoramiento genético (Cony & Trione, 1996a). La evaluación de la variabilidad genética de P. flexuosa se ha realizado en sus caracteres morfológicos (Mantován, 2002), fisiológicos (donde la variabilidad genética de la tolerancia al estrés hídrico y salino se manifestó a niveles de procedencias y de árboles individuales) (Cony, 1995; Cony & Trione, 1996a; Cony & Trione, 1996b; Cony & Trione, 1998; Mantován, 2002), y de crecimiento donde se observó que las procedencias del norte tienen mayor crecimiento en altura y diámetro basal que las del sur, existiendo alta heredabilidad a nivel de familias (Cony & Trione, 1996b; Cony & Trione, 1998).
Las flores son hermafroditas, protóginas, favoreciendo la fecundación cruzada. Se han observado híbridos de esta especie con P. chilensis y P. pugionata (Burkart, 1952; Roig, 1993a). La elevada capacidad de hibridación de este género determina que algunas especies como P. flexuosa, P. alba, P. nigra y P. chilensis, constituyan un grupo de especies que hibridan y se comportan como un complejo de especies que se mantiene aislado reproductivamente de otros complejos similares (Joseau et al., 2005).
Crecimiento y fenología
P. flexuosa inicia la producción de hojas a comienzos de la primavera. Como las otras especies de la sección Algarobia, la aparición del follaje nuevo es independiente de las precipitaciones. En el Monte, cuando las precipitaciones anuales están en su máximo (enero-febrero), la mayoría de las hojas están maduras (Mooney et al., 1977). En años con inviernos cálidos, los árboles pueden conservar sus hojas durante el invierno (Morello, 1958).
Para establecer la anualidad de las bandas de crecimiento leñoso, Villalba (1985) relacionó la formación del leño con las fases fenológicas de P. flexuosa en el centro del Monte, comprobando que la ciclicidad anual de la actividad cambial se encuentra fuertemente ligada al régimen climático. La actividad cambial alcanza el mayor nivel de producción de tejido leñoso en noviembre-diciembre (inicio de periodo de precipitaciones) y cae la actividad en abril. La dormancia cambial coincide con los meses fríos y secos, donde la biomasa de hojas es mínima (Villalba & Boninsegna, 1989; Villalba et al., 2000).
La presencia de anillos de crecimiento definidos (periodicidad anual), permite determinar tasas de crecimiento y que factores ecológicos lo condicionan (Villalba & Boninsegna, 1989). En estudios dendrocronológicos realizados en diferentes poblaciones de P. flexuosa, se observó que el crecimiento corriente anual es muy variable entre los distintos bosques y entre los individuos de un mismo bosque (Alvarez, 2008; Calzon Adorno, 1995; Perpiñal et al., 1995; Villagra et al., 2005a; Villagra et al., 2005b). Los valores medios de crecimiento anual disminuyen de norte a sur a lo largo del Monte entre los 25º y los 36ºS (Villagra et al., 2005b), lo que se relacionaría con temporadas de crecimiento mas largas en los bosques del norte (Fiambalá, Pipanaco) que en los del sur (Telteca, Ñacuñán, General Alvear) (Tabla 1).
Tabla 1
Incrementos anuales medios del diámetro, para P. flexuosa en algunos bosques de regiones áridas de Argentina

Se ha observado en especies de Prosopis que la presencia de múltiples fustes se debe a la destrucción del meristema apical por el accionar de animales, procesos físicos o heladas (Fisher, 1977). Alvarez (2008) comparó el crecimiento anual del leño en individuos de P. flexuosa con un fuste y multifustales encontrando un mayor incremento anual de biomasa en los individuos multifustales hasta los 60 años de edad. Sin embargo, en los multifustales la tasa de crecimiento desciende rápidamente a partir de los 60 años de edad, mientras que en los unifustales se mantiene en ascenso más allá de los 100 años analizados. Sabiendo que el patrón de ramificación de la planta afecta directamente su capacidad fotosintética y que el mismo se define en los estadíos juveniles (Fisher, 1986), la diferencia en el crecimiento anual entre unifustales y multifustales podría atribuirse a diferentes dinámicas en la capacidad de fotosíntesis a lo largo de la vida de los individuos (Alvarez, 2008). Ambos tipos de individuos analizados presentan una distribución concéntrica de los anillos de crecimiento en los primeros años, lo que sugiere que durante este período no hay interferencia entre los fustes y entre las copas de cada fuste (Villalba, 1985; Villalba & Boninsegna, 1989; Villalba et al., 2000). Entre los 15 y 60 años, la mayor superficie foliar en los pies multifustales determinaría una mayor tasa fotosintética de acuerdo a lo sugerido por Duff y colaboradores (1994). Sin embargo, cuando se supera un cierto tamaño de copa, los fustes empezarían a competir entre sí determinando la marcada disminución de la tasa de incremento de biomasa. Por el contrario, en los algarrobos de un solo fuste la capacidad fotosintética no se vería afectada por este proceso, por lo que mantiene el ritmo de incremento en el crecimiento (Alvarez, 2008).
Ecofisiología
Las semillas de P. flexuosa presentan dormición física debida a la cubierta impermeable que poseen (Catalán & Balzarini, 1992). Esta cubierta, sumada al mesocarpo nutritivo, constituye una adaptación a la dispersión endozoócora (Campos & Ojeda, 1997; Peinetti et al., 1993). Diversas técnicas de escarificación de semillas han sido desarrolladas para favorecer la germinación y posterior cultivo de esta especie, incluyendo la escarificación mecánica, química o tratamientos en agua caliente (Catalán & Macchiavelli, 1991; Vilela & Ravetta, 2001). La temperatura óptima de germinación de P. flexuosa es de 20-25ºC, siendo valores más bajos a los registrados para otras especies de este género que habitan regiones del norte del Monte como P. chilensis y P. argentina (Cony & Trione, 1996b; Villagra, 1995). Además, muestra alta capacidad de germinación bajo condiciones de salinidad y estrés hídrico en comparación con otras especies arbóreas del género, aunque una alta variabilidad intraespecífica fue observada (Cony & Trione, 1998). La tolerancia a la salinidad en la fase de plántula es mayor que durante los primeros estadios luego de la germinación, lo que sugiere como adecuada la forestación de áreas degradadas por salinización con individuos juveniles de esta especie (Catalán et al., 1994). Esta tolerancia aumenta a la largo de la ontogenia del individuo, siendo considerada una especie tolerante (Catalán et al., 1994; Cazebbone et al., 1999; Cony & Trione, 1998). La rapidez del proceso de germinación y crecimiento inicial (2-4 días para la emergencia radicular y 8-10 días para la aparición de las hojas) (Cony, 1996; Vilela & Ravetta, 2001) permiten un eficiente uso del agua durante los cortos períodos de disponibilidad hídrica en el área de distribución.
Su capacidad para sobrevivir en ambientes de extrema aridez radica principalmente en la posibilidad de alcanzar los niveles acuosos subterráneos, independizándose de las condiciones externas del ambiente desértico (Roig, 1985). En estudios recientes, se encontró que la composición isotópica del agua del xilema de esta especie es similar al del agua freática, demostrando que P. flexuosa funciona como freatófita facultativa en ambiente áridos extremos (Jobbágy et al., 2008). Estudios anatómicos del leño reflejaron la utilización de diferentes fuentes de agua a lo largo del desarrollo de la planta, con variaciones entre distintos ambientes (Villalba & Boninsegna, 1989). En estos estudios se observó que durante los años juveniles, la planta presentó un xilema con numerosos vasos pequeños y agrupados, lo que le da seguridad a la conducción del agua en condiciones de estrés hídrico, mientras que los árboles adultos, usando agua freática, presentaron una mayor proporción de vasos grandes y solitarios aumentando la eficiencia de la conducción de agua. Por el contrario, en zonas con precipitaciones mayores a 350 mm, este patrón no fue observado ya que la distribución de los vasos fue similar en todas las edades (Villalba & Boninsegna, 1989). Además, en esta región se encontró una relación directa entre las precipitaciones primavero-estivales y las características anatómicas de la madera de P. flexuosa, lo que sugiere un control de las precipitaciones sobre el crecimiento y una adaptación de la planta a la gran demanda de agua en el período primavero-estival (Giantomasi et al., 2008).
A pesar de su capacidad de explorar y obtener agua subsuperficial que le permite evitar el estrés hídrico, numerosas adaptaciones morfofisiológicas le confieren capacidad de tolerar cierto grado de déficit hídrico. Entre estas adaptaciones se ha descripto el control de la actividad estomática a bajo potencial hídrico, el plegado de las hojas para reducir la superficie de transpiración (Giordano, datos no publicados), y su capacidad de hacer ajuste osmótico (Cavagnaro & Passera, 1993). Respuestas similares al estrés hídrico se han observado en especies de Prosopis a lo largo del continente americano (Ansley et al., 1992; Nilsen et al., 1983; Ortiz et al., 1995 ; Sosebee & Wan, 1987).
La información sobre el efecto de otros factores ambientales sobre esta especie no es muy extensa. Por otra parte, si bien la irradiación no es un factor limitante en zonas áridas, la sombra disminuye la acumulación de biomasa, incrementa la relación vástago-raíz y disminuye la relación carbono-nitrógeno en hojas, lo que podría afectar la supervivencia de esta especie en ambientes estresantes (Vilela & Ravetta, 2000).
Interacciones biológicas
Prosopis flexuosa es un componente importante del hábitat para numerosas especies por lo que es considerada una especie estructuradora del sistema. Esta especie produce una gran cantidad de inflorescencias (F.A.O., 1994). Las flores producen néctar y son perfumadas, siendo visitadas por insectos de variadas especies, lo que indica que el modo primario de polinización sería biótico. En Aldalgalá (Catamarca) las especies de abejas que visitan P. flexuosa pertenecen a las familias: Colletidae, Andrenidae, Megachilidae y Anthophoridae (Simpson et al., 1977). En Mendoza, la familia Megachilidae presenta alta diversidad de especies (Durante & Cabrera, En prensa), que utilizan en su mayoría orificios de la madera muerta de Prosopis para hacer sus nidos.
Los predadores predispersivos, principalmente los brúquidos, pueden disminuir la producción de semillas entre un 25 y 70% (Kingsolver et al., 1977; Solbrig & Cantino, 1975). Entre los vertebrados, las vainas de algarrobo son consumidas en grandes cantidades por el loro barranquero (Cyanoliseus patagonus) cuando aún no han alcanzado la maduración (Roig, 1972a; Roig, 1972b).
Los árboles de P. flexuosa producen frutos muy apetecidos por granívoros, principalmente hormigas y roedores, como también por herbívoros. Mientras los primeros remueven y extraen rápidamente las semillas del sistema (Villagra et al., 2002), los herbívoros las dispersan aumentando su capacidad germinativa (Campos & Ojeda, 1997) y también disminuyen el tiempo de permanencia en el suelo, con lo cuál se reduciría la predación (Villagra et al., 2002). El efecto de los herbívoros sobre la germinación de semillas varía según las especies. Campos y Ojeda (1997) observaron que semillas de P. flexuosa provenientes de heces de maras (Dolichotis patagonum) y vacas presentaron mayor capacidad germinativa que las dispersadas por otros herbívoros. En el otro extremo se encuentra el jabalí europeo que destruye todas las semillas que consume (viabilidad cercana a cero).
En el Chaco árido el agente principal causante de los daños producidos en las ramas de P. flexuosa es Torneutes pallidipennis Reich (Coleoptera: Cerambycidae), donde la larva desarrolla galerías longitudinales ubicadas en el duramen (Coirini et al., 2001). La calidad de sitio influye en la intensidad del ataque, siendo mayor en sitios disturbados (Córdoba et al., 2000). En estudios realizados en Chancaní, se observó que a causa del ataque xilófago, el vigor de las plantas se redujo al disminuir las tasas de crecimiento leñoso y el espesor de la albura, registrándose pérdida de la biomasa leñosa (Ferrero, 2004).
P. flexuosa genera heterogeneidad espacial que modifica la distribución de las especies de los estratos arbustivos, herbáceos y de otros grupos biológicos (Rossi & Villagra, 2000; Villagra & Alvarez, 2006). Diversos mecanismos generan cambios de las condiciones microclimáticas bajo su cobertura: la moderación de temperaturas extremas, la disminución de evaporación, la redistribución de las precipitaciones y la disminución de la intensidad lumínica (Rossi, 2004). El incremento de la fertilidad del suelo, a través de la acumulación de nutrientes y los efectos físicos y químicos de la hojarasca (Alvarez et al., 2009), generan heterogeneidad espacial en las condiciones edáficas.
La materia orgánica aportada por esta especie, especialmente sus hojas, constituye un factor importante en el ciclado de nutrientes del Monte central, siendo bajo dosel de Prosopis el sitio con mayor aporte anual de mantillo (entre 320 y 527 gr m-2) (Alvarez et al., 2009). Por otro lado, es mayor la descomposición de mantillo de P. flexuosa en sitios bajo la copa de algarrobo y arbustos como la jarilla, que en áreas expuestas, siendo la descomposición de hojas de algarrobo mayor que la de otras fracciones de la planta (Miner, 2007).
Manejo de bosques de Prosopis
En árboles de zonas áridas, el crecimiento leñoso anual es relativamente bajo, siendo necesaria la aplicación de técnicas de manejo que mejoren la productividad o calidad de los productos obtenidos (Pasiecznik et al., 2001). Son incipientes las prácticas de manejo aplicadas en los algarrobales tanto de la región chaqueña Chaco árido cono en el Monte. En el Chaco seco, la presión sobre los bosques ha sido elevada (comercialización de grandes volúmenes de madera), no existiendo ordenamiento forestal ni reforestación en bosques degradados (Karlin & Díaz, 1984). En estudios realizados para los bosques nativos chaqueños, la productividad de madera del algarrobal alcanza 1 m3 ha-1 año-1 (algarrobales sin manejo silvícola) (Carranza & Ledesma, 2005). Para los algarrobales del Monte, la utilización de madera siguió un modelo de tipo extractivo (la tasa de extracción superó a la tasa de recuperación). Esto ha conducido a los algarrobales a diferentes grados de desertificación (Villagra & Alvarez, 2006). En el Monte la productividad maderable se encuentra entre 1,3 m3 ha-1year-1 en Pipanaco, 0,2 m3 ha-1year-1 en Telteca y 0,1 m3 ha-1year-1 en Ñacuñán lo que indica que el potencial económico de estos bosques varía de acuerdo a la condiciones ecológicas (Villagra et al., 2005a). El bajo crecimiento leñoso observado en el Monte central, sumado al hábito de crecimiento decumbente y la multifustalidad, restringen el uso forestal de estos bosques.
Entre las prácticas silvícolas más utilizadas en el manejo forestal de bosques de Prosopis se encuentra la poda (Patch et al., 1998). En individuos de muchos fustes, la extracción de fustes secundarios busca mejorar el hábito de crecimiento y el aumento en la cantidad y calidad de la madera producida en el largo plazo (Elfadl & Luukkanen, 2003; Patch & Felker, 1997). En el caso de P. flexuosa, la interferencia entre fustes descripta anteriormente, sugiere que la extracción de fustes podría disminuir la mismay favorecer el crecimiento de los fustes remanentes (Alvarez, 2008). En estudios recientes se observó en árboles podados en el noreste de Mendoza, la respuesta inmediata del árbol es la recuperación de la copa, aunque no se registró un claro incremento en el crecimiento leñoso del fuste remanente ni en el crecimiento en altura. Esto indicaría que la poda podría contribuir a mejorar la forma del árbol con fines forestales aunque no a aumentar la productividad, al menos durante los primeros años. Sin embargo, la generación de numerosos rebrotes luego de la poda es un problema a resolver ya que disminuye la calidad de la madera debido a la formación de nudos (Alvarez, 2008).
En zonas de baja productividad, la extracción de leña seca surge como una actividad de bajo impacto que permite complementar otras actividades productivas. En el Monte central, los árboles de P. flexuosa presentan madera muerta a partir de los 20 cm de diámetro basal (dab) (observ. pers.). En estudios realizados en el noreste de Mendoza se registró que la disponibilidad de leña seca varió entre 4 y 8 Tn ha-1 para un bosque abierto (57 individuos con dab > 20cm) y uno semidenso (80 individuos con dab > 20cm) respectivamente (Alvarez, 2008). Comparando el crecimiento radial anual entre ramas vivas y los últimos años de crecimiento de ramas secas, se observó que las ramas secas presentan un periodo de decaimiento de entre 18 y 20 años antes de la muerte de la rama (Alvarez, 2008).
Estudios de variabilidad genética intra e inter poblacional de los bosques del Monte sugieren que esta especie, a través de la selección y multiplicación de los genotipos adecuados, puede brindar ejemplares de características necesarias para usos diversos (rápido crecimiento, ausencia de espinas, resistencia a factores causantes de estrés) (Cony, 1996; Cony, 1998).
Usos etnobotánicos y comerciales
Como otras especies del género, P. flexuosa es utilizado por los pobladores del desierto desde épocas prehispánicas. En yacimientos arqueológicos de San Juan y Mendoza se han encontrado vainas de esta especie: Cueva del Toro, Las Heras (entre 1400-1100 años a.p.); Punta del Barro, Angualasto (1600 años a.p.); Agua de la Tinaja, Uspallata (4560 años a.p.) (Roig, 1993b). Como en otros yacimientos de Sudamérica, el buen estado de conservación de las vainas es posible debido a la extrema aridez de estos territorios.
La recolección del fruto (algarroba) era la actividad más importante para los pueblos que vivían en los algarrobales. Los huarpes ejercían derechos de propiedad sobre los bosques que ocupaban, pudiendo disponer del uso de los mismos. La utilización de algarroba como alimento humano se fue perdiendo al ser substituida por otros alimentos (Roig, 1993b). Sin embargo en bosques del noroeste argentino (Valle de Hualfín, Catamarca), la tradición de elaborar distintos productos de consumo humano con los frutos de algarrobo aún se conserva (el 82% de los pobladores usaba la algarroba en el pasado y en la actualidad solo el 36%) (Caparrelli 2007). Las vainas se pueden comer hervidas, dejándolas secar y moliéndolas posteriormente se pueden utilizar como harina para la elaboración del pan de algarroba o patay (Burkart, 1952; Del Castillo et al., 1990). En la primera mitad del siglo XX la producción de patay se realizaba a escala comercial con puntos de venta fuera de la región del Monte, mientras en la actualidad este producto se elabora a escala familiar (Cáceres Freyre, 1962; Capparelli, 2007). La elaboración de bebidas como la chicha, aloja o la añapa dependen de este fruto (Del Castillo et al., 1990). El uso de la vaina como forraje ha dependido de la introducción de ganado doméstico a partir del siglo XVI. De la corteza se extraen sustancias para teñir y taninos para la curtiembre (Del Castillo et al., 1990). La goma extraída de las semillas de esta especie posee características adecuadas como sustancia espesante para alimentos (Ibáñez & Ferrero, 2003).
Como con otras plantas del desierto, el uso medicinal de P. flexuosa es reconocido por varios autores (Roig, 1993b; Roig, 2001). Con las semillas molidas y fermentadas se produce una bebida con propiedades diuréticas (Roig, 2001). Otro uso de esta pasta molida es el astringente y la infusión preparada con hojas se utiliza como anticatarral (Roig, 1993b; Roig, 2001). Para el lavaje de ojos se utiliza la infusión de hojas machacadas (Roig, 2001).
En el aspecto forestal, esta especie provee a los pobladores locales de postes, leña, carbón, sombra y forraje para el ganado doméstico (Alvarez, 2002). Algunos pobladores efectúan la colecta de las vainas para alimentar sus animales, mientras que otros dejan que los animales consuman los frutos directamente desde el suelo (Caparrelli 2007). En la infraestructura del desierto, el algarrobo es el soporte obligado formando parte de las viviendas, corrales y galpones. Los implementos del hogar, muebles, herramientas, están confeccionados con esta madera (Cony & Mantován, 1998; Morlans, 1998). La extracción de agua por pozos balde es posible gracias al apuntalamiento de los mismos por medio de postes de algarrobo, evitando el desmoronamiento de los materiales finos de la zona (Alvarez, 2002).
La extracción de leña en el Monte ha estado relacionada con distintos factores según la región. Morlans (1998) describe que en Catamarca, la minería estuvo asociada al empleo de leña de algarrobos y retamos utilizados para fundición (época colonial); actualmente la explotación comercial de este recurso la realizan a pequeña escala los pobladores a través de la venta de leña y carbón. En Mendoza la extracción estuvo asociada a la llegada del ferrocarril y al afianzamiento del modelo vitivinícola a partir de 1.880, se estimó que la cantidad de hectáreas deforestadas fueron casi 200.000 (Abraham & Prieto, 1999).
Agradecimientos
Los autores quieren agradecer a María E. Soler por la traducción del resumen y al FONCyT (PICT 01222) y SECTyP - U.N. Cuyo (Programa 06/P05) por el financiamiento otorgado.
Literatura citada
1. Abraham, E. M., & M. R. Prieto. 1999. Vitivinicultura y desertificación en Mendoza, En B. García Martínez, ed. Estudios de historia y ambiente en América: Argentina, Bolivia, México, Paraguay, 109-135. IPGH - Colegio de México, México. [ Links ]
2. Alvarez, J. A. 2002. Estructura forestal y estado de conservación del bosque de algarrobos (Prosopis flexuosa) del noreste de la provincia de Mendoza y su aplicación al manejo. Tesis de grado, Universidad de Congreso, Mendoza. [ Links ]
3. Alvarez, J. A. 2008. Bases ecológicas para el manejo sustentable del bosque de algarrobos (Prosopis flexuosa DC.) en el noreste de Mendoza. Argentina. Tesis Doctoral, Universidad Nacional del Comahue, Bariloche (Argentina). [ Links ]
4. Alvarez, J. A., P. E. Villagra, B. E. Rossi, & E. Cesca. 2009. Spatial and temporal litterfall heterogeneity generated by woody species in the Central Monte desert. Plant Ecology 205: 295-303. [ Links ]
5. Alvarez, J. A., P. E. Villagra, M. A. Cony, E. Cesca, & J. A. Boninsegna. 2006. Estructura y estado de conservación de los bosques de Prosopis flexuosa DC. en el Noreste de Mendoza, Argentina. Revista Chilena de Historia Natural 79: 75-87. [ Links ]
6. Ansley, R. J., P. W. Jacoby, C. H. Meadors, & B. K. Lawrence. 1992. Soil and leaf water relations of differentially moisture-stressed honey mesquite (Prosopis glandulosa Torr.). Journal of Arid Environments 22: 147-159. [ Links ]
7. Antezana, C., M. Atahuachi, S. Arrázola, E. Fernandez, & G. Navarro. 2000. Ecología y Biogeografía del género Prosopis en Bolivia. Revista Boliviana de Ecología 8: 35-36. [ Links ]
8. Bonino, E. E., & P. Araujo. 2005. Structural differences between a primary and a secondary forest in the Argentine Dry Chaco and management implications. Forest Ecology and Management 206 (1-3): 407-412. [ Links ]
9. Brizuela, M. M., D. Burghardt, D. Tanoni, & R. A. Palacios. 2000. Estudio de la variación morfológica en tres procedencias de Prosopis flexuosa y su manifestación en cultivo bajo condiciones uniformes. Multequina 9: 7-15. [ Links ]
10. Burkart, A. 1952. Las Leguminosas Argentinas Silvestres y Cultivadas. 2 ed. Acme, Buenos Aires. [ Links ]
11. Burkart, A. 1976. A monograph of the genus Prosopis (Leguminosae subfam. Mimosoideae). Journal of the Arnold Arboretum 57: 219-249; 450-455. [ Links ]
12. Cabido, M., C. González, & S. Díaz. 1993. Vegetation changes along a precipitation gradient in Central Argentina. Vegetatio 109: 5-14. [ Links ]
13. Cabrera, A. L. 1976. Regiones Fitogeográficas Argentinas, En W. F. Kugler, ed. Enciclopedia Argentina de Agricultura y Jardinería., Vol. Tomo 2 Fascículo 1, 85pp, 2 ed. Editorial ACME, Buenos Aires. [ Links ]
14. Cáceres Freyre, J. 1962. Fabricación del «patay» en los algarrobales del Campo de Palcipas (La Rioja y Catamarca). . Instituto de la Producción, Fac. Cs. Econ., Univ. Nac. La Plata. Serie Contribuciones 76: 1-22. [ Links ]
15. Calzon Adorno, M. E. 1995. Estudio de la productividad económica de un bosque de algarrobo en el Departamento de Cafayate. Pcia de Salta. Tesis profesional, Universidad Nacional de Salta, Salta. [ Links ]
16. Campos, C. M., & R. A. Ojeda. 1997. Dispersal and germination of Prosopis flexuosa (Fabaceae) seeds by desert mammals in Argentina. Journal of Arid Environments 35: 707-714. [ Links ]
17. Capparelli, A. 2007. Los productos alimenticios derivados de Prosopis chilensis (Mol.) Stuntz y P. flexuosa DC., Fabaceae, en la vida cotidiana de los habitantes del NOA y su paralelismo con el algarrobo europeo. Kurtziana 33 (1): 103-119. [ Links ]
18. Carranza, C., & M. Ledesma. 2005. Sistemas silvopastoriles en el Chaco Árido. Revista IDIA XXI: 230-236. [ Links ]
19. Castro, M. A. 1994. Maderas Argentinas de Prosopis. Atlas Anatómico. Secretaría General de la Presidencia de la Nación., República Argentina. [ Links ]
20. Catalán, L., M. Balzarini, E. Taleisnik, R. Sereno, & U. Karlin. 1994. Effects of salinity on germination and seedling growth of Prosopis flexuosa (D.C.). Forest Ecology and Management 63: 347-357. [ Links ]
21. Catalán, L. A., & R. E. Macchiavelli. 1991. Improving germination in Prosopis flexuosa D.C. and Prosopis alba Griseb. with hot water treatments and scarification. Seed Science & Technology 19: 253- 262. [ Links ]
22. Catalán, L. A., & M. Balzarini. 1992. Improved laboratory germination conditions for several arboreal Prosopis species: P. chilensis, P. flexuosa, P. nigra, P.alba, Pcaldenia and P. affinis. Seed Science and Technology 20: 293-298. [ Links ]
23. Cavagnaro, J. B., & C. B. Passera. 1993. Relaciones hídricas de Prosopis flexuosa (algarrobo dulce) en el Monte, Argentina., En IADIZA, ed. Contribuciones Mendocinas a la Quinta Reunión de Regional para América Latina y el Caribe de la Red de Forestación del CIID. Conservación y Mejoramiento de Especies del Género Prosopis., 73-78, Mendoza, Argentina. [ Links ]
24. Cazebbone, C., A. I. Vega, D. A. Varela, & L. A. Cardemil. 1999. Salinity effects on germinations and growth of Prosopis chilensis. Revista Chilena de Historia Natural 72: 83-91. [ Links ]
25. Coirini, R., A. Córdoba, U. O. Karlin, & V. Mazzuferi. 2001. Pérdida económica producida por la infestación de Torneutes pallidipennis Reich (Coleoptera: Cerambycidae)en Prosopis aff. flexuosa del Chaco árido de Córdoba. Multequina 10: 25-34. [ Links ]
26. Cony, M. 1995. Reforestación racional de zonas áridas y semiáridas con árboles multipropósitos. Interciencia 20 (5): 249-253. [ Links ]
27. Cony, M. A. 1996. Genetic variability in Prosopis flexuosa D. C., a native tree of the Monte phytogeographic province, Argentina. Forest Ecology and Management 87: 41-49. [ Links ]
28. Cony, M. A. 1998. Importancia de los estudios fisiológicos, ecofisiológicos y genéticos sobre las especies del género Prosopis para la recuperación de ecosistemas áridos degradados de América Latina, En A. B. Cuba Salerno, et al., eds. Bosques secos y desertificación, 255-276. INRENA - Proyecto Algarrobo, Lambayeque, Perú [ Links ].
29. Cony, M. A., & S. O. Trione. 1996a. Germinación bajo estrés hídrico y salino de dos especies nativas de algarrobo. Su variabilidad genética. Actas XXI Reunión Argentina de Fisiología Vegetal. Mendoza: 130-131. [ Links ]
30. Cony, M. A., & S. O. Trione. 1996b. Germination with respect to temperature of two Argentinian Prosopis species. Journal of Arid Environments 33: 225-236. [ Links ]
31. Cony, M. A., & S. O. Trione. 1998. Inter- and intraspecific variability in Prosopis flexuosa and P. chilensis: seed germination under salt and moisture stress. Journal of Arid Environments 40 (3): 307-317. [ Links ]
32. Cony, M. A., & N. G. Mantován. 1998. Estudio integral de la región del Monte. Informe inédito: 1/44. [ Links ]
33. Córdoba, A., V. Mazzuferi, R. Coirini, & F. Casanoves. 2000. Efecto de la condición de sitio en el grado de infestación de Torneutes Pallidipennis Reich (Coleoptera: Cerambycidae) en algarrobos negros del Chaco Arido de Córdoba, Argentina. Perspectivas de manejo fitosanitario. Multequina 9: 99-110. [ Links ]
34. Correa, M. N. 1984. XVII. Prosopis. Flora Patagónica. Colección Científica INTA 4 (b): 258-267. [ Links ]
35. Dalmasso, A. D., & J. Anconetani. 1993. Productividad de frutos de Prosopis flexuosa (Leguminosae), algarrobo dulce, en Bermejo, San Juan. Multequina 2: 173-181. [ Links ]
36. Del Castillo, E. M., M. N. Gil, & C. Saravia-Toledo. 1990. El algarrobo en los Valles Calchaquíes. Desarrollo forestal participativo en Los Andes, F.A.O., Gobierno de los Países Bajos, Salta. [ Links ]
37. Duff, A. B., J. M. Meyer, C. Pollock, & P. Felker. 1994. Biomass production and diameter growth of nine half-sib families of mesquite (Prosopis glandulosa var. glandulosa) and a fast growing Prosopis alba half-sib family grown in Texas. Forest Ecology and Management 67: 257/266. [ Links ]
38. Durante, S., & N. Cabrera. En prensa. Megachilidae, En G. Debandi, et al., eds. Biodiversidad de Artrópodos Argentinos, Vol. 2. Sociedad Entomológica Argentina Ediciones, Tucumán. [ Links ]
39. Elfadl, M. A., & O. Luukkanen. 2003. Effect of pruning on Prosopis juliflora: considerations for tropical dryland agroforestry. Journal of Arid Environments 53: 441-455. [ Links ]
40. F.A.O. 1994. Consulta de expertos sobre el avance de la agroforestería en zonas áridas y semiáridas de América latina y el Caribe. [ Links ]
41. Ferrero, M. E. 2004. Efecto de la infestación de Torneutes pallidipennis Reich (Coleoptera: Cerambycidae) sobre el crecimiento leñoso de Prosopis flexuosa DC., en el Chaco Árido de Córdoba. Tesina de grado, Universidad Nacional de Córdoba, Córdoba. [ Links ]
42. Fisher, C. E. 1977. Mesquite and modern man in Southwestern North América, En B. B. Simpson, ed. Mesquite, Its biology in two desert ecosystems, 177- 188, Stroudsburg, Pennsylvania. [ Links ]
43. Fisher, J. B. 1986. Branching patterns and angles in trees, En T. J. Givnish, ed. On the economy of plant form and function, 717. Cambridge University Press, Cambridge. [ Links ]
44. Giantomasi, M. A., F. A. Roig Juñent, P. E. Villagra, & A. Srur. 2008. Annual variation and influence of climate on the ring width and wood hydrosystem of Prosopis flexuosa DC. Trees. [ Links ]
45. Hunziker, J. H., L. Poggio, C. A. Naranjo, R. A. Palacios, & A. B. Andrada. 1975. Cytogenetics of some species and natural hybrids in Prosopis (Leguminosae). Canadian Journal of Cytology 17: 253-262. [ Links ]
46. Ibáñez, C. I., & C. Ferrero. 2003. Extraction and characterization of the hydrocolloid from Prosopis flexuosa DC. seeds. Food Research International 36: 455-460. [ Links ]
47. Jobbágy, E. G., M. D. Nosetto, P. E. Villagra, & R. B. Jackson. 2008. Isótopos estables como trazadores de las fuentes de agua de bosques de algarrobo en un desierto arenoso. XXI Congreso Argentino de Ciencias del Suelo, Potrero de los Funes, San Luis. [ Links ]
48. Joseau, J., A. Verga, & M. d. P. Díaz. 2005. Los Recursos Genéticos de Prosopis. Revista IDIA XXI (8): 201- 205. [ Links ]
49. Karlin, U., & R. Díaz. 1984. Potencialidad y Manejo de Algarrobos en el Árido Subtropical Argentino. Secretaría de Ciencia y Técnica, República Argentina. [ Links ]
50. Kingsolver, J. M., C. D. Johnson, S. R. Swier, & A. L. Teran. 1977. Prosopis fruits as a resource for invertebrates, En B. B. Simpson, ed. Mesquite. Its biology in two Desert Scrub Ecosystems, 108-122. U.S./ibp synthesis series 4. Dowden, Hutchinson & Ross, Inc. [ Links ]
51. Larrea-Alcázar, D. M., R. P. López, & D. Barrientos. 2005. Efecto nodriza de Prosopis flexuosa DC. (Leg- Mim) en un valle seco de Los Andes Bolivianos. Ecotrópicos 18 (2): 89-95. [ Links ]
52. López, R. P. 2003. Diversidad florística y endemismo de los valles secos bolivianos. Ecología en Bolivia 38 (1): 27-60. [ Links ]
53. Mantován, N. g. 2002. Early growth differentiation among Prosopis flexuosa D.C provenances from the Monte phytogeographic province, Argentina. New Forests 23: 19-30. [ Links ]
54. Martijena, N., R. Coirini, L. De Monte, & L. Pietrarelli. 1988. Estimación de la producción económica de un bosque de algarrobo. (ed.). VI Congreso Forestal Argentino, Santiago del Estero. [ Links ]
55. Miner. 2007. Descomposición de materia orgánica en diferentes parches de vegetación de algarrobales del Monte, Reserva de Ñacuñán. Mendoza, Universidad de Congreso, Mendoza. [ Links ]
56. Mooney, H. A., B. B. Simpson, & O. T. Solbrig. 1977. Phenology, morphology, physiology, En B. B. Simpson, ed. Mesquite. Its biology in two Desert Scrub Ecosystems, 26-43. U.S./ibp synthesis series 4. Dowden, Hutchinson & Ross, Inc. [ Links ]
57. Morello, J. 1958. La Provincia Fitogeográfica del Monte. Opera Lilloana 2: 5-115. [ Links ]
58. Morlans, M. C. 1998. Diagnóstico y valoración ambiental del Bolsón de Pipanaco (Catamarca, Argentina): Bases para el ordenamiento del espacio. Tesís de Maestría, Universidad Internacional de Andalucía, Huelva (España). [ Links ]
59. Nilsen, E. T., M. R. Sharifi, P. W. Rundel, W. M. Jarrell, & R. A. Virginia. 1983. Diurnal and seasonal water relations of the desert phreatophyte Prosopis glandulosa (Honey mesquite) in the Sonoran Desert of California. Ecology 64 (6): 1381-1393. [ Links ]
60. Noy-Meir, I. 1973. Desert Ecosystems: Environment and Producers. Annual Review of Ecology and Systematics 4: 25-51. [ Links ]
61. Ortiz, C., L. Bravo, M. Pinto, & L. Cardemil. 1995 Physiological and molecular responses of Prosopis chilensis under field and simulation conditions. Phytochemistry 40: 1375-1382. [ Links ]
62. Palacios, R. A., & M. M. Brizuela. 2005. Fabaceae, parte 13. Subfam. II. Mimosoideae, parte 4. Tribu VI. Mimoseae, parte B. Prosopis L, En A. M. Anton and F. O. Zuloaga, eds. Fl. Fanerog. Argent., Vol. 92, 3- 25. [ Links ]
63. Pasiecznik, N. M., P. Felker, P. J. C. Harris, L. N. Harsh, G. Cruz, J. C. Tewari, K. Cadoret, & L. J. Maldonado. 2001. The Prosopis juliflora - Prosopis pallida complex: a monograph. HDRA, Coventry. [ Links ]
64. Patch, N. L., & P. Felker. 1997. Influence of silvicultural treatments on growth of mature mesquite (Prosopis glandulosa var. glandulosa) nine years after initiation. Forest Ecology and Management 94 (1-3): 37-46. [ Links ]
65. Patch, N. L., D. Geesing, & P. Felker. 1998. Suppression of resprouting in pruned mesquite (Prosopis glandulosa var glandulosa) saplings with chemical or physical barrier treatments. Forest Ecology and Management 112 (1-2): 23-29. [ Links ]
66. Peinetti, R., M. Pereyra, A. Kin, & A. Sosa. 1993. Effects of cattle ingestion on viability and germination rate of caldén (Prosopis caldenia) seeds. Journal of Range Management 46: 483-486. [ Links ]
67. Perpiñal, E., M. Balzarini, L. Catalán, L. Pietrarelli, & E. Karlin. 1995. Edad de culminación del crecimiento en Prosopis flexuosa D.C. en el Chaco árido argentino. Investig. Agr.: Sist. Recur. For. 4 (1): 45-55. [ Links ]
68. Perry, G. 1988. Prosopis. Flora of Australia 12: 7-13. [ Links ]
69. Roig, F. A. 1972a. Bosquejo fisonómico de la vegetación de la provincia de Mendoza. Boletín de la Sociedad Argentina de Botánica XIII (Suplemento): 49-80. [ Links ]
70. Roig, F. A. 1985. Árboles y bosques de la región árida centro oeste de la Argentina (Provincias de Mendoza y San Juan) y sus posibilidades silvícolas, pp. 145- 188 Forestación en zonas áridas y semiáridas. Segundo encuentro regional CIID. América Latina y el Caribe. CIID., Santiago - Chile. [ Links ]
71. Roig, F. A. 1987a. Árboles y Arbustos de Prosopis flexuosa y P. alpataco. Parodiana 5 (1): 49-64. [ Links ]
72. Roig, F. A. 1987b. Los árboles indígenas de las Provincias de Mendoza y San Juan. I parte. Serie Científica 31: 13-15. [ Links ]
73. Roig, F. A. 1993a. Informe Nacional para la Selección de Germoplasma en Especies del Género Prosopis de la República Argentina., En IADIZA, ed. Contribuciones Mendocinas a la Quinta Reunión de Regional para América Latina y el Caribe de la Red de Forestación del CIID. Conservación y Mejoramiento de Especies del Género Prosopis., 1-36. IADIZA-CRICYT-CIID, Mendoza, Argentina. [ Links ]
74. Roig, F. A. 1993b. Aportes a la etnobotánica del género Prosopis, En IADIZA, ed. Contribuciones Mendocinas a la Quinta Reunión de Regional para América Latina y el Caribe de la Red de Forestación del CIID. Conservación y Mejoramiento de Especies del Género Prosopis., 99-119, Mendoza, Argentina. [ Links ]
75. Roig, F. A. 2001. Flora medicinal mendocina. Las plantas medicinales y aromáticas de la provincia de Mendoza (Argentina). (Aborígenes, exóticas espontáneas o naturalizadas y cultivadas) EDIUNC, Mendoza. [ Links ]
76. Roig, F. A., A. Berra, M. González Loyarte, E. Martínez Carretero, & C. Wuilloud. 1992. La Travesía de Guanacache, tierra forestal. Multequina 1: 83-91. [ Links ]
77. Roig, V. G. 1972b. Esbozo General del poblamiento animal en la Provincia de Mendoza. Boletín de la sociedad Argentina de Botánica. XIII (Suplemento): 81-88. [ Links ]
78. Rossi, B. E. 2004. Flora y vegetación de la Reserva de Biosfera de Ñacuñán después de 25 años de clausura. Heterogeneidad espacial a distintas escalas. Tesis doctoral, Universidad Nacional de Cuyo, Mendoza. [ Links ]
79. Rossi, B. E., & P. E. Villagra. 2000. Efecto de la cobertura de Prosopis flexuosa sobre la composición de los estratos arbustivos y herbáceos en el Monte Central (Argentina), En P. Jiménez, et al., eds. Memorias del IV Congreso Latinoamericano de Ecología. Ecología y Desarrollo Sostenible: Reto de América Latina para el Tercer Milenio, 163-165. Instituto Regional de Ciencias Ambientales - UNESCO, Arequipa, Perú [ Links ].
80. Simpson, B. B., J. L. Neff, & A. R. Moldenke. 1977. Prosopis flowers as a resource, En B. B. Simpson, ed. Mesquite. Its biology in two Desert Scrub Ecosystems, 84-105. US/IBP Synthesis Series 4. Dowden, Hutchinson & Ross, Inc. [ Links ]
81. Solbrig, O. T., & P. D. Cantino. 1975. Reproductive adaptations in Prosopis (Leguminosae, Mimosoideae). Journal of the Arnold Arboretum 56 (2): 185-210. [ Links ]
82. Sosebee, R. E., & C. Wan. 1987. Plant ecophysiology: a case study of honey mesquite. A. Wallace, et al. (ed.). Symposium on Shrub Ecophysiology and Biotechnology., Logan, Utah. [ Links ]
83. Steibel, P. E., & H. O. Troiani. 1999. El género Prosopis (Leguminosae) en la provincia de La Pampa (República Argentina). Revista de la Facultad de Agronomía (La Pampa): 25-48. [ Links ]
84. Torres, E., & J. Zambrano. 2000. Hidrogeología de la Provincia de Mendoza, En E. M. Abraham and F. Rodríguez Martínez, eds. Argentina. Recurso y problemas ambientales de la zona árida. Provincias de Mendoza, San Juan y La Rioja, 49-58. Junta de Gobierno de Andalucía - Universidades y Centros de Investigación de la Región Andina Argentina, Mendoza. [ Links ]
85. Trenchard, L. J., P. J. C. Harris, S. J. Smith, & N. M. Pasiecznik. 2008. A review of ploidy in the genus Prosopis (Leguminosae). Botanical Journal of the Linnean Society 156 (3): 425 - 438. [ Links ]
86. Vilela, A. E., & D. A. Ravetta. 2000. The effect of radiation on seedling growth and physiology in four species of Prosopis L. (Mimosaceae). Journal of Arid Environments 44: 415-423. [ Links ]
87. Vilela, A. E., & D. A. Ravetta. 2001. The effect of seed scarification and soil-media on germination growth, storage, and survival of seedlings of five species of Prosopis L. (Mimosaceae). Journal of Arid Environments 48: 171-184. [ Links ]
88. Villagra, P. E. 1995. Temperature effects on germination of Prosopis argentina and P. alpataco (Fabaceae, Mimosoideae). Seed Science & Technology 23: 639- 646. [ Links ]
89. Villagra, P. E., & J. A. Alvarez. 2006. Algarrobo, fuente de recursos naturales. Ciencia Regional (Mendoza) 2 (4): 12-15. [ Links ]
90. Villagra, P. E., L. Marone, & M. A. Cony. 2002. Mechanism affecting the fate of Prosopis flexuosa seeds during secondary dispersal en the Monte desert. Austral Ecology 27: 416-421. [ Links ]
91. Villagra, P. E., R. Villalba, & J. A. Boninsegna. 2005a. Structure and growth rate of Prosopis flexuosa woodlands in two contrasting environments of the central Monte desert. Journal of Arid Environments 60: 187-199. [ Links ]
92. Villagra, P. E., J. A. Boninsegna, J. A. Alvarez, M. Cony, E. Cesca, & R. Villalba. 2005b. Dendroecology of Prosopis flexuosa woodlands in the Monte desert: Implications for their management. Dendrochronologia 22 (3): 209-213. [ Links ]
93. Villagra, P. E., M. A. Cony, N. G. Mantován, B. E. Rossi, M. M. González Loyarte, R. Villalba, & L. Marone. 2004. Ecología y Manejo de los algarrobales de la Provincia Fitogeográfica del Monte, En M. F. Arturi, et al., eds. Ecología y Manejo de Bosques Nativos de Argentina. Editorial Universidad Nacional de La Plata. [ Links ]
94. Villagra, P. E., G. Defossé, H. Del Valle, M. S. Tabeni, C. M. Rostagno, E. Cesca, & E. M. Abraham. 2009. Land use and disturbance effects on the dynamics of natural ecosystems of the Monte Desert. Implications for their management. Journal of Arid Environments 73: 202-211. [ Links ]
95. Villalba, R. 1985. Xylem structure and cambial activity in Prosopis flexuosa D.C. IAWA Bulletin n.s. 6 (2): 119-130. [ Links ]
96. Villalba, R., & J. A. Boninsegna. 1989. Dendrochronological studies on Prosopis flexuosa D.C. IAWA Bulletin 10 (2): 155-160. [ Links ]
97. Villalba, R., P. E. Villagra, J. A. Boninsegna, M. S. Morales, & V. Moyano. 2000. Dendroecología y dendroclimatología con especies del género Prosopis. Multequina 9 (2): 1-18. [ Links ]
Original recibido el 28 de agosto de 2009;
primera decisión: 5 de diciembre de 2009;
aceptado el 18 de diciembre de 2009.
Editor responsable: Guillermo Funes.

