










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkKurtziana
versión On-line ISSN 1852-5962
Kurtziana vol.36 no.1 Córdoba ene./jun. 2011
ARTÍCULOS ORIGINALES
Morfología profunda del gineceo en Vigna (Leguminosae, Papilionoideae)
Susana Inés Drewes 1 & Cecilia Gamba 1
1 Departamento de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales (U.B.A.). Pab. 2, 4° piso, Ciudad Universitaria. C. 1428 EHA, Buenos Aires. Argentina. e-mail: drewes@bg.fcen.uba.ar
Resumen
Se describe la morfología del gineceo en especies de Vigna nativas de Argentina, utilizando microscopía electrónica de barrido (MEB). En las flores al comienzo de la antesis, el ovario está revestido por dos tipos de tricomas, posee un disco nectarífero basal y el estilo rostrado o no, puede ser apenas curvo, con curvatura sigmoidea o con hasta cinco vueltas completas de espiralización, presenta un cepillo estilar en la porción apical introrsa, y termina en un estigma húmedo, que puede estar rodeado por tricomas. En las especies examinadas se encontraron variaciones en cuanto a: tipos de tricomas del indumento del ovario; forma del disco nectarífero basal; largo y curvatura del estilo; presencia, forma y posición del rostro; ubicación del cepillo estilar; esculturación de los tricomas del cepillo estilar; posición y forma del estigma; forma y ubicación del anillo peristigmático y disposición, longitud y esculturación de los tricomas del anillo peristigmático. Se brinda la descripción comparativa de los estados de carácter que presenta el gineceo en cada una de las especies, siendo ésta la primera observación de la esculturación de los tricomas en el género Vigna. Se discuten los resultados obtenidos con respecto a algunos aspectos de la biología floral de las especies estudiadas.
Palabras clave: Estilo; Estigma; Cepillo estilar; Anillo peristigmático; Tricomas.
Abstract
Gynoecium deep morphology in Vigna (Leguminosae, Papilionoideae).
The deep morphology of gynoecium in species of Vigna from Argentina was studied using scanning electron microscopy (SEM). At the beginning of anthesis, the ovary presents two types of trichomes; has a basal nectar disc; and the style beaked or not, can appear scarcely curved, sigmoid or curved up to five complete turns of spiraling; presents a distal stylar brush, and an apical wet stigma, which can be surrounded by trichomes. Among the species examined, variation was found in: types of trichomes of ovary indumentum; nectar disc shape; length and curvature of the style; presence, shape and position of the stylar beak; location of the stylar brush; sculpturing of the stylar brush trichomes; position and form of the stigma; form and location of the peristigmatic ring and disposition and length of its trichomes. Based on the variations in these characters, a comparative description of the gineceum of each species is provided. This is the first description of the sculptured trichomes in Vigna. The results are discussed in relation to some aspects of the floral biology of the species.
Key words: Style; Stigma; Stylar brush; Peristigmatic ring; Trichomes.
Introducción
El género Vigna Savi pertenece al complejo Phaseolus-Vigna (Maréchal et al., 1978) dentro del grupo Phaseolastrae Baudet & Maréchal (1976) el cual reúne aquellos géneros de la subtribu Phaseolinae (Phaseoleae, Papilionoideae, Leguminosae) que se caracterizan por poseer un estilo con tricomas sobre la cara interna.
Lavin & Delgado (1990) señalaron la gran variación morfológica que presenta la distribución del indumento en los estilos-estigmas de Papilionoideae, mucha de la cual es útil para diagnosticar géneros y grupos de géneros. Reconocen distintas condiciones: el estilo ciliado, el estigma ciliado o peniciliado y el cepillo polínico; que han surgido independientemente en varios taxa. En Phaseolineae la hipótesis de homología del cepillo polínico es sugerida por su coocurrencia con otros caracteres taxonómicos, tales como el epihilo (Lackey, 1981) y una inversión de 78 kb en el ADN de cloroplasto (Bruneau et al., 1990).
En otros grupos de leguminosas la morfología de la región estigmática parece proveer caracteres muy conservativos, con especies más o menos relacionadas siendo uniformes morfológicamente y presentándose diferencias más significativas por encima del nivel de género (Small & Brookes, 1983).
Las revisiones de las características del estigma realizadas en varios géneros de leguminosas indican que poseen estigmas húmedos que corresponden al grupo III (Helslop-Harrison & Shivanna, 1977; Heslop-Harrinson, 1981), con la superficie receptiva constituida por papilas pequeñas a medianas.
Estudios de morfología profunda y desarrollo del gineceo en el género Macroptilium (Benth.) Urban (perteneciente al complejo Phaseolus-Vigna), permitieron reconocer en M. fraternum (Piper) Juárez et Pérez , flores casmógamas y cleistógamas (Drewes & Hoc, 2000); y 4 tipos de estilos-estigmas presentes en las especies argentinas (Drewes, 2005).
Castro & Agulló (1998) en estudios anatómicos realizados en Vigna adenantha (G. Meyer) Maréchal, Mascherpa & Stainer indican que posee estigma húmedo pero la ubican en el grupo IV, con la superficie receptiva no papilosa y necrosada a la madurez. En el estudio del desarrollo del estilo-estigma en Vigna longifolia (Benth.) Verdc. se describe al estigma húmedo con una membrana cuticular que se desprende cuando el estigma es receptivo a la antesis (Gamba & Drewes, 2001).
En algunas especies del género se han realizado estudios de biología floral que aportan datos sobre la morfología floral pero no profundizan en los detalles que presenta el gineceo, pudiendo citarse los trabajos de Hedström & Thulin (1986) para V. vexillata (L.) A. Rich, de Gopinathan & Babu (1987) para V. minima (Roxb.) Ohwi & Ohasi, de Hoc et al. (1993) para V. longifolia, de Agulló et al. (1993) para V. luteola (Jacq.) Benth. y de Etcheverry et al. (2008) para V. caracalla (L.) Verdc.
Las especies de Vigna que se encuentran en Argentina (Hoc, 1999) poseen un potencial valor económico, como forrajeras de suelos húmedos, y otras tienen un reconocido valor ornamental (Izaguirre & Beyhaut, 1997).
En este trabajo se presenta un estudio detallado de la morfología del gineceo de siete especies de Vigna que crecen en Argentina. La descripción de la micromorfología de los tricomas representa la primera mención acerca de la variabilidad de estos en el género Vigna. Los resultados obtenidos se discuten en las especies en las que se conoce la biología floral. Se espera que a partir de los nuevos caracteres presentados, se generen hipótesis de homología, que puedan ser útiles para comprender mejor la sistemática del género.
Materiales y métodos
Se han estudiado los siguientes taxa que crecen en Argentina: Vigna adenantha (G. Mey.) Maréchal, Mascherpa & Stainier, V. candida (Vell.) Maréchal, Mascherpa & Stainier, V. caracalla (L.) Verdc., V. lasiocarpa (Benth.) Verdc., V. longifolia (Benth.) Verdc., V. luteola (Jacq.) Benth. y V. peduncularis (Kunth.) Fawc. & Rendle var. clitoroides (Benth.) Maréchal. El material estudiado se encuentra depositado en los herbarios oficiales mencionados de acuerdo con la nomenclatura de Holmgren et al. (1990) (Tabla 1).
Tabla 1. Material examinado
Para la observación de la morfología floral, se utilizaron inflorescencias conservadas en FAA y alcohol 70 % procedentes de poblaciones naturales de Vigna y de material cultivado. Se seleccionaron capullos en diferente estadio de desarrollo y flores al comienzo de la antesis, bajo el microscopio estereoscópico, se retiraron selectivamente las estructuras que impedían la visualización de las partes a examinar.
Para obtener las fotografías con microscopio electrónico de barrido (MEB), el material se deshidrató en una serie creciente de alcohol etílico (80, 90 y 100%), y luego en acetona 100%, se llevó al punto crítico y se metalizó con aleación oro-paladio.
Las flores se describen vistas desde la posición externa.
Se registró: la forma e indumento del ovario, la forma del disco nectarífero; la forma del estilo y ubicación del cepillo estilar; la esculturación y disposición de los tricomas del cepillo estilar; la rotación de la porción distal del estilo sobre su eje; la posición y forma del estigma; la presencia de anillo peristigmático; la esculturación, disposición y longitud de los tricomas del anillo peristigmático.
Resultados
Todas las especies estudiadas presentan un ovario pubescente revestido por abundantes tricomas simples, largos, lisos, de ápice ahusado que se disponen inclinados hacia el estilo; entre los mismos se encuentran tricomas glandulares de menor longitud, 4-8 celulares. En V. candida poseen un pie y una cabezuela formada por tres células alineadas. En V. longifolia la cabezuela es alargada y puede estar formada por 3 células alineadas o 2 células basales alineadas y 2 o 3 células apicales. En V. peduncularis var. clitoroides la cabezuela es globosa formada por 4 células (Fig. 1 A, B y C).
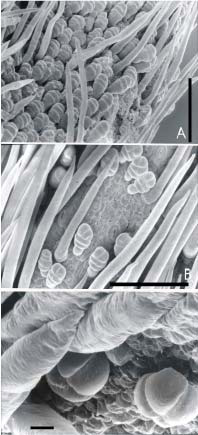
Fig. 1.- Indumento del ovario (A: V. candida, Palacios 880, B: V. peduncularis var. clitoroides, Palacios y Bravo 1294, C: V. longifolia, Palacios y Bravo 1278). La escala vale 10 μm para C y 100 μm para A y B.
En la base del ovario se ubica un disco nectarífero anular completo con el margen liso, apenas ondulado o lobulado. En V. adenantha es irregular, siendo la menor altura, donde se ubica el estambre vexilar.
El estilo en general supera ampliamente la longitud del ovario, acompaña el movimiento de la quilla en su recorrido y gira en sentido anti-horario, justo debajo del estigma. Puede presentar una prolongación más allá del estigma (rostro) y éntasis. Un cepillo de tricomas se ubica en el sector apical introrso.
En los primeros estadios del desarrollo floral el estilo es corto apenas curvo y el cepillo estilar se ubica extrorso, a medida que el estilo se elonga y curva; el cepillo estilar acompaña la torsión de este hasta su posición definitiva.
En V. luteola el estilo es algo más largo que el ovario, de grosor uniforme y la mitad distal produce medio giro en una curva abierta. En V. candida y en V. peduncularis var. clitoroides presenta el ápice hacia la derecha en una curvatura sigmoidea. En V. longifolia y en V. lasiocarpa está curvado en una espiral de 270°. En V. adenantha tiene de 4-5 cm de long., es espiralado levógiro dando 2 y ½ vueltas de giro (1000°); en V. caracalla el estilo es marcadamente largo, 7-8,5 cm de long., y efectúa de cuatro a cinco vueltas completas de espiralización (1440º).
Se diferencian dos grupos de especies, las que presentan rostro: V. candida, V. luteola, V. lasiocarpa, V. longifolia y V. peduncularis var. clitoroides y especies que no presentan rostro: V. adenantha y V. caracalla.
De las especies con estilo rostrado se distingue a Vigna candida del resto debido a que, en esta especie, el rostro se dirige hacia el mismo lado donde se dispone el estigma (en las demás especies el rostro se curva hacia la cara opuesta al estigma) (Fig. 2 A). En V. luteola, el rostro, que se curva abruptamente, se afina hacia el ápice (Fig. 2 B). V. longifolia presenta el rostro perpendicular al estilo, curvado hacia atrás y a la derecha del estigma (Fig. 2 C y D). V. lasiocarpa presenta el rostro, muy curvado hacia atrás, adopta una posición casi paralela al estilo (Fig. 2 E). En comparación con las especies descriptas, V. peduncularis var. clitoroides presenta un rostro menos desarrollado (Fig. 2 F).
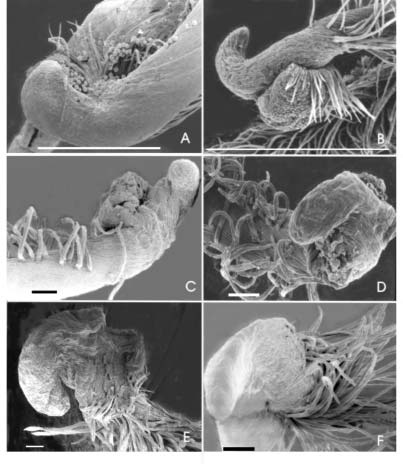
Fig. 2.- Estilos-estigmas con rostro. (A: V. candida, Palacios 880, B: V. luteola Palacios 1300, C y D: V longifolia, Palacios y Bravo 1278, E: V. lasiocarpa, Palacios y Bravo 1285, F: V. peduncularis var. clitoroides, Palacios y Bravo 1294). La escala vale 100 μm para A, B y 1000 μm para C, D, E, y F.
El estigma ocupa la porción terminal o subterminal, puede ser desde mas o menos plano hasta globoso y prominente, presenta la superficie rugosa, irregular, no papilosa, y cubierta por una membrana cuticular a la antesis (Fig. 3 B y D) y posee un anillo peristigmático incompleto que se ubica rodeando la parte basal introrsa del estigma. V. longifolia constituye la única especie de las estudiadas que no presenta tricomas rodeando al estigma. En los primeros estadios de desarrollo floral, la superficie estigmática presenta papilas, pero a medida que madura el estigma, las células necrosan y producen un exudado que queda retenido debajo de la membrana cuticular hasta que esta se rompe. Los restos de la membrana cuticular y granos de polen se observan en las flores polinizadas; en muchos casos este exudado inunda los pelos del anillo peristigmático y al mismo se adhiere el polen (Fig. 3 E y F). En V. caracalla el estigma es oblongo y ocupa una posición terminal en el estilo pero, debido a un brusco giro de este último, es presentado de forma lateral. Los tricomas del anillo peristigmático, se disponen en una o dos hileras, siendo más cortos y lisos los laterales, mientras que los centrales son más largos y levemente estriados (Fig. 3 A y C). Vigna adenantha presenta una superficie estigmática casi semicircular, en posición terminal, presenta tricomas largos y extendidos que se disponen en varias hileras ocupando el semicírculo orientado hacia el cepillo estilar. Del lado opuesto a esos tricomas, justo debajo del estigma, el estilo presenta una constricción (Fig. 3 E). V. candida, la superficie estigmática es ovalada, presenta un anillo peristigmático con largos tricomas lisos (Fig. 2 A). V. luteola presenta un estigma globoso muy prominente, el anillo estigmático se encuentra reducido a una hilera de tricomas cortos y largos, lisos, alineados en la base del estigma (Fig. 2 B). V. peduncularis var. clitoroides presenta un estigma globoso con varias hileras de largos tricomas densamente dispuestos (Fig. 2 E). V. lasiocarpa presenta un estigma globoso rodeado por tricomas cortos lisos en la base (Fig. 2 F).
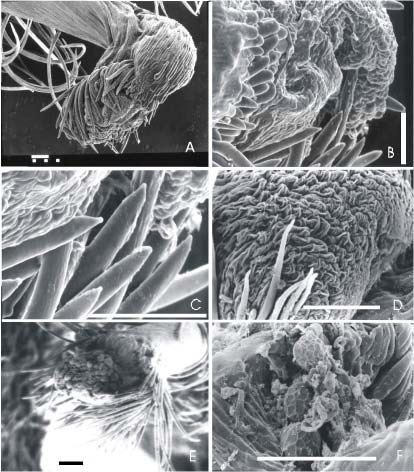
Fig. 3.- Estigmas (A, B y C: V. caracalla, Palacios y Bravo 1351, D: V. luteola Palacios 1300, E y F: V. adenantha, Drewes 507) A: rotación del ápice del estilo; B: superficie del estigma previa a la antesis; C: tricomas del anillo peristigmático; D: superficie del estigma previa a la antesis; E: estilo-estigma; F: estigma con granos de polen, restos de membrana y exudado estigmático. La escala vale 100 μm para A, B, C, D, E y 20 μm para F.
El cepillo estilar ocupa un área triangular en la porción distal interna que acompaña la rotación del estilo sobre su eje. Los tricomas se disponen en la región basal en forma de V ubicándose los más largos en la región media y apical, una zona desnuda del estilo lo separa del anillo peristigmático; salvo en V. lasiocarpa, V. candida y V. peduncularis var. clitoroides que se continúa con los tricomas del anillo peristigmático. En V. candida los tricomas son cilíndricos, rectos, largos, patentes, densamente dispuestos; la superficie es lisa en la base, pero a partir de 1/3 de la longitud del tricoma una mitad del cilindro presenta tanto estrías longitudinales cortas como largas, de relieve positivo; la otra mitad es lisa (Fig. 4 A y B). En V. caracalla los tricomas están laxamente dispuestos, rectos en su mayoría, se dirigen hacia el ápice del estilo, algunos de los tricomas más apicales del cepillo se curvan en el ápice. Son cilíndricos, en una mitad del cilindro la superficie del tricoma presenta estrías longitudinales cortas y largas, de relieve positivo; la otra mitad es lisa (Fig. 4 C y D). En V. peduncularis var. clitoroides se encuentran dispuestos frecuentemente pares de tricomas apareados en su longitud desde 1/3 o menos de la base y hasta casi el ápice que se presenta levemente curvo. Los tricomas son cilíndricos y la superficie es lisa (Fig. 4 E y F). En V. adenantha el cepillo estilar posee gran densidad de tricomas largos que se orientan hacia el ápice del estilo y se curvan laxamente en su mitad superior en forma de hoz, los tricomas distales del cepillo estilar quedan del lado opuesto al anillo peristigmático debido a la rotación del eje. Los tricomas son cilíndricos canaliculados, con un surco longitudinal que se desarrolla en toda la extensión del tricoma. La superficie del tricoma presenta a un lado del surco estrías cortas longitudinales y al otro lado diminutas deposiciones irregulares; la superficie opuesta al surco es lisa. (Fig. 4 G y H). En V. luteola los tricomas son largos y se dirigen desde la línea media del cepillo hacia ambos lados, envolviendo al estilo. Son canaliculados, con un surco longitudinal profundo interrumpido regularmente por unas estructuras lenticulares que sobresalen del mismo. La superficie es lisa a ambos lados del surco y en la mitad opuesta al mismo (Fig. 5 A y B). En V. longifolia los tricomas son sigmoideos, se encuentran dispersos y no presentan una orientación homogénea. Son canaliculados, presentan sobre el surco ancho concavidades bien marcadas, limitadas por crestas dispuestas regularmente en toda la longitud. La superficie es lisa a ambos lados del surco, y en la superficie opuesta a éste (Fig. 5 C, D, E y F). En V. lasiocarpa los tricomas de la mitad basal del cepillo son sigmoideos y se dirigen hacia el ápice del estilo; los de la mitad apical son rectos y se dirigen hacia la base del estilo. Son canaliculados, con un surco no muy marcado, que presenta pliegues transversales los cuales delimitan superficies más planas y anchas, levemente cóncavas. La superficie es lisa a ambos lados del surco, y en la mitad opuesta al mismo (Fig. 5 G y H)
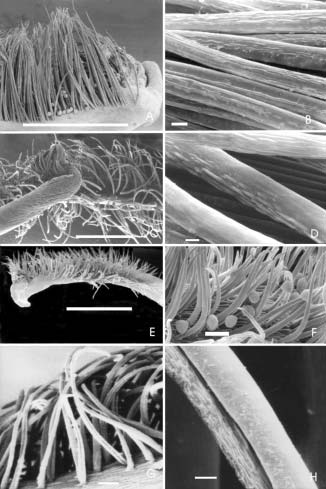
Fig. 4.- Cepillo estilar (A y B: V. candida, Palacios 880; C y D: V. caracalla, Palacios y Bravo 1351; E y F: V. peduncularis var. clitoroides, Palacios y Bravo 1294; G y H: V. adenantha, Drewes 507). A: vista del cepillo estilar; B: detalle de los tricomas del cepillo estilar; C: vista del cepillo estilar; D: detalle de los tricomas del cepillo estilar; E: vista del cepillo estilar; F: detalle de los tricomas del cepillo estilar; G y H: detalle de los tricomas del cepillo estilar. La escala vale 5 μm para H; 10 μm para B y D; 50 μm para G; 100 μm para F y 1000 μm para A, C y E.
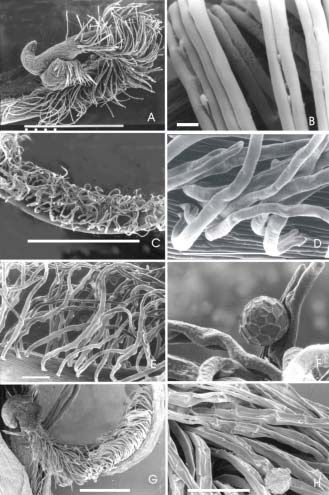
Fig. 5.- Cepillo estilar (A y B: V. luteola Palacios 1300: C, D, E y F: V longifolia, Palacios y Bravo 1278, G y H: V. lasiocarpa, Palacios y Bravo 1285) A: estilo y estigma; B: detalle de tricoma del cepillo estilar; C: vista del cepillo estilar; D, E: vista anterior y lateral de tricomas; F: detalle del tricoma con grano de polen; G: vista del cepillo estilar; H: detalle de los tricomas del cepillo estilar. La escala vale 10 μm para B y F; 100 μm para D, E y H y 1000 μm para A, C y G.
Los esquemas comparativos del ápice del estilo se presentan en la Fig. 6.
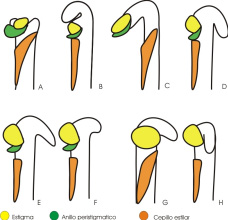
Fig. 6.- Esquemas de los estilos-estigmas (A, B, C, D, E y F: vista lateral; H: vista frontal). A: V.adenantha; B: V: candida; C: V. caracalla; D: V. lasiocarpa; E: V. luteola; F: V. peduncularis var. clitoroides; G y H: V. longifolia.
Discusión
Las especies estudiadas poseen un patrón similar de desarrollo del gineceo. La ubicación extrorsa del cepillo estilar (en los primeros estadios) cambia a introrsa (a la antesis), debido a la secuencia de curvatura y de rotación del estilo sobre su eje, como fue observado en V. longifolia (Gamba & Drewes, 2001).
En las flores en antesis varían: las relaciones de longitud estilo/ovario, los ángulos de curvatura del estilo, la ubicación del estigma, la presencia y la forma del rostro, las características del anillo peristigmático, las características del cepillo estilar y la esculturación de los tricomas del cepillo estilar. Estas variaciones son particulares para cada una de las especies, así como la combinación de las mismas, y definen distintos tipos de estilos/ estigmas (cfr. Fig. 6). Estas observaciones en las especies de Vigna, las diferencian de otros grupos de leguminosas, como Medicago (Trifoliae), donde la morfología de la región estigmática parece proveer caracteres muy conservativos, con especies más o menos relacionadas siendo uniformes morfológicamente y presentándose diferencias más significativas por encima del nivel de género (Small et al., 1983).
Los estudios de la biología floral realizados en varias especies de Vigna, las ubican como xenógamas facultativas, comprobándose la existencia de autofecundación (Hoc et al., 1993; Agulló et al., 1993; Etcheverry et al., 2008). En estas especies la asimetría floral y la curvatura del estilo favorecen el contacto con el visitante floral; el cepillo estilar interviene en la presentación secundaria del polen, mientras que la ubicación terminal o subterminal del estigma y la presencia de tricomas largos en el anillo peristigmático, impedirían la llegada del polen propio al estigma (Lavin & Delgado, 1990; Lloyd & Schoen, 1992).
Con respecto a las revoluciones de la quilla, Faegri & Van der Pijl (1979) especulan que más revoluciones y complejidad en las flores causarían la pérdida de funcionalidad floral y una tendencia hacia la autogamia. Las revoluciones de la quilla también son observadas en el estilo; como una consecuencia el estilo de V. caracalla es muy largo (85 mm) (Etcheverry et al., 2008), excediendo los valores máximos relatados por Shivanna & Owens (1989) en su revisión de Papilionoideae (30-60 mm máximos en Arachis). Mulcahy (1979) notó que el estilo de angiospermas proporciona un área selectiva; ya que mutaciones deletéreas podrían retardar el crecimiento del tubo polínico, o sea, cuanta mayor distancia a recorrer menor posibilidad de competir por los óvulos, y así ser eliminado del fondo genético.
La posición del estigma queda definida por la curvatura del estilo que acompaña la quilla y su ubicación es fundamental para la zona de contacto con el polinizador. En V. minima la quilla cumple un giro completo, por lo cual el mecanismo de polinización es esternotribo, los insectos visitantes se posan sobre el ala izquierda, al igual que en V. longifolia, sin embargo, en esta última especie, la quilla no cumple un giro completo sino de 270º, siendo el mecanismo de polinización nototribo (Hoc et al., 1993). La morfología floral de V. longifolia es una imagen especular de V. vexillata, ambas presentan mecanismo de polinización nototribo (Hoc et al., 1993). En V. luteola el estilo apenas curvo, queda encerrado en la quilla navicular, el mecanismo de polinización es esternotribo explosivo leve, dado que por acción del polinizador se produce el descenso de la quilla y las alas, y emergen bruscamente el estilo (que presenta en forma secundaria el polen en el cepillo estilar) y los estambres, el polen es expuesto a modo de una nube que cubre principalmente el abdomen del visitante (Agulló et al., 1993).
Entre las especies de Vigna que se conoce la biología floral sólo V. minima y V. luteola presentan un mecanismo de polinización esternotribo, mientras que V. vexillata, V. longifolia, V. adenantha y V. caracalla tienen un mecanismo de polinización nototribo. Considerando que el sitio de colocación de polen, más encontrado en Papilionoideae cigomorfas es el esternotribo; la variación a nototribo está presente en las especies con morfología floral más compleja (Faegri & Van der Pijl, 1979) y muestra una tendencia a economizar néctar y polen por medio de mecanismos de polinización que sólo pueden ser activados por los visitantes adecuados (Arroyo, 1981). Probablemente este relacionado con un mecanismo para reducir al mínimo la deposición de polen heteroespecífico, ya que varias especies de papilionoideas cigomorfas viven en simpatría y compiten por los mismos polinizadores (Etcheverry et al., 2008).
Lloyd & Schoen (1992) concluyeron que las familias caracterizadas por mecanismos de polinización especializados, como Papilionoideae, han sufrido la selección natural que promueve la polinización cruzada, considerando dos rasgos estructurales del gineceo que son relacionados con el sistema reproductivo: 'la membrana' estigmática y el anillo de tricomas peristigmático.
Los estigmas de las especies estudiadas corresponden al tipo húmedo según Heslop-Harrison & Shivanna (1977) y Heslop-Harrison (1981), presentan papilas en los primeros estadios, las que necrosan a medida que producen un exudado que queda retenido debajo de una membrana cuticular hasta su rotura, tal como lo señalado por Castro & Agulló (1998) para V. adenantha. La membrana estigmática previene la autopolinización y requiere del paso de un polinizador que cause su ruptura para la polinización eficaz (Arroyo, 1981), aunque ha sido señalada su rotura espontánea en las últimas etapas de la antesis en Clianthus puniceus (Heenan, 1998). Esta membrana, esta presente en siete tribus de Faboideae, entre ellos Phaseoleae (Shivanna & Owens, 1989).
La presencia de pelos peristigmáticos puede maximizar la fecundación cruzada previniendo la deposición del polen propio sobre el estigma durante las primeras etapas de antesis. Observaciones similares han sido relatadas para otras legumbres Papilionoideae, como Macroptilium panduratum (Drewes, 2005), Phaseolus coccineus (Ibrahim & Coyne, 1975) y Crotalaria micans (Etcheverry et al., 2003).
Siguiendo el criterio de Lavin & Delgado (1990), se puede considerar que, V. adenantha, V. luteola, V. peduncularis var. clitoroides, V. caracalla y V. candida, poseen estigma penicilado por la longitud de los tricomas del anillo peristigmático y su ubicación sobrepasando las anteras.
El estigma de V. longifolia no posee anillo peristigmático y el de V. lasiocarpa es incompleto con pelos cortos lo que no impediría la llegada del polen propio, así mismo la curvatura del estilo lo acerca a las anteras.
La distribución del indumento en el estilo coincide con el cepillo polínico descripto en Macroptilium lathyroides por Lavin & Delgado (1990), quienes lo definen como «una densa agregación de tricomas erectos iniciados directamente en el extremo distal del estilo». En este trabajo se utiliza el término cepillo estilar para denominar a esa agregación de tricomas en la porción distal del estilo (diferenciado de los tricomas que se originan rodeando al estigma y de aquellos que son una continuación de la pubescencia del ovario), debido a que este cepillo puede no estar directamente relacionado con la presentación secundaria del polen en la polinización o bien ésta puede presentarse en cualquier otra estructura de la flor. Por ejemplo, en Macroptilium fraternum el cepillo estilar no interviene en la presentación secundaria del polen y constituye una condición relictual en flores cleistógamas y autógamas cuya autopolinización ocurre en el capullo previo a la antesis (Drewes & Hoc, 2000; Hoc et al., 2003).
En todas las especies estudiadas está presente el cepillo estilar en una ubicación más o menos similar, los tricomas están dispuestos de forma tal que aumentarían la eficacia en la presentación secundaria del polen, siendo la esculturación de los mismos una adaptación para la mejor adherencia de los granos. Este cepillo contiene una gran masa de polen que es dirigido para su expulsión por la apertura terminal de la quilla, por los movimientos que produce el insecto polinizador, y que va a ser descargado completamente sólo después de varias visitas (Etcheverry et al., 2008).
Este trabajo es la primera mención de la variación que presentan los tricomas del cepillo estilar, encontrándose características que tampoco habían sido señaladas en tricomas del indumento de otros órganos como: tricomas canaliculados, tricomas canaliculados con lentes y tricomas con concavidades.
Los tricomas del cepillo estilar de V. longifolia pueden considerarse como la máxima expresión de la adaptación a la presentación secundaria del polen. Los pliegues curvos que presentan estos tricomas poseen las concavidades de un diámetro algo mayor al de los granos de polen por lo que estos encajan perfectamente (cfr. Fig. 5 F).
En V. peduncularis var. clitoroides los tricomas del cepillo estilar son cilíndricos, no presentan surco y la superficie es homogéneamente lisa; Sin embargo, se encuentran frecuentemente pares de tricomas apareados en su longitud desde 1/3 o menos de la base y hasta casi el ápice. De manera que, funcionalmente, un par de tricomas presenta una superficie similar a la de un tricoma canaliculado.
A partir de la descripción de los nuevos caracteres señalados en este trabajo, se podrían generar hipótesis de homología de los estados que presentan las especies, esperando que sean útiles para comprender mejor la sistemática y las relaciones de parentesco tan problemáticas de resolver en el género Vigna.
Agradecimientos
Al Dr. Pablo Picca por su atención en microscopía electrónica, a las sugerencias de los revisores, a la Universidad de Buenos Aires por los subsidios otorgados (UBACYT X430), con parte de los cuales fue financiada la presente investigación.
Referencias bibliográficas
1. Agulló, M. A., M. M. Brizuela, P. S. Hoc, V. S. Di Atilio, R. A. Palacios, J. Genise & P. Hazeldine. 1993. Relación unidad de polinización- visitantes florales en Vigna luteola (Leguminosae, Phaseoleae). Bol. Soc. Argent. Bot. 29 (3-4): 131-138. [ Links ]
2. Arroyo, M. T. K. 1981. Breeding systems and pollination biology in Leguminosae, en: R. M. Polhill & P. H. Raven (eds.). Advances in legume systematic, pp. 723-769. Part 2. Royal Botanic Gardens, Kew, London. [ Links ]
3. Baudet, J. C. & R. Maréchal. 1976. Signification taxonomique de la presence de poils uncinulés chez certains genres de Phaseoleae et d´Hedysareae (Papilionaceae). Bull. Jard. Bot. Natl. Belg. 46: 419-426. [ Links ]
4. Bruneau A., J. J. Doyle & J. D. Palmer 1990. A chloroplast DNA inversion as a subtribal character in the tribe Phaseoleae (Leguminosae). Syst. Bot. 15: 378-386. [ Links ]
5. Castro, M. A. & M. A. Agulló. 1998. Anatomy of the stigma of Vigna adenantha (G. F. Meyer) Marechal, Mascherpa & Stainer (Leguminosae, Papilionoideae). Biocell 22: 9-18. [ Links ]
6. Drewes, S. I. & P. S. Hoc. 2000. Morfología y desarrollo de flores cleistógamas en Macroptilium fraternum (Fabaceae). Kurtziana 28: 229-238. [ Links ]
7. Drewes, S. I. 2005. Morfología de estilos y estigmas en Macroptilium (Fabaceae). Kurtziana 31: 29-38. [ Links ]
8. Etcheverry A.V., J. J. Protomastro & C.Westerkamp. 2003. Delayed autonomous self-pollination in the colonizer Crotalaria micans (Fabaceae: Papilionoideae): structural and functional aspects. Plant Syst. Evol. 239: 15-28. [ Links ]
9. Etcheverry A. V., M. M. Alemán & T. Figueroa Fleming. 2008. Flower Morphology, Pollination Biology and Mating System of the Complex Flower of Vigna caracalla (Fabaceae: Papilionoideae). Ann. Bot. 102: 305-316. [ Links ]
10. Faegri K. & L. Van der Piel. 1979. The principles of pollination ecology (ed. 3). UK Pergamon Press, Oxford. [ Links ]
11. Gamba C. & S. I. Drewes. 2001. Desarrollo morfológico del estilo/estigma en Vigna longifolia (Fabaceae). Bol. Soc. Argent. Bot. 36: 16 [ Links ]
12. Gopinathan M. Ch. & R. Babu.. 1987. Breeding systems and pollination in Vigna minima (Leguminosae, Papilionoideae). Plant Syst. Evol. 156: 117-126. [ Links ]
13. Hedstrom I. & M. Thulin. 1986. Pollination by a hugging mechanism in Vigna vexillata (Leguminosae, Papilionoideae). Plant Syst. Evol. 154: 275-283. [ Links ]
14. Heenan P. B. 1998. The pollination system and stigmatic cuticle of Clianthus puniceus (Fabaceae). New Zealand J. Bot. 36: 311-314. [ Links ]
15. Heslop Harrison Y. & K. R. Shivanna. 1977. The receptive surface of the Angiosperm stigma. Ann. Bot. 41: 1233-1258. [ Links ]
16. Heslop Harrison Y. 1981. Stigma characteristics and angiosperm taxonomy. Nord. J. Bot. 1: 401-420. [ Links ]
17. Hoc P. 1999. Vigna, en Zuloaga F. O. & O. Morrone (eds.).Catálogo de las Plantas Vasculares de la República Argentina II, pp. 740-741. Missouri Botanical Garden Press. [ Links ]
18. Hoc P., V. Di Stilio, M. Agulló, M. Brizuela, R. Palacios, J. Genise & P. Hazeldine. 1993. Biología floral de Vigna longifolia (Leguminosae, Phaseoleae). Darwiniana 32: 27-39 [ Links ]
19. Hoc P., S. Drewes & M. T. Amela. 2003. Biología floral, sistema y éxito reproductivo de Macroptilium fraternum (Fabaceae). Rev. Biol. Trop. 51: 369-380 [ Links ]
20. Holmgren P. K., N. H. Holmgren & C. Batnett. 1990. Index Herbariorum. Part I: The Herbaria of the World. Ed. 8 Reg. Veg. 120: 1-693. New York Botanical Garden, New York. [ Links ]
21. Ibrahim A. & D. Coyne. 1975. Genetics of stigma shape, cotyledon position and flower color in reciprocal crosses between Phaseolus vulgaris and Phaseolus coccineus. J. Amer. Soc. Hort. Sci. 100: 622-626. [ Links ]
22. Izaguirre P. & R. R. Beyhaut. 1997. Las Leguminosas en Uruguay. Montevideo. [ Links ]
23. Lackey J. A. 1981. Phaseoleae, en M. Polhill & P. H. Raven (eds.) Advances in Legume Systematics, part 1, pp. 301-327. Royal Botanic Garden. Kew. [ Links ]
24. Lavin M. & A. Delgado Salinas. 1990. Pollen brush of Papilionoideae (Leguminosae): Morphological variation and systematic utility. Am. J. Bot. 77: 1294-1312. [ Links ]
25. Lloyd D. G. & D. J. Schoen. 1992. Self-and cross-fertilization in plants. I. Functional dimensions. Int. J. Pl. Sci. 153: 358-369. [ Links ]
26. Marèchal R., J. M. Mascherpa & F. Stainer. 1978 a. Etude taxonomique d'un groupe complexe d'espèces des genres Phaseolus et Vigna (Papilionaceae) sur la base de donneés morphológiques et polliniques, traiteés par l'analyse informatique. Boissiera 28: 1-273. [ Links ]
27. Mulcahy D. L. 1979. The rise of angiosperms: a genecological factor. Science 206: 20-23 [ Links ]
28. Small E. & B. Brookes. 1983. The systematic value of stigma morphology in the legume tribe Trifolieae with particular reference to Medicago. Canad. J. Bot. 61: 2388-2404. [ Links ]
29. Shivanna K. R. & S. J. Owens. 1989. Pollen pistil interactions (Papilionoideae). In C. H. Stirton & J. L. Zarucchi (eds.). Advances in legume biology. Monogr. Syst. Bot. Missouri Bot. Gard. 29: 157-182, Missouri Botanical Garden Press. [ Links ]
Original recibido el 19 de julio de 2010;
primera decisión: 26 de septiembre de 2010;
aceptado el 19 de noviembre de 2010.
Editor responsable: Julieta Nattero.

