










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkKurtziana
versión On-line ISSN 1852-5962
Kurtziana vol.37 no.1 Córdoba ene./jun. 2012
ARTÍCULOS ORIGINALES
New species and reports of Inocybe (Agaricales) from Guyana
P. Brandon Matheny 1, M. Catherine Aime 2, Matthew E. Smith 3, * & Terry W. Henkel 4
1 Department of Ecology and Evolutionary Biology, University of Tennessee, Knoxville, TN 37996-1610, U.S.A.
2 Department of Plant Pathology and Crop Physiology, Louisiana State University Agricultural Center, Baton Rouge, LA 70803, U.S.A.
3 Biology Department, Duke University, Durham, NC 27705, U.S.A.
4 Department of Biological Sciences, Humboldt State University, Arcata, CA 95521, U.S.A.
* Current address: Department of Plant Pathology, University of Florida, Gainesville, FL 32611, U.S.A.
Summary
Since our last report in 2003, we document the discovery of an additional seven species of Inocybe from Dicymbe (Fabaceae) forests of Guyana in northeastern South America. Of these, five are described as new: I. enigmatica, I. lepidotella, I. magnifolia, I. marginata, and I. rhodella. Two additional species described originally from Brazil and Venezuela, I. amazoniensis and I. lasseri, respectively, are reported as new records for Guyana. This raises the number of Inocybe species reported from Guyana to eleven. Molecular identifications of ectomycorrhizal root tips of species of Dicymbe and Aldina (Fabaceae) match five of the known Inocybe species from Guyana. An additional eight Inocybe ITS sequences distinct at the species level were detected from ectomycorrhizal root tips of Fabaceae in Guyana but at present match no known morphological species represented in regional or global sequence databases. A revised key to species of Inocybe from Guyana is presented together with illustrations of new taxa. Data from examination of types of I. amazoniensis and I. matrisdei are presented, as well as supplementary reports of I. epidendron and I. pulchella, both originally described from Guyana.
Key words: Ectomycorrhizal fungi; Fabaceae; Guiana Shield; Inocybaceae; Neotropics; New species; South America; Systematics.
Resumen
Nuevas especies y registros de Inocybe (Agaricales) de Guyana
Desde nuestro último reporte en 2003, documentamos el descubrimiento de siete especies adicionales de Inocybe en bosques de Dicymbe (Fabaceae) en Guyana, en el norte de Sudamérica, de las cuales cinco son especies nuevas: I. enigmatica, I. lepidotella, I. magnifolia, I. marginata e I. rhodella, dos son especies descritas originalmente de Brasil y Venezuela: I. amazoniensis e I. lasseri, respectivamente y son nuevos registros para la región. Esto eleva el número de especies de Inocybe registradas para Guyana a once. Identificaciones moleculares de puntas de raíces ectomicorrizadas de Dicymbe y Aldina (Fabaceae) coinciden con cinco de las especies de Inocybe conocidas de Guyana. Adicionalmente se detectaron ocho secuencias de ITS distintas a nivel de especie que se obtuvieron a partir de puntas de raíces ectomicorrizadas de Fabaceae, estas secuencias no coinciden con ninguna especie morfológica representada en las bases de datos, tanto regionales como mundiales. Se presenta una clave taxonómica de las especies de Inocybe de Guyana, con ilustraciones de los nuevos taxones. Así como datos de los tipos de I. amazoniensis e I. matrisdei y material suplementario de I. epidendron e I. pulchela, estas últimas originalmente descritas de Guyana.
Palabras clave: Hongos ectomicorrícicos; Fabaceae; Escudo de Guyana; Inocybaceae; Neotrópicos; Especie nueva; Sudamérica; Sistemática.
Introduction
The family Inocybaceae Jülich, containing the genera Inocybe (Fr.) Fr., Auritella Matheny & Bougher, and Tubariomyces Esteve-Rav. & Matheny, is a highly diverse monophyletic group of ectomycorrhizal (ECM) fungi that comprises between 500 and 700 species worldwide (Matheny et al. 2009, Alvarado et al. 2010). Between 70% and 80% of species in the family have been described from the north temperate zone in association primarily with ECM plant families Pinaceae, Fagaceae, and Salicaceae. However, most studies on fungal ecology and biodiversity have focused on taxa outside the neotropics, a region where ECM fungal taxa deserve more attention (Alexander & Selosse 2009, Smith et al. 2011).
While some recent progress has been made on documentation of Inocybe from South America (Cortez & Coelho 2005, Meijer 2006, Wartchow et al. 2008), reports of Inocybe from the neotropics are generally infrequent. An earlier study by Matheny et al. (2003) estimated that 25 sufficiently documented species of Inocybe were known from the neotropics, including lowland rainforests and montane oak forests. However, molecular sampling has indicated that the neotropical Inocybe assemblage is polyphyletic (at least four independent lineages have been identified, three from Guyana) and contains isolated evolutionary branches that appear to have originated during the Miocene and Paleogene (Matheny et al. 2009). Unfortunately, the ecology of neotropical Inocybe is rather vague due to uncertainty of plant associate data. Moreover, the trophic status for several species is unclear due to fruiting patterns on elevated trunks of Dicymbe, a caesalpinoid genus of Fabaceae (Henkel et al. 2011), and/or on woody debris. It is not known if such a habit is an adaptation to extremely wet environments as in some ECM fungi (Henkel et al. 2000), or whether the habit is indicative of saprotrophy or parasitism of live trees.
Here we document seven additional species of Inocybe from Guyana, five of which are described as new. We also confirm the trophic status for five species of Inocybe, three of which are lignicolous or tree trunk inhabiting, as ECM associates of Dicymbe and/or Aldina, two genera of Fabaceae sympatric in Guyana, in which the ECM condition evolved independently (Smith et al. 2011). The report of seven additional species here raises the number of Inocybe species reported from Guyana to eleven.
Materials and methods
Field locations
Collections were made during expeditions to the Upper Potaro River Basin of the west-central Pakaraima Mountains of Guyana during the May- July rainy seasons of 2001-2006 and 2010, and the December-January rainy seasons of 2003-2004. Specimens were collected within a ca. 5 km radius of a base camp located at N5°18'04.8", W59°54 '40.4" in forests dominated by ECM trees of Dicymbe corymbosa Spruce ex Benth. (Fabaceae, subfamily Caesalpinioideae) (Henkel et al. 2011).
Specimen-vouchers
Color notations used to describe basidiomata in the field followed Kornerup & Wanscher (1978), where, for example, 6D5 refers to plate 6, column D, row 5, or were approximated. Additional color notations were made using the Munsell Soil Color Charts (1954) and Ridgway (1912) in comparison with digital photographs of specimens in the field taken under natural lighting. Munsell designations were made according to plate, value, and chroma (e.g., 10YR 5/4). Ridgway colors are capitalized. Specimens were described following methods of Matheny et al. (2003) and were dried with silica beads in individual containers and later archived in airtight plastic bags.
Holotype collections are deposited at BRG, the Guyana National Herbarium in Georgetown, and isotypes (where possible) at the University of Tennessee (TENN). Herbarium abbreviations follow that of the online version of Thiers [continuously updated]. Latin descriptions are not provided for novel taxa since Latin validating diagnoses are no longer required according to rules of the International Code of Nomenclature for algae, fungi, and plants (Miller et al. 2011). Mycobank (MB) numbers are indicated for novel taxa.
Microscopy
Dried specimens were sectioned by hand and rehydrated in 5% KOH solution and examined under a Nikon Eclipse 80i compound light microscope. Typically, twenty basidiospores were measured from a single specimen and include projections of nodules. Basidiospore measurements were made with use of a calibrated micrometer or NIS elements D imaging software. The number of basidiospores measured is indicated using the notation (n=30/2) where 30 is the total number of basidiospores observed from two collections. Quotients (Q) of basidiospore length divided by width are included. Outlying measurements observed less than 5% of the time are placed in parentheses. Mean values are italicized, and standard deviations are provided. The number of nodules per basidiospore was estimated across all possible planes of view.
DNA extraction, PCR, and sequencing of basidiomata
We amplified the internal transcribed spacers (ITS) region and the first 900-1400 base pairs of the nuclear large subunit ribosomal RNA gene (LSU) region. DNA extraction, PCR, and sequencing protocols were previously published as follows: in the Hibbett laboratory at Clark University (Matheny et al. 2007), the Aime laboratory at Louisiana State University (Henkel et al. 2010), the Vilgalys laboratory at Duke University (Henkel et al. 2010), and the Matheny laboratory at the University of Tennessee (Judge et al. 2010). Twenty new sequences (JN642226-JN642244) from basidiomata have been submitted to GenBank.
ECM root sampling of Dicymbe corymbosa, D. altsonii, and Aldina insignis
Ectomycorrhizal (ECM) roots of Inocybe were sampled during two different studies of belowground ECM fungal communities in the Upper Potaro region. One study conducted in the long-term research plots of Henkel et al. (2011) used 454 pyrosequencing and a cloning-based approach to sequence ECM fungi from 80 pooled root samples of large, multistemmed Dicymbe corymbosa trees (Smith & Henkel, unpublished). The other study from a nearby site sampled roots from three leguminous ECM tree species in two genera (Dicymbe and Aldina) and employed Sanger sequencing of individual ECM roots (Smith et al. 2011). Molecular operational taxonomic units (MOTUs) of Inocybe species from ECM roots were calculated by grouping sequences with 97% similarity across the entire internal transcribed spacer region (ITS1-5.8S-ITS2) in Sequencher (Gene Codes Corporation, Ann Arbor, Michigan). Grouping ITS sequences at 97% similarity level has shown to correspond well with species for most taxonomic groups of ECM fungi (Smith et al. 2007). Six new sequences from ECM root tips (JN663842-JN663844, JN681195-JN681197) have been submitted to GenBank.
Results
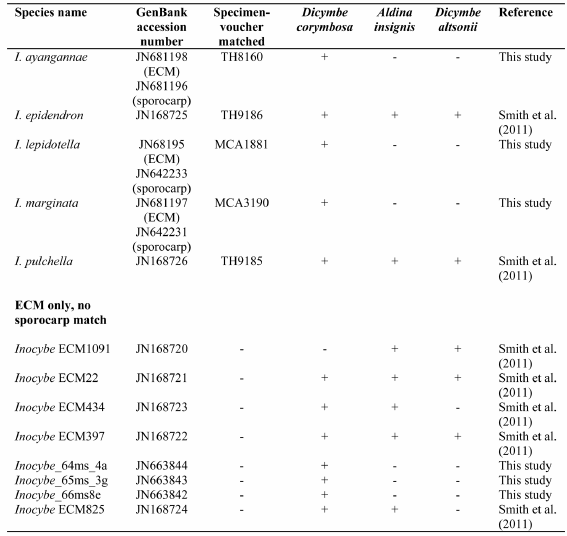
Table 1 provides a synopsis of Inocybe ITS sequences detected from ECM roots of Dicymbe corymbosa, D. altsonii, and Aldina insignis. Of the eleven species of Inocybaceae known from basidiomata in Guyana, we confirmed ECM root colonization by five species (I. epidendron Matheny, Aime & T.W. Henkel, I. pulchella Matheny, Aime & T.W. Henkel, I. lepidotella sp. nov., I. ayangannae Matheny, Aime & T.W. Henkel, and I. marginata sp. nov.). All five species are confirmed on ECM roots of D. corymbosa. Two species, I. epidendron and I. pulchella, are confirmed as ECM symbionts of D. altsonii and A. insignis, in addition to D. corymbosa (Smith et al. 2011). We were unable to obtain ITS data from basidiomata of I. lasseri so cannot confirm or deny the occurrence of this species on roots.
Table 1
Synopsis of Inocybe species confirmed from roots of Dicymbe corymbosa, Dicymbe altsonii, and Aldina insignis in Guyana.
Artificial taxonomic key to morphological species of Inocybe from Guyana
1. Basidiomata minute (pileus < 5 mm wide), dark yellowish brown or brown, disc with erect scales; spores smooth, pleurocystidia absent, on trunks of Dicymbe .................................................... Inocybe lepidotella
1'. Basidiomata larger than above (pileus > 10 mm wide), if smaller, then pleurocystidia present or spores nodulose or sterile, disc scaly or not, on ground in Dicymbe forest or on trunks of Dicymbe .................... 2
2. Stipe pruinose at least on the upper half and bearing metuloid caulocystidia on the lower part of the stipe, cortina absent .............................................................................................................................. 3
2'. Stipe fibrillose or squamulose, not pruinose, cortina present............................................................... 53. Basidiomata large, pileus 45-50 mm wide, stipe 50-65 x 9-14 mm at the apex with a bulb 14-25 mm wide; pink tones absent from lamellae and stipe surface ......................... Inocybe magnifolia
3'. Basidiomata smaller than above, pileus 10-25 mm wide, stipe 25-40 x 3-4 mm wide; pink tones present on lamellae and stipe ....................................................................................................... 44. Stipe with a distinct rimmed marginate bulb ........................................... Inocybe marginata
4'. Stipe base bulbous but not distinctly marginate ......................................... Inocybe rhodella5. Basidiomata fulvous or brownish yellow, pileus smooth, stipe scurfy-fibrillose, flesh in stipe blushing pink where exposed, pleurocystidia absent, sterile .......................................................................................................... Inocybe enigmatica
5'. Basidiomata brown, dark brown or with lilac or pink colors, pileus squamulose or fibrillose, stipe fibrillose-squamulose or fibrillose, stipe flesh not changing color where exposed, pleurocystidia present, basidiomata spore-producing .................................... 66. Pileus and stipe squamulose to squarrose, pileus 10-40 mm wide, terrestrial ...... 7
6'. Pileus smooth or appressed furfuraceous-scaly and stipe fibrillose, very small to small, 3-10 mm wide, terrestrial or on woody debris or trunks of Dicymbe ......... 87. Basidiomata brown to dark brown when fresh, pileus 20-40 mm wide, lamellae brown at maturity and lacking olivaceous tones ............ Inocybe ayangannae
7'. Basidiomata cream or pale yellowish with lilac-tipped squamules when fresh, pileus 5-15 mm wide, lamellae pale olivaceous in youth becoming isabelline or pale yellowish brown at maturity....................... Inocybe lilacinosquamosa8. Basidiospores > 10.0 mm long, subglobose to elliptic in outline with numerous coarse nodules ................................................. Inocybe lasseri
8'. Basidiospores < 10.0 mm long, polygonal in outline with moderate-sized nodules ................................................................................................... 99. Pileus with pinkish lavender tints when fresh, fading to brown or pale brown, basidiomata very small, pileus 2-6 mm wide, cheilocystidia and pleurocystidia thin-walled ............................. Inocybe pulchella
9'. Pileus lacking pinkish lavender tints, when fresh brown, light brown, or yellowish brown, basidiomata larger than above, pileus 5-15 mm wide, cheilocystidia and pleurocystidia thick-walled ................... 1010. Pileus squamulose with recurved scales to fibrillose-squamulose, at most with a slight obtuse umbo, occurring on trunks of Dicymbe or rarely on woody debris .......... Inocybe epidendron
10'. Pileus fibrillose and with a nipple-like umbonate disc with age, terrestrial ................................................ Inocybe amazoniensis
Taxonomy
Inocybe amazoniensis Singer & I.J. Araujo, Beih. Nova Hedwigia 77: 182. 1983. TYPE: Brazil. Amazonas, Estrada Manaus- Caracarai, B11019 (paratype, INPA!). Figs. 1, 7.

Figs. 1-6. Microscopic features of species of Inocybe newly reported from Guyana: a. basidiospores, b. pleurocystidia, c. caulocystidia and/or cauloparacystidia Fig. 1. I. amazoniensis (MCA3142). Fig. 2. I. lasseri (MCA1971). 3. I. lepidotella (MCA1881, holotype). 4. I. magnifolia (MCA2441, holotype). 5. I. marginata (MCA3190, holotype). 6. I. rhodella (MCA3917, holotype). Scale bars are equal to 10 µm.

Figs. 7-14. Basidiomata of Inocybe species from Guyana. Fig. 7. I. amazoniensis (MCA3142). Fig. 8. I. enigmata (MCA1868, holotype). Fig. 9. I. epidendron (MCA3160). Fig. 10. I. lasseri (MCA1971). Fig. 11. I. lepidotella (MCA1881, holotype). Fig. 12. I. marginata (MCA3190, holotype). Fig. 13. I. magnifolia (MCA1822, paratype). Fig. 14. I. rhodella (MCA3917, holotype). Scale bars are equal to 1 cm.
Pileus 5-10 mm diam, conical, expanding in age with nipple-like umbonate disc; margin decurved to straight, undulating, entire at first but later torn and rimose with striate appearance; surface dry, fibrillose, breaking up into appressed scurfy scales in age; light brown (near 6C4) or brown to yellowish brown (10YR 5/3-5/4) or near Tawny-Olive, pallid between spreading fibrils towards the margin; context very thin, odor salty or sweaty, taste none. Lamellae adnexed to sinuate, seceding in age, subdistant to moderately close, 24-30 L with several tiers of lamellulae; pale brown to yellowish brown (10YR 6/3-5/4), edges white and fimbriate but less obvious so with age. Stipe 20-23 x 1 mm, widest at the apex, tapered towards the base, flexuous, whitish fibrillose, pallid overall, nowhere pruinose, cortina probably evanescent though not directly observed.
Basidiospores 6.5-7.8-8.5 ± 0.57 x (5.0-) 5.5-6.0-6.5 ± 0.45 µm, Q = (1.08-) 1.14-1.29- 1.42 ± 0.09 (n=30/2), nodulose around mostly subelliptic outline with mostly 8-11 distinct or moderate-sized nodules, apiculus distinct, light yellowish brown or ochraceous-buff. Basidia 21-28 x 7-9 µm, 4-sterigmate, clavate, hyaline or filled with yellowish-ochre content when collapsed. Pleurocystidia 42-59 x 10-15 µm, fusiform to cylindric, necks not differentiated, apices obtuse and sparsely crystalliferous or most often bare, with a short basal pedicel; thick-walled, walls 1.0-2.5 µm thick apically, thin-walled towards the pedicel; hyaline to pale yellow. Cheilocystidia similar to pleurocystidia but shorter; paracystidia clavate, thin-walled, hyaline. Lamellar trama composed of parallel hyphae that are cylindric to inflated, up to 18 mm diam, almost hyaline or pale yellowish brown in mass. Pileipellis a cutis composed of cylindric to inflated hyphae up to 22 µm diam, thinwalled or at times slightly thickened at septa, hyphae incrusted, pale yellowish brown in mass. Stipitipellis lacking any caulocystidia, refractive hyphae not observed. Clamp connections present.
Material studied:
BRAZIL: Amazonas, Estrada Manaus-Caracarai, km 125, B11019 (paratype INPA), leg. Singer & Araujo, 21-May-1978, on leaf mold and among superficial roots in campinarana vegetation, associated with Fabaceae. GUYANA: Region 8 Potaro-Siparuni - Pakaraima Mountains, Upper Potaro River Basin, elevation 710 - 750m, scattered on ground across the river from base camp in Dicymbe forest, MCA3142 (BRG), leg. M.C. Aime, 30-Jun-2006 [JN642232-ITS, JN642237- LSU].
Observations: The macroscopic features are based on material from Guyana (MCA3142). Microscopic details are a composite of our examination of the paratype (B11019, INPA) from Brazil and MCA3142 from Guyana, which are in excellent agreement. Inocybe amazoniensis is characterized by a small hygrophanous pileus, a conspicuous papilla or nipple-like umbo on the disc of the pileus, the tapered stipe, nodulose spores, thick-walled hymenial cystidia, absence of caulocystidia, and a putative ECM association with Fabaceae, features shared by the Guyana material. LSU data of I. amazoniensis suggest the species may be most closely related to I. epidendron Matheny, Aime & T.W. Henkel (97% pair-wise similarity based on a BLASTn search). This is not surprising as a close relationship was suggested between the two by Matheny et al. (2003). Inocybe epidendron differs from I. amazoniensis by the scaly pileus and occurrence on trunks of Dicymbe or woody debris. Inocybe amazoniensis features a non-scaly pileus and appears terrestrial.
Phenology: May to June.
Habitat and Geographical Distribution: on ground in association with leguminous host genera Dicymbe in Guyana and Aldina in campinarana vegetation in Brazil (Singer et al. 1983).
Inocybe enigmatica Matheny & Aime, sp. nov. Fig. 8. MB 563638.
Pileus 10-15 mm diam, convex to broadly convex, at times with slightly depressed center, margin incurved, surface dry, minutely scurfy to smooth, brownish yellow or fulvous throughout or at the center and shading pale yellow (3A3) towards the margin, hygrophanous, fading to pale yellow (2A3); context -3 mm thick, odor unpleasant with a fishy-spermatic smell. Lamellae adnate to arcuate, subdistant, ca. 20- 24 L with few tiers of lamellulae, almost white, very narrow, ca. 1 mm deep. Stipe 20-30 x 4-8 mm, even, tapered, or somewhat enlarged at the base, scurfy-fibrillose to shaggy fibrillose, fulvous or colored like the pileus; context solid, white but blushing pink where exposed.
Basidiospores none. Basidia none or rarely observed, when present 35-47 x 7-8 µm and appearing as though 2-sterigmate;"hymenium" composed of hyaline, thin-walled, cylindric to slenderly subclavate cells, 31-48 x 5-7 µm, at times these connected by a clamp connection to subtending cells. Pleurocystidia none. Cheilocystidia not observed. Pileipellis a cutis and golden brown to brownish yellow in mass, hyphae cylindric, mostly 7-17 µm wide, thinwalled, lacking any obvious incrusting pigments; mounts exuding dull pinkish red pigment in 5% KOH, refractive hyphae not observed. Stipitipellis lacking caulocystidia, mounts not exuding pigments in KOH. Clamp connections present.
Material studied:
GUYANA: Region 8 Potaro-Siparuni - Pakaraima Mountains, Upper Potaro River Basin, elevation 710 - 750 m, on ground, deep in soil in Dicymbe plot 2, MCA1490 (BRG), leg MC Aime, 27-May-2001 [EU600897-LSU, EU600896-rpb1]; on ground in Dicymbe plot 2, MCA1868 (holotype BRG, isotype TENN066448), leg MC Aime, 1-Jun-2001 [JN642229-ITS, JN642236-LSU].
Observations: Phylogenetic data for I. enigmatica (as"I. sp. MCA1490") suggest a distant but well-supported position sister to a clade composed of I. ayangannae Matheny, Aime & T.W. Henkel and I. lilacinosquamosa Matheny, Aime & T.W Henkel, two additional species known only from Guyana (Matheny et al. 2009). Morphologically, I. enigmatica is unusual due to its sterile state, absence of pleurocystidia, reaction of pileus tissue to KOH, and pink staining of the stipe context. As of our 2003 publication (Matheny et al. 2003), we were aware of I. enigmatica but delayed documentation of the species due to anticipation of collecting additional mature specimens. Unfortunately, the species has not been observed since 2001. We describe I. enigmatica here as new because of its unique morphological features and strong support by molecular data as a unique and autonomous species (Matheny et al. 2009).
A third sterile collection (MCA2353, BRG; TENN066448) was observed 9-Jul-2003 near the base camp under Dicymbe but differs by the presence of numerous thick-walled, fusiform, hymenial cystidia, the stipe flesh not changing color where exposed, and the lack of a color reaction to pileus tissue mounted in KOH. Unfortunately, the material is in poor condition ensconced entirely by a hyphomycete.
Phenology: May to June.
Habitat and Geographical Distribution: on ground under Dicymbe corymbosa, known only from Guyana.
Inocybe epidendron Matheny, Aime, & T.W. Henkel, Mycol. Res. 107: 496. 2003. TYPE: Guyana: Region 8 Potaro-Siparuni, Pakaraima Mountains, MCA1473 (holotype, BRG). Fig. 9.
Material studied:
GUYANA: Potaro-Siparuni, Pakaraima Mountains, Upper Potaro River Basin, elevation 710-750 m, Benny's Ridge, on trunk of Dicymbe corymbosa, MCA3160 (BRG,), leg. MC Aime, 2-Jul-2006 [JN642238-LSU].
Observations: In addition to material cited in the protologue, we now add MCA3160. LSU sequence data of this species are the same as EU569840 (MCA1880). Inocybe epidendron is probably most closely related to I. amazoniensis based on morphology and LSU sequence similarity but differs in the field from the latter by the squamulose pileus. Smith et al. (2011) confirmed I. epidendron (and I. pulchella, see below) from ECM root samples of Dicymbe altsonii Sandwith and Aldina insignis (Table 1). Thus, these fungi form ECM associations despite their occurrence on the trunks of their hosts.
Phenology: May to July.
Habitat and Geographical Distribution: on root mat or trunks of Dicymbe corymbosa, also a confirmed root associate of Dicymbe altsonii and Aldina insignis, known only from Guyana.
Inocybe lasseri Dennis, Bull. Soc. mycol. Fr. 69: 197. 1953. TYPE: Venezuela. Figs. 2, 10.
Pileus 13 mm diam, plane with a decurved margin, umbo absent, surface translucentstriate to disc, glabrous with a scurfy disc, dull in appearance; brown (5D7) or Snuff Brown, pale brown between striae; context very thin. Lamellae adnexed, distant, ca. 20 L with few tiers of lamellulae, brown, edges pallid and fimbriate, narrow. Stipe 46 x 1.5 mm, even, smooth in appearance at the apex but with few scurfy fibrils below, cortina not directly observed; pallid with lavender (12B2) tones especially on the upper part; context solid, pallid with a lilac tinge.
Basidiospores 10.5-10.9-12.0 ± 0.69 x (9.0) 9.5-9.7-10.0 ± 0.30 µm, Q = 1.05-1.12-1.20 ± 0.06 (n=20/1), coarsely nodulose about a subglobose to elliptic outline, often with at least one bifid nodule, mostly with 14-16 nodules, some spores with multiple bifid nodules. Basidia 35-41 x 10-11 µm, 4-sterigmate, clavate, hyaline but lutescent and collapsing in age. Pleurocystidia 67-72 x 12-14 µm, mostly fusiform with obtuse apices and tapered basal pedicel, thick-walled, walls 2.0-3.0 mm thick and yellow. Cheilocystidia 41-84 (-100) x 10- 11 µm, hair-like, versiform, many slenderly clavate, some cylindric to fusiform, at times bifurcated near the apex, apices often swollen and wider than elsewhere, walls slightly thickwalled, often ochraceous. Pileipellis a cutis of cinnamon brown to tawny pigmented hyphae, these cylindrical and often 10-20 µm wide, slightly thick-walled, smooth in appearance. Stipitipellis with caulocystidioid cells similar to cheilocystidia though thin-walled and hyaline, these descending to just above the base, cylindric, unlike pleurocystidia in shape and size, metuloid caulocystidia observed only on the upper one-third of stipe surface. Clamp connections present.
Specimens studied:
GUYANA: Region 8 Potaro-Siparuni - Pakaraima Mountains, Upper Potaro River Basin, elevation 710 - 750 m, Dicymbe plot 2, MCA1971 (BRG), leg. M.C. Aime, 12- Jun-2002 [EU569856-LSU, EU569857-rpb2].
Observations: Inocybe lasseri is a poorly known species from the neotropics with a somewhat questionable taxonomic history. The type was recorded from Rio Chacaito, Venezuela by Dennis (1953). Dennis later (1970) included I. lasseri in a key with two other species of Inocybe in a fungus flora for Venezuela. Singer et al. (1983) considered I. lasseri (based on the description only) to be a tropical variant of the subtropical I. hyperythra Rick, described originally from São Leopoldo, in the state of Rio Grande do Sul, Brazil. However, I. hyperythra is more robust in size (pileus 10- 30 mm, stipe 30 x 4 mm) than I. lasseri and with cheilocystidia similar to pleurocystidia in morphology. Inocybe hyperythra is also reported with pink lamellae when young and featuring an almond taste. Unfortunately, we lack data on both taste and color of young lamellae of the Guyana material, but the cheilocystidia strike us as quite different in morphology than the pleurocystidia, and the size is not as robust as in I. hyperythra. In the protologue of I. lasseri, Dennis describes the context without any particular odor and mentions no pink coloration to the lamellae. However, the Guyanese material exhibits lavender tones on the stipe, whereas Singer reports a pink coloration to the stipe of I. hyperythra, in contrast to the protologue of I. lasseri, which describes the stipe as concolorous with the"Mars Brown" pileus. Pegler (1983), who reported I. lasseri from a degraded xerophytic forest on Guadeloupe, described the spores as 8-10.5 x 7-9 µm, somewhat smaller than the material from Guyana, spores of which match very well with those described in the protologue of I. lasseri.
In Guyana I. lasseri is readily recognized by the small brown basidiomata with a lavender tinge to the stipe, the versiform and hair-like cheilocystidia, and the large subglobose spores that are coarsely nodulose often with one or more saddle-shaped or bifid nodules, features consistent with the various interpretations of I. lasseri in a broad sense described above. Inocybe pulchella and I. epidendron may be confused with I. lasseri in the field, but the spores of these two species are much smaller, and both occur on trunks or root mats. Inocybe pulchella exhibits purplish to pinkish tones to the pileus, but its cystidia are consistently thinwalled in contrast to those of I. epidendron and I. lasseri. Matheny et al. (2009) indicated I. lasseri is the sister lineage to the clade containing I. pulchella and I. epidendron. This constitutes the first documentation of I. lasseri from Guyana.
Phenology: June (in Guyana).
Habitat and Geographic Distribution: on soil under Dicymbe, known from Venezuela, Guyana, and Guadeloupe (Lesser Antilles).
Inocybe lepidotella Matheny & Aime, sp. nov. Figs. 3, 11. MB 563639.
Pileus 3 mm diam, conical to deeply so, margin incurved to almost straight, dry, disc with erect squamules, sulcate towards the margin; dark brown to dark yellowish brown (5E8) on the disc, shading to strong brown or reddish brown or yellowish brown towards the margin; context very thin-fleshed, odor not observed. Lamellae adnexed, moderately close, narrow, brown. Stipe 20 mm x 0.5 mm, flexuous, even, surface with scattered pallid superficial fibrils at least at the apex, otherwise finely fibrillose, vinaceous brown to reddish brown or dark reddish brown overall.
Basidiospores 10.0-10.5-11.5 ± 0.46 x 7.0- 7.4-8.0 ± 0.34 µm, Q = 1.25-1.41-1.51 ± 0.07, (n=22/1), broadly amygdaliform to citriniform, at times mucronate, ovate in face view, yellowish brown, apiculus small and indistinct. Basidia 26-36 x 8-12 µm, clavate, 4-sterigmate, hyaline to yellowish. Pleurocystidia none. Cheilocystidia 15-35 x 8-10 µm, articulated or frequently septate, apical cell cylindric to saccate, hyaline, thin-walled, not frequent. Stipitipellis lacking caulocystidia. Pileipellis composed of cylindric hyphae, these mostly 7-15 µm wide, end cells not differentiated, thick-walled but with weak incrusting pigments, primarily with cytoplasmatic pigment. Clamp connections present.
Material examined:
GUYANA: Region 8 Potaro-Siparuni - Pakaraima Mountains, Upper Potaro River Basin, elevation 710 - 750 m, Dicymbe 1 plots, on trunk of Dicymbe in humus layer, MCA1881 (holotype BRG, isotype TENN066442) [JN642233-ITS, JN642235- LSU], leg M.C. Aime, 5-Jun-2001.
Observations: Inocybe lepidotella is an outstanding new species due to the very small size, scaly disc, conical pileus, vinaceous brown colored stipe, smooth spores, absence of pleurocystidia, and occurrence on the trunk of a Dicymbe. This represents the first report of a smooth-spored species of Inocybe from Guyana. Both ITS and LSU sequence data suggest I. lepidotella is a member of the Pseudosperma clade (Matheny et al. 2009), or section Rimosae s. str. (Larsson et al. 2009) based on blastn searches of GenBank. Unpublished phylogenetic analyses also affirm this placement.
Six other species in section Rimosae have been described from the neotropics. Five of these are considerably more robust than I. lepidotella. Pegler (1983) recorded two species from the Lesser Antilles: I. littoralis Pegler, a Coccoloba associate with an amber yellow pileus, and I. ingae Pegler, which differs easily by its pale ochraceous pileus and narrow phaseoliform spores. Singer et al. (1983) reported two species in section Rimosae from Costa Rica: I. nothodryina Singer, a Quercus associate that features a pale tan pileus with a white center, and I. fastigiata var. umbrinella (Bres.) R. Heim with a distinctive rimose pileus. Inocybe tropicalis Guzmán, originally described from tropical rainforest in Mexico, differs as well due to the brownish orange to pale brown and rimose pileus and oblong-ellipsoid spores (Guzmán 1982). Singer (1953) reported I. violaceolamellata Rick, a small species (pileus -10 mm wide) recorded from a mossy trunk in São Leopoldo, Brazil. This species differs from I. lepidotella by the violaceous colored lamellae and smaller phaseoliform spores reported by Singer (1953) to be about 9.5 x 4.8 µm. Rick (1961) applied the names of two European species, I. rimosa (Bull.: Fr.) P. Kumm. and I. fastigiata (Schaeff.) Quél, to two robust species from Brazil, but these names are likely misapplied assuming the materials were collected in habitats with native vegetation.
Phenology: June.
Habitat and Geographical Distribution: in humus layer on the trunk of a live Dicymbe (Fabaceae) tree, known only from Guyana.
Inocybe magnifolia Matheny, Aime, & T.W. Henkel, sp. nov. Figs. 4, 13. MB 563640.
Pileus 45-52 mm diam, convex to plane with a large low subacute umbo, margin decurved, surface dry, fibrillose, splitting and peeling at times, radially rimose to rimulose towards the margin, elsewhere with smooth appearance; buff to cinnamon buff or dull yellowish tan (4A3-4A5 to 4B3-4B5); context white to pallid, not changing color upon exposure, 3-6 mm thick underneath the disc, odor spermatic or weakly so, taste not remarkable. Lamellae adnexed, close with several tiers of lamellulae, undulating, pale yellowish white, brownish yellow, to yellowish brown with age, edges not distinctly pallid-fimbriate, 4-6 mm deep. Stipe 53-65 x 9-14 mm at the apex, base forming a tuberous bulb 14-25 mm wide, surface pruinose or indistinctly so, striatulate to appressed longitudinally fibrillose in appearance, at first white but at times umbrinous or Saccardo's Umber in dried state, bruising slowly pinkish when cut, especially at the base, but this feature may not be consistent; context white when fresh, spongy in the base.
Basidiospores 7.5-8.5-9.0 ± 0.40 x 5.0-6.4- 7.0 (-8.0) ± 0.63 µm, Q = (1.06-) 1.23-1.34-1.55 ± 0.12, (n=20/1), nodulose with 7-9 moderate sized nodules about a polygonal outline, yellowish brown. Basidia 29-35 x 9-12 µm, 4-sterigmate, clavate, hyaline. Pleurocystidia 68-84 x 15-22 µm, fusiform to fusiform-ventricose, apices obtuse, bare or sparsely crystalliferous, thick-walled, walls -3.0 thick, hyaline, often with a tapered basal pedicel. Cheilocystidia similar to pleurocystidia but mixed with colorless paracystidia. Stipitipellis with caulocystidia similar to hymenial cystidia, mixed with hyaline cauloparacystidia, abundant at the apex and present just above the stipe base. Pileipellis a cutis with a relatively thin suprapellis, this hyaline and composed of narrow, interwoven hyphae -8 µm wide; subpellis composed of a broader buff to very pale brown pigmented layer of cylindric hyphae, these 5-13 µm wide and weakly incrusted. Clamp connections present.
Material studied:
GUYANA: Region 8 Potaro-Siparuni - Pakaraima Mountains, Upper Potaro River Basin, elevation 710-750 m, in Dicymbe plot P3, MCA2441 (holotype BRG, isotype TENN066444), leg. M.C. Aime, 6-Jan- 2004 [EU600900-LSU, JN642244-LSU, JN642228-ITS]; same locality as above but near base camp on ground under Dicymbe, MCA1822 (BRG, TENN066443), leg. M.C. Aime, 7-Jun-2001 [JN642226-ITS]; 2 km southeast of Ayanganna airstrip near Tadang camp under Dicymbe altsonii and Aldina insignis, TH9132 (BRG; HSU) [JN642227- ITS].
Observations: The most outstanding features of I. magnifolia are its large size with respect to other species from Guyana and the pronounced tuberous bulb at the base of the stipe. Inocybe xerophytica Pegler bears a superficial resemblance to I. magnifolia-both share a pale colored pileus (buff to light buff) at least when fresh, a bulbous stipe base, nodulose spores, caulocystidia descending the length of the stipe, and a spermatic odor. However, I. xerophytica, the most common species of Inocybe on Guadeloupe in the Lesser Antilles (Pegler 1983), differs by its smaller size, smaller cystidia, plant host preference, and phylogenetic position (Matheny et al. 2009). Inocybe magnifolia (as I. sp. MCA2441) occurs in a clade together with I. marginata and I. rhodella (see below), I. mixtilis (Britzelm.) Sacc., and other nodulose-spored species with a caulocystidiate stipe from the northern and southern hemispheres (Matheny et al. 2009). Collection MCA1822 is sterile, but sequence data support its conspecifity with the type of I. magnifolia.
Phenology: January to June.
Habitat and Geographic Distribution: solitary on ground under Dicymbe or Aldina (Fabaceae), known only from Guyana.
Inocybe marginata Matheny, Aime, & T.W. Henkel, sp. nov. Figs. 5, 12. MB 563641.
Pileus 1.5-2.5 cm diam, plane, plicatestriate on the margin, edge scalloped, center glabrous and greasy and with a broad and low umbo, elsewhere dry, matt, appressed fibrillose, hygrophanous with fibrils becoming silvery-white after fading; when fresh color tan caramel to pale tan (5C6-5D5) on the umbo, pale pinkish taupe (6C3-6B3-6B2) around the umbo and towards the margin, most specimens with a pinkish tint towards the margin; context thin, pallid, not changing color where bruised, odor spermatic, taste not remarkable. Lamellae adnexed, subdistant, up to 20 L with several tiers of lamellulae, white at first becoming pallid (4B3) with a sordid salmon-pink tint (5A2- 5A3), becoming more or less clay color (5C4), edges pallid-fimbriate, subventricose, narrow to medium, up to 3 mm deep. Stipe 30-40 x 3-4 mm, even or tapered down to the abruptly marginate volvate-like bulb, this up to 6.5 mm wide, cortina absent, surface pruinose under a hand-lens with few scurfy fibrils on the lower part, generally with a glabrous appearance; white becoming pale clay, at times with pinkish tones and concolorous with the pileus; context hollow with white cottony-like fibrils in the interior.
Basidiospores (6.0-) 6.5-7.1-8.0 ± 0.52 x 5.0-5.8-6.5 ± 0.35 µm, Q = 1.08-1.22-1.36 ± 0.08, (n=30/2), exceptionally up to 10.0 x 9.0 µm probably from 1- or 2-sterigmate basidia, nodulose with mostly 8-11 moderate sized bluntly conic nodules about an angular to pentagonal outline in face view, yellowish brown, apiculus distinct. Basidia 20-32 x 7-10 µm, clavate or cylindrico-clavate, mostly 4-sterigmate, occasionally 1- or 2-sterigmate, hyaline to ochraceous. Pleurocystidia (38-) 60-75 x (14-) 19-33 µm, frequent, broadly to very broadly fusiform, occasionally lageniform, often with a short neck, generally with a short tapered pedicel or at times appearing sessile, thick-walled (rarely thin), walls 2.5- 5.0 µm thick, hyaline. Cheilocystidia similar to pleurocystidia, mixed with paracystidia. Stipitipellis with caulocystidia similar to pleurocystidia descending below the center of the stipe. Lamellar trama regular, compact, hyaline, composed of cylindric hyphae up to 13 µm diam, subhymenium pseudoparenchymatous. Pileipellis with a suprapellis composed of interwoven, smooth, hyaline, cylindric hyphae, mostly 4-7 µm wide, subtended by a parallel subcutis of mainly cylindric hyphae that are ochraceous brown in mass, hyphae up to 15 µm wide, incrusting pigments weak or not evident. Clamp connections present.
Material examined:
GUYANA: Region 8 Potaro-Siparuni - Pakaraima Mountains, Upper Potaro River Basin, elevation 710 - 750m, Dicymbe plot 1, MCA1882 (BRG), leg. M.C. Aime, 5-Jun-2001 [AY509115-LSU, AY509116- rpb1, AY509114- rpb2); in Dicymbe P1 plot, MCA2045 (BRG, TENN066451), leg. M.C. Aime, 17-Jun-2002 [JN642240-LSU]; on mineral soil in Dicymbe plot 1, MCA2166 (BRG), leg. M.C. Aime, 30-Jun-2002; Dicymbe plot 1, MCA2265 (BRG), leg. M.C. Aime, 30-Jun-2003; across Potaro River from base camp, MCA3190 (holotype BRG), leg. M.C. Aime, 4-Jul-2006 [JN642231- ITS, JN642239-LSU]; four km southeast of Potaro base camp near Dicymbe plot 2, TH8290 (BRG, HSU), leg. T.W. Henkel, 14-Jun-2001.
Observations: Inocybe marginata appears similar to a Bolivian species, I. matrisdei Singer (Singer 1962; Singer et al. 1983), but differs in having smaller spores and a non-squamulose pileus. The coloration of the two species appears similar with reddish and pinkish tones ascribed to the pileus and stipe by Singer, similar to I. marginata and I. rhodella (see below). Inocybe rhodella can be distinguished from I. marginata by a swollen bulb that is not distinctly marginate and differences in ribosomal RNA sequences. Molecular sampling of ECM root tips of Dicymbe corymbosa confirms the presence of I. marginata (Table 1).
Phenology: June to July.
Habitat and Geographical Distribution: typically occurring singly, terrestrial on mineral or clay soil under Dicymbe (Fabaceae), known only from Guyana
Inocybe pulchella Matheny, Aime, & T.W. Henkel, Mycol. Res. 107: 498. 2003. TYPE: Guyana: Potaro-Siparuni, Pakaraima Mountains, MCA1879 (holotype, BRG).
Pileus 5-7 mm diam, up to 4 mm high, surface shiny and glabrous when first collected but becoming matte and appressed fibrillose hours after collection; deep violet to lilac (9B3) with a rusty brown colored disc (5B5), margin appendiculate with lilac fibrils like combed hair; odor none. Lamellae adnexed, moderately close, lilac with rusty hues, edges minutely fimbriate, medium. Stipe 18 x 1 mm, even, ground color burgundy (9D6) to purple (11E6), inserted on woody substrate.
Material studied:
GUYANA: Region 8 Potaro-Siparuni -Pakaraima Mountains, Upper Potaro River Basin, elevation 710-750 m, on Dicymbe trunk in Dicymbe plot 1, MCA2264 (BRG), leg M.C. Aime, 30-Jun-2003; Dicymbe plot 1, on decorticated wood, MCA2270 (BRG) [JN642241-LSU], leg M.C. Aime, 30-Jun-2002; Dicymbe plot 2, MCA2341 (BRG) [JN642242-LSU], leg M.C. Aime, 8-Jul-2002.
Observations: Three additional collections of I. pulchella are reported here to supplement the original gross morphological description. Microscopic data from these collections are similar to those presented in the protologue (Matheny et al. 2003). nLSU sequences of two of these (MCA2270, MCA2341) are identical to MCA1488 (JN642243) and MCA1122 (EU600842), the latter published in Matheny et al. (2009), and cited in the original description. Inocybe pulchella is distinctive due to its very small size (pileus 5-7 mm wide), purplish to pinkish tones of the basidiomata that fade rapidly after collection, thin-walled hymenial cystidia, and occurrence on trunks of Dicymbe or decorticated wood. Despite its non-terricolous habit, I. pulchella is a confirmed ECM symbiont of Dicymbe and Aldina (Smith et al. 2011; Table 1).
Phenology: May to July.
Habitat and Geographical Distribution: On and inside hollow of trunks of living Dicymbe (Fabaceae), also on decorticated wood, known only from Guyana
Inocybe rhodella Matheny, Aime, & M.E. Smith, sp. nov. Figs. 6, 14. MB 563642.
Pileus 10-15 mm diam, plane with a broadly umbonate disc, smooth and unbroken at the center, outwards radially fibrillose, not at all squamulose, rimose, no velipellis observed; reddish brown (8E6) (2.5YR 4/4-4/2 or 5YR 4/3) at the center and towards the margin, light reddish brown to pinkish gray (5YR 6/3-6/2) between spreading fibrils; context pallid, thickest under the disc, very thin elsewhere, not changing upon exposure, odor not recorded. Lamellae sinuate to adnexed, subdistant, 28-30 L, with several tiers of lamellulae, pale red to pinkish gray after expansion (6C4) (2.5YR 6/2-5YR 6/2) or Light Vinaceous-Fawn to Vinaceous-Fawn, edges pallid and weakly fimbriate, ventricose. Stipe 25-30 x 3.0 mm, central, terete, enlarged slightly towards the base, the bulb of which is swollen but not marginate; base covered with pallid mycelium, apex scurfy-pruinose, elsewhere with a finely-fibrillose appearance; pallid with pale pinkish or pinkish-red tones at apex and just above the base; context white in the bulb.
Basidiospores (6.0-) 6.5-6.8-7.0 (-7.5) ± 0.40 x (5.0-) 5.5-5.7-6.0 ± 0.30 µm, Q = 1.08- 1.20-1.27 ± 0.06 (n=20/1), gibbous about a polygonal outline, mostly with 8-9 moderate to small-sized nodules, light yellowish brown. Basidia 4-sterigmate, cylindrico-clavate, hyaline, mostly 22-24 x 8 µm. Pleurocystidia mostly 53-67 x 14-19 µm, often lageniform but at times subfusiform or subcylindric with rounded base (lacking a distinct basal pedicel), apices rounded or obtuse, crystals mostly absent, very thick-walled with walls often 4.0- 5.0 µm thick, hyaline, frequent. Cheilocystidia similar to pleurocystidia but often shorter or ventricose to broadly lageniform, mixed with short, clavate, hyaline paracystidia. Lamellar trama parallel, hyaline, composed of cylindric to inflated hyphae that are mostly 5-12 µm diam. Stipitipellis covered with caulocystidia down to base, frequent at apex to the center, less frequent below, similar in shape to cheilocystidia but similar in size to pleurocystidia, mixed with hyaline cauloparacystidia. Pileipellis composed of a suprapellis characterized by interwoven colorless, cylindric hyphae mostly 4-6 µm diam, occasionally mixed with wider diameter weakly incrusted hyphae; subpellis with an interwoven appearance but light cinnamon-brown in mass, many hyphae here incrusted, up to 16 µm diam. Clamp connections present.
Material examined:
GUYANA: Region 8 Potaro-Siparuni - Pakaraima Mountains, Upper Potaro River Basin, elevation 710 - 750 m, on ground, vicinity of base camp, MCA3917 (holotype BRG, isotype TENN066450), leg. M.C. Aime, 15-May-2010 [JN642230-ITS, JN642234-LSU].
Observations: Inocybe rhodella differs from I. marginata morphologically by the lack of a distinctive marginate bulb and smaller size. The two are separated distinctly by pair-wise comparison of LSU nucleotide sequences (97% similar). Both species differ readily from I. matrisdei Singer (see below), a Bolivian species, by their smaller spores. The latter has yet to be recorded from Guyana.
Phenology: May.
Habitat and Geographical Distribution: on ground under Dicymbe, known only from Guyana.
Additional material examined
Inocybe matrisdei Singer, Sydowia 15: 113. 1962 [1961]. TYPE: BOLIVIA: Prov. Manuripi, Dpto. Pando, B2285 (holotype!, LIL).
Pileus 20 mm diam, reddish grayish brown to tawny, rimose and at the same time squamulose, with a fine and slight whitish overcast, convex, obtuse, lacking any conspicuous umbo, context white, unchanging, odor none. Lamellae argillaceous with paler or pallid edge, medium broad, close, sinuateadnexed. Stipe 27 x 3.5 mm, brownish pink, entirely pruinose, with a margined bulb at base, hollow or becoming hollow in age.
Basidiospores 7.5-8.5-9.0 (-10.0) ± 0.60 x (6.0-) 6.5-7.0-7.5 (-8.0) ± 0.47 µm (n=20/1), Q = (1.06) 1.13-1.20-1.33 (-1.42) ± 0.01, gibbous, coarsely nodulose with mostly 10-13 moderatesized nodules about an elliptic outline (similar to I. stellatospora), yellowish brown. Basidia not studied. Pleurocystidia 36-53 x 13-19 µm, most often ventricose with short necks and usually tapered towards the base, frequent, thick-walled, walls 2.0-4.0 µm thick, hyaline, apices with crystals at times. Cheilocystidia and Lamellar trama not studied. Stipitipellis with numerous caulocystidia that are mostly utriform to saccate and lacking any pronounced necks.
Material examined.
BOLIVIA: Prov. Manuripi, Dpto. Pando, Madre de Dios River gallery forest, near Conquista, on the ground in marginal forest, inundation zone, B2285 (holotype LIL), leg. R. Singer, 27-Mar-1956.
Observations: The above macroscopic description is taken from Singer (1962) and modified here for formatting. The anatomical observations are our own and based on examination of the holotype (B2285, LIL). The type material consists of a single or one and a half specimens with scattered patches of white moldy growth mostly on the pileus. The material is fragmented, fragile, and in poor condition. The type was examined by E. Horak in the mid-1970s, but to our knowledge this is the first published re-examination.
Our basidiospore measurements are somewhat larger than originally recorded by Singer (1962). Singer reported basidiospores 6.8-8.3 x 5.5-7.2 µm. For I. matrisdei we observed a mean of 8.5 x 7.0 µm, dimensions larger than in I. marginata and I. rhodella. The basidiospores of I. matrisdei are more coarsely-nodulose as well. The Bolivian species is otherwise quite similar to I. marginata and shares the possession of a marginate bulb, but differs in outward appearance by the squamulose pileus surface and lack of pink tinged lamellae characteristic for both I. marginata and I. rhodella. All three species share pink or reddish tones to the stipe and pileus.
Phenology: March.
Habitat and Geographic Distribution: on ground in tropical forest, Bolivia
Acknowledgements
The authors thank the following sources for funding: NSF DEB-0949517 (PBM), W.K. Smith and NSF DEB-0732968 (MCA), Harvard University Herbaria (MES), and the National Geographic Society's Committee for Research and Exploration, and NSF DEB- 0918591 (TWH). Field assistance in Guyana was provided by M. Chin, C. Andrew, V. Joseph, F. Edmond, L. Edmond, and L. Williams. Research permits were granted by the Guyana Environmental Protection Agency. The authors thank Aaron Wolfenbarger, Sarah Sprague, and R. Kaur for laboratory assistance. We also acknowledge herbarium staff and director, Ana María Frías, at LIL in Tucumán, Argentina, and staff at INPA in Manaus, Brazil, for access to type materials of Rolf Singer. We also thank Marisol Sanchez (University of Tennessee) for a translation of the summary into Spanish. This paper is number 180 in the Smithsonian Institution's Biological Diversity of the Guyana Shield Program publication series.
References
1. Alexander I. & M.-A. Selosse 2009. Mycorrhizas in tropical forests: a neglected research imperative. New Phytol. 182: 14-16. [ Links ]
2. Alvarado P., J. L. Manjon, P. B. Matheny & F. Esteve- Raventós 2010. Tubariomyces, a new genus of Inocybaceae from the Mediterranean region. Mycologia 102: 1389-1397. [ Links ]
3. Cortez V. G. & G. Coelho 2005. Additions to the mycobiota (Agaricales, Basidiomycetes) of Rio Grande do Sul, Brazil. Iheringia, Ser. Bot., Porto Alegre 60: 69-75. [ Links ]
4. Dennis R. W. G 1953. Les Agaricales de l'ile de la Trinité: rhodosporae-ochrosporae. Bull. Trim. Soc. Mycol. Fr. 69: 145-198. [ Links ]
5. Dennis R. W. G. 1970. Fungus flora of Venezuela and adjacent countries. Kew Bull., Add. Ser. 3. HMSO, London. [ Links ]
6. Guzman G. 1982. New species of fungi from the Yucatan Peninsula. Mycotaxon 16: 249-261. [ Links ]
7. Henkel T. W., M. C. Aime, M. Chin, S. L. Miller, R. Vilgalys & M. E. Smith 2011. Ectomycorrhizal fungal sporocarp diversity and discovery of new taxa in Dicymbe monodominant forests of the Guiana Shield. Biodiversity and Conservation, in press. [ Links ]
8. Henkel T. W., M. C. Aime & S. L. Miller 2000. Systematics of pleurotoid Russulaceae from Guyana and Japan, with notes on their ectomycorrhizal status. Mycologia 92: 1119-1132. [ Links ]
9. Henkel T. W., M. E. Smith & M. C. Aime 2010. Guyanagaster, a new wood-decaying sequestrate genus of Agaricales from the Guiana Shield. Am. J. Bot. 97: 1474-1484. [ Links ]
10. Judge B., J. F. Ammirati, G. Lincoff , J. H. Trestrail & P. B. Matheny 2010. Ingestion of a newly described North American mushroom species from Michigan resulting in chronic renal failure: Cortinarius orellanosus. Clin. Toxicol. 48: 545- 549. [ Links ]
11. Kornerup A. & J. H. Wanscher 1978. Methuen handbook of colour. 3rd ed. Eyre Methuen, London. [ Links ]
12. Larsson E., M. Ryberg, P.-A. Moreau, A. D. Mathiesen & S. Jacobsson 2009. Taxonomy and evolutionary relationships within species of section Rimosae (Inocybe) based on ITS, LSU and mtSSU sequence data. Persoonia 23: 86-98. [ Links ]
13. Matheny P. B., M. C. Aime, N. L. Bougher, B. Buyck, D. E. Desjardin, E. Horak, B. R. Kropp, D. L. Lodge, J. M. Trappe & D. S. Hibbett 2009. Out of the palaeotropics? Historical biogeography and diversification of the cosmopolitan mushroom family Inocybaceae. J. Biogeography 36: 577- 592. [ Links ]
14. Matheny P. B., M. C. Aime & T. W. Henkel 2003. New species of Inocybe from Dicymbe forests of Guyana. Mycol. Res. 107: 495-505. [ Links ]
15. Matheny P. B., Z. Wang, M. Binder, J. M. Curtis, Y.-W. Lim, R. H. Nilsoon, K. W. Hughes, V. Hofstetter, J. F. Ammirati, C. L. Schoch, E. Langer, G. Langer, D. J. McLaughlin, A. W. Wilson, T. Frøslev, Z.-W. Ge, R. W. Kerrigan, J. C. Slot, Z.-L. Yang, T. J. Baroni, M. Fischer, K. Hosaka, K. Matsuura, M. T. Seidl, J. Vauras & D. S. Hibbett 2007. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenet. Evol. 43: 430-451. [ Links ]
16. Meijer, A. R. de 2006. Preliminary list of the macromycetes from the Brazilian state of Parana. Bol. Mus. Bot. Municipal (Curitiba) 58: 1-55. [ Links ]
17. Miller, J. S., V. A. Funk, W. L. Wagner, F. Barrie, P. C. Hoch & P. Herendeen 2011. Outcomes of the 2011 botanical nomenclature section at the XVIII International Botanical Congress. PhytoKeys 5: 1-3. [ Links ]
18. Munsell Soil Color Charts 1954. Munsell Color Company, Baltimore 18, Maryland, USA. [ Links ]
19. Pegler D. (1983) Agaric flora of the Lesser Antilles. Kew Bull. Ser. IX: 683 pp. [ Links ]
20. Rick J. 1961. Basidiomycetes eubasidii in Rio Grande do Sul - Brasilia. Iheringia, Sér. Bot. (8): 296- 450. [ Links ]
21. Ridgway R. 1912. Color standards and color nomenclature (published by the author). Washington, D.C., USA. [ Links ]
22. Singer R. 1953. Type studies on Basidiomycetes VI. Lilloa 26: 57-159. [ Links ]
23. Singer R. 1962. Monographs of South American Basidiomycetes, especially those of the east slop of the Andes and Brazil. 4. Inocybe in the Amazone region, with a supplement to part 1 (Pluteus in South America). Sydowia 15: 112-132. [ Links ]
24. Singer R., I. Araujo & M. H. Ivory (1983) The ectotrophically mycorrhizal fungi of the neotropical lowlands, especially central Amazonia. Beih. Nova Hedwigia 77: 1-339. [ Links ]
25. Smith M. E., T. W. Henkel, M. C. Aime, A. K. Fremier & R. Vilgalys 2011. Ectomycorrhizal fungal diversity and community structure on three co-occurring leguminous canopy tree species in a Neotropical rainforest. New Phytol. doi: 10.1111/j.1469-8137.2011.03844.x [ Links ]
26. Smith M.E., G. W. Douhan & D. M. Rizzo 2007. Intraspecific and intrasporocarp ITS diversity of ectomycorrhizal fungi as assessed by ITS sequencing of bulked roots and sporocarps from a Quercus woodland. Mycorrhiza 18: 15-22. [ Links ]
27. Thiers B. [continuously updated]. Index Herbariorum: a global directory of public herbaria and associated staff. New York Garden's Virtual Herbarium. http://sweetgum.nybg.org/ih/ [ Links ]
28. Wartchow F., L. C. Maia & M. A. Q. Cavalcanti 2008. Inocybe martinica: a new record from South America and type studies of allied species from the Lesser Antilles. Mycotaxon 104: 43-49. [ Links ]
Original recibido el 21 de Septiembre de 2011;
primera decisión: 1 de Noviembre de 2011;
aceptado el 1 de Noviembre de 2011.
Editor responsable: Carlos Urcelay.

