










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBAG. Journal of basic and applied genetics
versión On-line ISSN 1852-6233
BAG, J. basic appl. genet. v.20 n.1 Ciudad Autónoma de Buenos Aires ene./jun. 2009
ARTÍCULOS ORIGINALES
Relationships among agronomic traits and seed yield in pea
Espósito, María Andrea 1*, Martin2, Eugenia Alejandra, Cravero2, Vanina Pamela, Liberatti2, David, López Anido1, Fernando Sebastián & Cointry1, Enrique Luis.
1Department of Genetic and Plant Breeding -Rosario National University, Zavalla. Argentina.
2CONICET –Zavalla. Argentina.
*Corresponding author at: Genetic and Plant Breeding Department, Rosario National University (UNR), CC 14, Zavalla S2125ZAA, Argentine. Tel: 54-341-4970080; fax: 54-341-4970085.
e-mail address: mesposi@unr.edu.ar
ABSTRACT
The evaluation of selection criteria using correlation coefficients, multiple regression and path analysis was carried out for a period of two years on forty pea genotypes. The correlation analysis revealed that grain yield had genotypic relationships with numbers of pods, seeds per plot, length of the internodes and plant height in 2007 and also with grain diameter, length and width of leaflets and number of nodes at the first pod in 2008. The highest positive direct effects in 2007 were length of the internodes (0.68), seeds per plot (0.38) and numbers of pods (0.26). Length leaflets exhibited a negative direct effect (-0.46). The highest positive indirect contribution of plant height mediated by length of the internodes was 0.50. The highest negative indirect contribution was pod length via length of the internodes (-0.35). In 2008, the highest positive direct effects were seeds per plot (0.67), width leaflets (0.33) and numbers of pods (0.25). Length leaflets presented the highest negative direct effect (-0.34). The indirect effects were observed via seeds per plot, length and width leaflets; therefore numbers of pods and seeds per plot can be used for indirect selection. The parameter estimated showed that number of pods and seeds, and pod length determined the yield during 2007 and number of pod and seeds, and grain diameter during 2008. The R2 values for both models were 0.60 and 0.89 respectively. The number of pod and seeds per plot were the main components of seed yield, having the maximum direct effects on this trait. These results might be used as selection criteria in order to increase the selection efficiency in pea breeding programs.
Keywords: Pisum sativum L.; Yield components; Genetic correlation; Stepwise regression; Path coefficient.
RESUMEN
El coeficiente de correlación, la regresión múltiple y el análisis de sendero se emplearon para determinar criterios selectivos utilizando durante dos años, 40 genotipos de arveja. El rendimiento se correlacionó genotípicamente con número de vainas, semillas/parcela, longitud del entrenudo y altura de planta en 2007 y también con diámetro de grano, largo y ancho de folíolo y número de nudos a primera vaina en 2008. Durante 2007 los rasgos que mostraron efectos positivos directos elevados fueron longitud de entrenudos (0,68), semillas/parcela (0,38) y número de vainas (0,26). La longitud del folíolo exhibió un efecto directo negativo (-0,46). La mayor contribución indirecta positiva fue la de altura de planta vía longitud de entrenudos (0,50) y la mayor contribución negativa indirecta fue longitud de vaina vía longitud de entrenudos (-0,35). Durante 2008, efectos positivos directos elevados fueron para semillas/parcela (0,67), ancho del folíolo (0,33) y número de vainas (0,25). La longitud de folíolo presentó el mayor efecto directo negativo (-0,34). Los efectos indirectos fueron observados vía semillas/parcela, longitud y ancho de folíolos, por lo que el número de vainas y semillas por parcela, pueden ser utilizadas para selección indirecta. La correlación múltiple mostró que el número de vainas y semillas y la longitud de la vaina determinaron el rendimiento durante el 2007 y el número de vainas y semillas y el diámetro del grano durante el 2008. Los valores de R2 para los dos modelos fueron 0,60 y 0,89, respectivamente. El número de vainas y semillas por vaina fueron los principales componentes del rendimiento, teniendo máximos efectos directos sobre este carácter. Estos resultados podrían ser utilizados como criterios de selección a fin de aumentar la eficiencia en programas de mejora de arveja.
Palabras clave: Pisum sativum L.; Componentes de rendimiento; Correlación genética; Regresión múltiple; Análisis de sendero.
INTRODUCTION
Pea is an Old World cool season annual legume crop whose origins trace back to the primary centre of origin in the Near and Middle East. Their presence is found in remains at sites in Southern Europe soon after (Zohary and Hopf, 1973). Although it cannot be proved, it is highly likely that peas were consumed both as fresh vegetable and as cooked forms. The genus Pisum comprises only a small number of diploid taxa (2n=14) Ben-Ze'ev and Zohary, 1973). The increasing demand for protein-rich raw materials for forage or intermediary products for human nutrition have led to a greater interest in this crop as a protein source (Santalla et al., 2001). Traditionally, plant breeders have optimized yield largely by empirical selection with little regard for the physiological processes involved in yield increase. More recently, strategies to optimize yield in pea have focused on the physiological mechanisms involved in the seed setting and fruit filling. However, selection of high yielding cultivars via specific traits requires knowledge of not only final yield but also the many compensation mechanisms among yield components resulting from changing genotypic, environmental and management factors. Grain yield of pea is a quantitative trait which is affected by many genetic and environmental factors (Singh and Singh, 2004; Ceyhan and Avci, 2005; Ranjan et al., 2006).
Since grain yield is a complex trait, indirect selection through correlated, less complex and easier measurable traits would be an advisable strategy to increase the grain yield.
Efficiency of indirect selection depends on the magnitude of correlations between yield and target yield components.
In agriculture, correlation coefficients in general show associations among characteristics. It is not sufficient to describe this relationship when the causal association among characteristics is needed (Toker and Cagirgan, 2004). If there is genetic correlation between two traits direct selection of one of them will cause change in the other. When more than two variables are involved, the correlations per se do not give the complete picture of their interrelationships (Fakorede and Opeke, 1985). The path analysis has been used by plant breeders (Indu Rani et al., 2008, Togay et al., 2008, Ali et al, 2009) to assist in identifying traits that are useful as selection criteria to improve crop yield because it identifies the causes, measures the relative importance of the association, and is used to determine the amount of direct and indirect effect of the causal components on the effect component (Dewey and Lu, 1959). Multiple regression and path coefficient analyses are particularly useful for the study of cause-and-effect relationships because they simultaneously consider several variables in the data set to obtain the coefficients. Determination of correlation and path coefficients between yield and yield criteria is important for the selection of favorable plant types for effective pea breeding programs.
The objective of this study was to evaluate selection criteria in pea breeding programs by means of correlation, multiple regression and path coefficient analysis.
MATERIALS AND METHODS
The present investigation for correlation, multiple regression and path analysis studies of morphological traits of pea was conducted in the research station of Faculty of Agriculture of Rosario University, Argentine (33°1’ S and 60° 53’ W), in 2007 and 2008. The experimental material consists of forty genotypes of pea from North and South America, Europe, Australia, India and local breeding programs material. These genotypes were planted in the field based on the randomized complete block design with three replications in each of the years. Plots were arranged ten rows of 2 m length with inter and intra row spacing of 70 and 10 cm, respectively. All other agronomic practices were kept uniform. Characters were evaluated on ten randomly selected plants in the three mid-rows of plots. Length (LS) and width (WS) of stipule, leaflets (LL, WL) and pod (LP, WP), length of the internodes (LI), plant height (PH), number of nodes at the first pod (NFP), and numbers of days to flowering (DF). Numbers of pods (NP) and seeds per plot (NS), yield (Y) and grain diameter (GD) were measured at the dry seed stage. Therefore thirty plants per genotype were included in this analysis.
Length and width of stipule, leaflets, and pods, length of the internodes and plant height were recorded in centimeters and the number of nodes at the first flower was counted with the average of three plants randomly selected in the center of rows. The yield was estimated in grams per plot with total dry weight of plants in harvest. Seeds per plot were counted. The days to flowering were estimated as number of days from sowing to the date on which at least 50% of the plants in the plot had started to flowering. All these traits were included in the path and correlation analyses and multiple regressions. The multiple regression model was Y= β0 + β1X1 + β2X2 + ...+ βiXi+...+ βnXn + ε, were Y represents the yield (dependent variable), βi and Xi the multiple regression coefficient and the recorded value of the iesim explanatory (independent) variable and ε indicated the experimental error. The model for all traits included random genotype effects. Genetic correlations between traits were determined according to the method of Know and Torrie (1964): rg= Covgij/ (σ2gi* σ2gj) where Covgij, σ2gi and σ2gj are the estimates of covariance and variance for traits i and j respectively. Multiple regression, step-wise regression and genetic correlations were obtained by the GENES software (Cruz, 2001) and path analysis by InfoGen software (Balzarini and Di Renzo, 2003).
RESULTS AND DISCUSSION
Correlation coefficient study
Means values and standard errors for each morphological trait are presented in Table I. The genotypic correlation coefficients were higher as compared to phenotypic correlation coefficient in most of the cases (Table II). This indicates greater contribution of genotypic factor in the development of the character associations.
Table I: Genotypic (rg) and phenotypic (rp) correlation coefficients between different traits in pea during 2007 (above diagonal) and 2008 (below diagonal).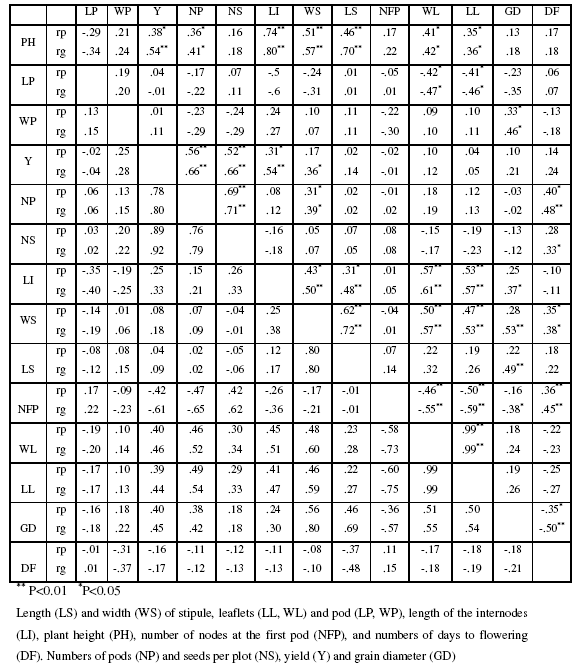
Table IIa: The direct and indirect contribution of different traits to yield in pea during 2007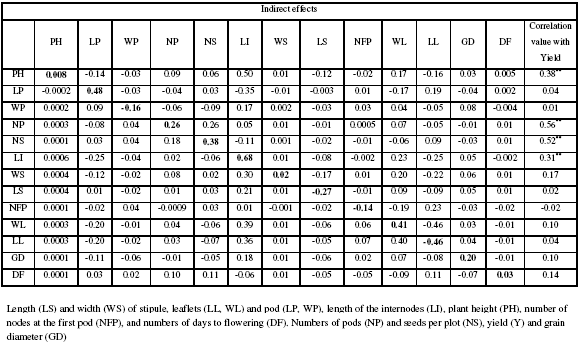
Table IIb: The direct and indirect contribution of different traits to yield in pea during 2008
Significant positive genotypic correlation of days to flowering with numbers of pods (0.48**), seeds per plot (0.33*), stipule width (0.38*) and of nodes at the first pod (0.45**) were observed. Negative genotypic correlation of days to flowering with grain diameter (-0.50**) was also observed and this fact is important to develop cultivars with short duration cycle and high grain diameter. Similarly, negative association of nodes at the first pod with length and width of stipule (-0.55** and -0.59**, respectively) were found.
Positive genotypic correlation of plant height with internode length (0.80**) and stipule length (0.70**) were also observed. Pod length had significant and negative genotypic correlation with leaflet length and width (-0.47** and 0.46**, respectively).
Grain yield had highly significant positive genotypic correlation with total number of pods (0.66**), numbers of seeds per plot (0.66**) and internode length (0.33). These positive and strong associations with grain yield revealed the importance of these characters in determining grain yield and indicate that selection for either or both of these traits would result in superior yield (Pandey and Gritton, 1975).
Khanghah and Sohani (1999) Rajanna et al., (2000) and Singh and Yadava (2000) also reported similar findings for different parameters in soybean. According to Siahsar and Rezai (1999) number of pod per plant had the greatest genotypic correlation with seed yield in soybean which also confirms the results of present investigation. Pandey and Gritton (1975) founded that the plant height was often positively correlated with pods/plant, seeds/plant, and yield/plant, indicating superior yielding ability of taller plants in pea.
Path coefficient analysis
Yield is a complex character with polygenic inheritance that, from a crop physiology perspective, is the culmination of a series of processes (phenological and canopy development, radiation interception, biomass production and partitioning) that are driven by environmental influences (Charles-Edwards, 1982). A genotype’s ultimate performance is determined by how it integrates genotype and environmental influences. The end result is seed yield, which has often been described as the product of its components: number of plants per unit area, number of seeds per unit area (number of pods per plant, number of seeds per pod), and mean seed weight (Moot and McNeil, 1995). These yield components show interdependence or plasticity (Wilson, 1987). For example, compensation is observed between the number of pods per plant and number of seeds per pod (Moot and McNeil, 1995), or between seed number and seed weight (Sarawat et al., 1994). The path coefficient analysis initially suggested by Wright (1921) and described by Dewey and Lu (1959) allows partitioning of correlation coefficient into direct and indirect effects of various traits towards dependent variable and thus helps in assessing the cause - effect relationship as well as effective selection. Thus, the path analysis plays an important role in determining the degree of relationship between yields and yield components effects and also permits critical examination of specific factors that provide a given correlation.
The effects of yield components via path analysis were given in Table IIIa-b. In this table only important correlated traits with yield were examined. In 2007, the highest positive direct effects on yield were LI (0.68), NS (0.38) and NP (0.26) while PH (0.008) was positive but low. Meanwhile, LL exhibited an import negative direct effect (-0.46). The highest positive indirect contribution of PH via LI was 0.50 followed by NP via NS (0.26). It also observed that highest negative indirect contribution was LP via LI (-0.35).
Table IIIa: The step-wise parameters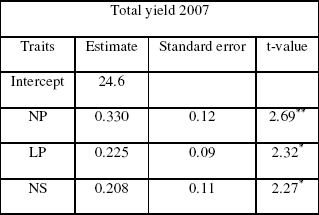
Table IIIb: The step-wise parameters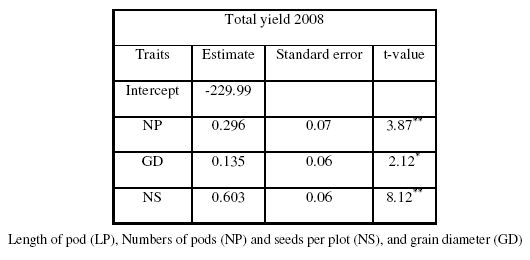
In 2008, the highest positive direct effects on yield were NS (0.67), WL (0.33) and NP (0.25) meanwhile LL presented the highest negative direct effect (-0.34). The indirect effects were observed via NS, LL and WL. Because in both years, NP and NS shown the highest positive direct effects on yield, clearly indicated that these can be used for indirect selection because LI, LL and WL are influenced by environmental condition.
Multiple Regressions
The stepwise regression variance analysis results indicated that model was significant to perform the stepwise regression analysis for yield (Table IVa-b). The parameter estimated showed that NP, LP and NS determined the yield during 2007 and NS, NP and GD during 2008. The R2 values for both models were 0.60 and 0.89 respectively. Thus, the yield could be increased directly through NP and NS but indirectly through LI, LL and WL because taller plants involves a larger number of pods per plant, seeds per plant and yield per plant and the leaf area is one of the most essential processes, such as one of the physiological determinants of plant growth is the efficiency of the leaves with which the intercepted light energy is used in the production of new dry matter (Uzun, 1999). Moreover, leaf area is an indicator of photosynthetic capacity and growth rate of a plant and its measurement is of value in studies of plant competition for light and nutrients. Common measurements in pea include leaf length and leaf width is the leaf area a good indicator of yield potential because both traits are positively correlated but influenced in opposite ways the number of seeds which is an important component of yield.
CONCLUSION
The different analyses carried out coincide in that the number of pod and seeds per plot were the main yield components having maximum direct effects on seed yield. The results identify these traits as selection criteria in further studies in order to increase the selection efficiency in pea breeding program.
REFERENCES
1. Ali, M.A.; Nawad, N.N.; Abbas, A.; Zulkiffal, M. & Sajjad, M. (2009). Evaluation of selection criteria in Cicer arietinum L. using correlation coefficients and path analysis. Australian J. Crop Sci. 3(2):65-70. [ Links ]
2. Balzarini, M. & Di renzo, J. (2003). Infogen: Software para análisis estadísticos de marcadores genéticos. Facultad de Ciencias Agropecuarias. Universidad Nacional de Córdoba. Córdoba. [ Links ]
3. Ben-ze'ev, N. & Zohary, D. (1973). Species relationships in the genus Pisum L. Isr. J. Bot. 22:73-91. [ Links ]
4. Ceyhan, E. &. Avci, M.A. (2005). Combining ability and heterosis for grain yield and some yield components in pea (Pisum sativum L.). Pakistan Journal of Biological Sci. 8(10):1447-1452. [ Links ]
5. Charles-Edwards, D.A. (1982). Physiological determinants of crop growth. Academic Press, Sydney, Australia. [ Links ]
6. Cruz, C.D. (2001). Programa GENES: aplicativo computacional em genética estatística. Viçosa: Editora UFV. 648p. [ Links ]
7. Dewey, D.R. & Lu, K.H. (1959). A correlation and path coefficient analysis of components of crested wheat grass seed production. Agron. J. 51:515-518. [ Links ]
8. Fakorede, M.A.B. & Opeke, B.O. (1985). Whether factors affecting the response of maize to planting date in a tropical rainforest location. Exp. Agric. 21:31-40. [ Links ]
9. Indu Rani, C.; Veeraragathantham, D. & Sanjutha, S. (2008). Studies on correlation and path coefficient analysis on yield attributes in Root Knot Nematode Resistant F1 hybrids of tomato. J. Appl. Sci. Res. 4(3):287-295. [ Links ]
10. Khanghah, H.Z. & Sohani, A.R. (1999). Genetic evaluation of some important agronomic traits related to seed yield by Multivariate of soybean analysis methods. Iranian J. Agri. Sci. 30:807-816. [ Links ]
11. Know, S. & Torrie, J. (1964). Heritability and inter-relationship among traits of two soybean populations. Crop Sci. 4:196-198. [ Links ]
12. Moot, D.J. &. Mcneil, D.L. (1995). Yield components, harvest index and plant type in relation to yield differences in field pea genotypes of partial resistance to field epidemics of ascochyta blight of pea. Euphytica 86:31-40. [ Links ]
13. Pandey, S. & Gritton, E.T. (1975). Genotypic and phenotypic variances and correlations in peas. Crop Sci. 15:353-356. [ Links ]
14. Rajanna, M.P.; Viswanatha, S.R; Kulkarrni; R.S. & Ramesh, S. (2000). Correlation and path analyses in soybean Glycine max (L.). Crop Res. Hisar., 20(2):244-247. [ Links ]
15. Ranjan, S.; Kumar, M. & Pandey, S.S. (2006). Genetic variability in peas (Pisum sativum L.). Legume Research 29(4):311-312. [ Links ]
16. Santalla, M.; Amurrio, J. & De Ron, A. (2001). Food and feed potential breeding value of green, dry and vegetal pea germplasm. Can. J. Plant Sci. 8:601-610. [ Links ]
17. Sarawat, P.; Stoddard, F.L.; Marshall, D.R. & Ali, S.M. (1994). Heterosis for yield and related characters in pea. Euphytica 80:39-48. [ Links ]
18. Siahsar, B. & Rezai, A. (1999). Correlation and path coefficient analysis of morphological and phonological traits associated with yield in soybean (Glycine max (L.) Merr.). Iranian J. Agri. Sci., 30:685-696. [ Links ]
19. Singh, J.D. & Singh, I.P. (2004). Selection parameters for seed yield in field pea (Pisum sativum L.). Nat. J. Plant Improvement 6(1):51-52. [ Links ]
20. Singh, J. & Yadava, H.S. (2000). Factors determining seed yield in early generation of soybean. Crop Res. Hisar. 20:239-243. [ Links ]
21. Toker, C. & Cagirgan, M.I. (2004). The use of phenotypic correlations and factor analysis in determining characters for grain yield selection in chickpea (Cicer arietinum L.). Hereditas 140:226-228. [ Links ]
22. Togay, N., Y. Togay, B. Yildirin and Y. Dogan. 2008. Relationships between yield and some yield components in pea (Pisum sativum ssp arvense L.) genotypes by using correlation and path analysis. Afr. J. Biotechnol. 7(23):4285-4287. ISSN: 1684-5315. [ Links ]
23. Uzun S. & Celik, H. (1999). Leaf area prediction models (Uzçe-lik-I) for different horticultural plants. Turkish Journal of Agriculture and Forestry. 23:645-650. [ Links ]
24. Wilson, D.R. (1987). New approaches to understanding the growth and yield of pea crops. p. 23-28. In W.A. Jermyn and G.S Wratt (ed.). Peas: Management for quality. Spec. Publ. No. 6. Agron. Soc. of New Zealand. [ Links ]
25. Wright, S. (1921). Correlation and causation. J. Agric. Res.20:557-585.Zohary, D. & Hopf, M. 1973. Domestication of pulses in the old world. Science 182:887-894. [ Links ]
26. Zohary, D. & Hopf, M. 1973. Domestication of pulses in the old world. Science 182:887-894. [ Links ]
Recibido: 01-11-2009
Aceptado: 09-02-2010.

