










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBAG. Journal of basic and applied genetics
versión On-line ISSN 1852-6233
BAG, J. basic appl. genet. vol.21 no.2 Ciudad Autónoma de Buenos Aires jul./dic. 2010
Utilizar regiones heteróticas mantenidas por letales balanceados en el genoma del maíz se hace realidad
Ing. Agr. Dr. Juan Carlos Salerno, M. V. Kandus, R. Boggio Ronceros y D. Almorza
Instituto de Genética "Ewald A. Favret", C.C. 25 (1712) Castelar, Bs. As.
jsalerno@cnia.inta.gov.ar
RESUMEN
Para incrementar la eficiencia en la producción de semillas híbridas en maíz se necesitan altos rendimientos con un correlativo bajo costo de producción. Esto requiere una expansión de los conocimientos básicos de la herencia de caracteres que posibiliten desarrollar nuevas técnicas de mejoramiento. Los sistemas de letales balanceados permiten estudiar la contribución relativa de distintos segmentos heteróticos en el vigor híbrido dado que permiten lograr la heterocigosis quasi-permanente de una porción del genomio, mientras el resto del genoma se vuelve homocigota por autofecundaciones sucesivas. Estos sistemas pueden ser transferidos a líneas endocriadas para incrementar el rendimiento del grano y la producción de polen mediante el aumento del tamaño de la panoja. En los últimos 15 años se incorporaron segmentos heteróticos de tres líneas reguladas por distintos sistemas de letales balanceados (BLS) en líneas S5 derivadas de dos híbridos comerciales ACA 2000 y Cóndor de pedigree cerrado, con textura dura (flint), con la finalidad de aumentar el rendimiento en grano o el tamaño de la panoja. El análisis estadístico mostró un incremento significativo en el rendimiento en grano y el tamaño de la panoja de las líneas S5 derivadas de ambos híbridos comerciales, portando cada una de ellas segmentos heteróticos provenientes de tres líneas reguladas por BLS diferentes.
Palabras clave: Sistemas de letales balanceados; Segmentos heteróticos; Vigor híbrido.
ABSTRACT
In order to increase the efficiency of hybrid seed production in maize, it is necessary to achieve a high grain yield to reduce production costs. This goal requires an expansion of the basic knowledge of the inheritance of traits to develop new breeding techniques. The balanced lethal systems allow the study of the relative contribution of different chromosomal segments to hybrid vigour due to the heterozigosity of certain chromosomal sectors while the rest of the genome becomes homozygous through continuous selfing. In this way, these segments can be transferred to inbred lines in order to increase grain yield or pollen production through the increase of tassel size. In the last fifteen years, the transference of heterotic segments that belong to three lines regulated by different balanced lethal systems (BLS) to S5 flint lines derived from two commercial hybrids, (ACA 2000 and Cóndor with closed pedigree) was carried out, with the aim of increasing grain yield or tassel size. The statistical analysis showed a significant increase in grain yield and tassel size in the S5 flint lines of both commercial hybrids, carrying three different heterotic segments of the BLS lines.
Key Words: Balanced lethal systems; Heterotic segments; Hybrid vigour
DESARROLLO
Para incrementar la eficiencia en la producción de semillas híbridas en maíz se necesitan altos rendimientos con un correlativo bajo costo de producción. La importancia de disponer de líneas de alto rendimiento para la formación de híbridos queda resaltada en los estudios realizados en Estados Unidos de América (EUA), indicando que desde 1930 el incremento de rendimiento de los híbridos sobre los padres fue constante y que los mayores rendimientos se debieron a líneas paternas más productivas (Duvick, 1977, 2001). El logro de este objetivo requiere una expansión de los conocimientos básicos de la herencia de caracteres que posibiliten desarrollar nuevas técnicas de mejoramiento sobre materiales experimentales locales de textura dura (flint) especialmente, ya que en los últimos años hubo una incorporación de germoplasma de textura blanda (dentada) que provocó un deterioro en la calidad final del grano, lo cual genera problemas en los sectores vinculados a su industrialización y exportación.
La Genética Cuantitativa incluye el estudio de acciones e interacciones de numerosos factores genéticos y su interacción con los factores ambientales en cada etapa del ciclo de vida de las plantas. Tanto los factores citoplásmicos como nucleares pueden ser incluidos. El desarrollo de modelos genéticos cuantitativos teóricos utilizando conceptos mendelianos en Genética, incluyendo epistasis y ligamiento, comenzó en la segunda mitad del siglo pasado (Cockerham, 1954; Kempthorne, 1954; Schnell, 1963). También han sido desarrollados modelos genéticos para la estimación de parámetros genéticos y diseños de apareamiento (Comstock & Robinson, 1948; 1952; Griffing, 1956; Gardner & Eberhart, 1968), entre otros. A su vez, numerosos trabajos realizaron aportes al estado de conocimiento en genética cuantitativa y sus aplicaciones en el mejoramiento genético vegetal y animal (Carson, 1967; Wright, 1968; Wallace, 1970; Mather & Jinks, 1971; Wright, 1978; Harth, 1980; Falconer, 1981; Hallauer & Miranda, 1981; Gallais, 1990; Russell, 1991). Los modelos genéticos y los diseños de apareamiento han probado ser extremadamente útiles para entender mecanismos y acciones genéticas cuantitativas y para desarrollar sistemas de mejoramiento (Cockerham, 1963). El uso de modelos matemáticos, estadísticos y genéticos, la simulación computada, el uso de diseños de apareamiento especiales y diseños experimentales en ensayos a campo han sido útiles para obtener la información básica que se requiere para definir métodos y diseños de selección.
El descubrimiento del vigor híbrido en maíz es considerado una de las mayores innovaciones del siglo XX, y marca la consolidación de una verdadera industria de semillas. Estudios avanzados sobre la heterosis o vigor híbrido enfatizan que ésta no necesita estar basada en un gran número de genes sino de la combinación de variantes alélicas de unos pocos genes, por lo tanto habría una cantidad considerable de vigor híbrido como resultado de las interacciones entre dichos genes (Gustafsson, 1946; Whaley, 1964; Stuber, 1992; 1994; Crow, 1999). No obstante la aceptación general de la hipótesis de la dominancia para la heterosis, los recientes estudios de estimaciones fenotípicas y de análisis de QTLs (Quantitative Trait Loci) para rendimiento en grano muestran verdadera sobredominancia (Lu et al., 2002; 2003). Posibles mecanismos para la superioridad de los heterocigotas incluyen la acción de selección de un mejor balance metabólico (Mangelsdorf, 1952), un control metabólico de flujos (Kacser et al., 1981), o selección para genes modificadores que favorecen a los heterocigotas (Fisher, 1928).
El maíz ha sido la primera especie que permitió aprovechar la heterosis, es decir la superioridad de la F1 respecto a la media de los progenitores o a su progenitor más rendidor. Para lograr heterosis es imprescindible tener en cuenta la aptitud combinatoria (capacidad de una línea endocriada para dar descendencia híbrida caracterizada por la elevada expresión de un carácter o grupo de caracteres). En este sentido, la metodología tradicional aplicada al mejoramiento genético del maíz fue desarrollada en EUA, basada en una búsqueda aleatoria en donde los cruzamientos óptimos se hallan según una metodología de cruzar todas las líneas mejoradas entre sí. A mayor cantidad de cruzamientos, mayor número de híbridos que se pueden seleccionar por alto rendimiento y por otras características agronómicas. Esto implica la necesidad de partir de un volumen de material elevado para poder probar todas las combinaciones posibles, descartando en el proceso de selección la mayor parte del material. Kiesselbach (1951) estimó que de las 100.000 líneas de maíz que se habían ensayado por entonces, sólo unas 60 líneas serían promisorias y un par de ellas llegarían al final del proceso. En Argentina, Luna & Safont (1978) mostraron la falta de diversidad genética en el mejoramiento de maíz, que continúa en la actualidad, ya que los híbridos comerciales de mayor difusión están integrados por muy pocas líneas endocriadas de bajo rendimiento en grano y polen. En resumen, basándose en los conceptos mencionados en la técnica tradicional de obtención de híbridos de maíz importada de EUA para germoplasma de textura dentada, se observa que el número de líneas flint evaluadas en las pruebas de aptitud combinatoria ha sido muy alto, pero el número de líneas efectivamente usadas en la producción ha sido muy bajo.
Además de las 10 generaciones que se requieren para el desarrollo de las líneas endocriadas, las pruebas de aptitud combinatoria general y específica representan el 75% del costo de la semilla híbrida a comercializar. A esto se le suma el bajo rendimiento de las líneas madres, debido a la pérdida de vigor por endocría. Esto trae como consecuencia que para producir en gran escala el híbrido simple elegido, cuanto menor es el rendimiento de la línea madre, mayor será la superficie que se necesite sembrar para su multiplicación, encareciendo el costo final de producción de la semilla híbrida en aproximadamente un 25% por cada tonelada de disminución del rendimiento de la línea madre.
El estudio de la explotación de la heterosis en maíz es un área de interés esencial, tanto desde el punto de vista práctico, para su utilización a nivel comercial, como del teórico, para el estudio de los mecanismos implicados en su expresión. Las poblaciones base de selección son el punto de partida para la obtención de líneas. Los cambios que ocurren en las poblaciones debido a la selección natural y artificial dependen de la diversidad genética de las mismas. Los loci con mutaciones que tienen un efecto perjudicial (deletéreo) en forma homocigota, son parte de esa diversidad genética. Estos genes mutantes pueden aumentar la adaptabilidad de sus portadores heterocigotas y son llamados la "carga genética" de la población. La definición más clara de este término es la expresada por Freire- Maia (1963), que considera que la "carga genética" es el precio que debe pagar toda población para sobrevivir y reproducirse a lo largo de su existencia. La moneda de pago son los distintos tipos de letalidad que pueden presentarse en una población. El estudio de la carga genética permite conocer la diferenciación, el grado de adaptabilidad y al mismo tiempo da una medida de la variabilidad genética de una población. En este sentido, es muy importante considerar los mecanismos genéticos que pueden mantener la variabilidad genética, que constituye el componente esencial en el mejoramiento genético de las especies vegetales.
Las mutaciones clorofílicas, debido a su fácil identificación y a la frecuencia de aparición, han sido utilizadas como indicadores de la actividad mutagénica de las radiaciones ionizantes y algunos mutágenos químicos (Muller, 1950). Además, son apropiadas para el estudio simultáneo de la mutación, la selección y los sistemas de apareamiento en las poblaciones vegetales. La frecuencia de mutaciones recesivas, como son la mayoría de las mutaciones clorofílicas, dan una idea objetiva de la variación genética dentro de una población. Favret & Godeck (1959) midieron el índice de mutación espontánea en cebada y otras gramíneas arribando a la misma conclusión, destacando en un trabajo posterior el posible uso de la heterocigosis permanente en cebada (Favret & Ryan, 1966). Crumpacker (1967) hizo una revisión de la carga genética en varias especies vegetales y animales mostrando que la frecuencia de mutaciones recesivas es muy alta en diferentes poblaciones (salvajes y domesticadas).
El estudio de la carga genética en poblaciones de maíz fue llevado a cabo en profundidad en el Instituto de Genética "IGEAF" del INTA Castelar (CICVyA), en poblaciones locales de maíz de endosperma vítreo (flint), encontrando sistemas de letales balanceados caracterizadas por el mantenimiento del vigor (Salerno, 1981, 2007; Boggio et al., 1997). Por otro lado, el vigor híbrido constituye una definición operativa, dado que aún no se han dilucidado las bases fisiogenéticas de dicho fenómeno. No obstante, se sabe que la expresión del mismo depende de la condición heterocigota de una porción del genoma. Los sistemas de letales balanceados permiten estudiar la contribución relativa de distintos segmentos cromosómicos al vigor híbrido dado que permiten lograr la heterocigosis quasi-permanente de ciertos segmentos cromosómicos, mientras el resto del genoma se vuelve homocigota por autofecundaciones sucesivas.
La utilización de los sistemas de letales balanceados que se pueden hallar en distintos sectores del genoma de maíz es considerada una innovación biotecnológica y tiene la ventaja de evaluar fácilmente el impacto del segmento con respecto a caracteres de interés agronómico, ya que éste se mantiene heterocigota a diferencia del resto del genoma y además, permite el aprovechamiento comercial de las líneas obtenidas debido a la fijación del segmento heterocigota. De esta manera, líneas endocriadas de las compañías semilleras que tienen buena aptitud combinatoria, pero bajo rendimiento en grano o producción de polen pueden ser mejoradas y aprovechadas. Por otro lado, el entendimiento de los factores involucrados en el mantenimiento de los genes letales en maíz resulta interesante desde el punto de vista teórico y práctico. En este sentido, el número total de genes involucrados en los letales clorofílicos no se conoce con exactitud. Emerson, Beadle & Fraser (1935) mencionan aproximadamente 90 genes. Weijer (1952) compila un número de 120 genes, mientras que Riman (1963) menciona 180 genes. Se considera que 0.00001 es un valor razonable de mutación espontánea (Crow, 1948) que coincidiría con un número de 200 genes para este carácter. El ligamiento de genes letales con otros genes que tienen un efecto favorable en caracteres de interés agronómico, como el rendimiento del grano en maíz, tenderá a mantener los genes letales en la población. El siguiente modelo puede explicar esto:
WA1 > WA2 > wA2.
WA2 WA1 wA2
→ disminuye la aptitud. en el cual w es un gen recesivo (albino) A1, es un alelo con una aptitud del locus que suma 0 a la aptitud del genotipo y A2 es un alelo que suma una cantidad positiva a la aptitud del genotipo. El estado heterocigota en ambos loci ligados produce el genotipo con más ventaja; de esta manera, el gen w tenderá a permanecer en la población hasta que la recombinación produzca el genotipo de mayor aptitud, WA2/WA2 y WA2/wA2, lo cual comenzará luego a reemplazar al doble heterocigota y disminuir la frecuencia de w. Si A2 muestra alguna dominancia sobre A1, o si A1 y A2 tienen sobredominancia en la combinación, el proceso de reemplazo para el gen será lento, permaneciendo muchas generaciones en la población.
El ligamiento entre estos genes clorofílicos con otros que regulan procesos de crecimiento, y la distribución de este tipo de letales en diferentes cromosomas del maíz, hace que la eliminación de plantas que segregan letales clorofílicos, como ocurre normalmente en los programas de mejoramiento genético, tienda a remover indefectiblemente genes favorables que están en el mismo cromosoma (Lindstrom, 1920; Wentz & Goodsell, 1929; Jones,1945; 1952; Gustafsson, 1946; 1947; 1953; Band et al., 1961; 1963; Apirion et al., 1961; Redei, 1962; Allard et al., 1964).
Todas estas evidencias llevarían a considerar que la sobredominancia asociada con ligamiento es un factor que puede involucrarse en el mantenimiento de genes letales en poblaciones autógamas como cebada, trigo, sorgo y, además, en ciertos casos es posible detectar su efecto contra un espectro genético relativamente heterocigota. Los procesos involucrados en el mantenimiento de los genes letales incluyen la mutación, migración, selección para varios tipos, deriva génica, ligamiento y endocría. Estos factores interactúan entre sí y con los factores biológicos y fisiológicos del ambiente en procesos complejos, determinando la estructura genética poblacional para el mejoramiento genético.
Teniendo en cuenta estos conceptos se evaluó la incorporación de segmentos heteróticos de tres líneas reguladas por distintos sistemas de letales balanceados (BLS) en dos líneas de maíz de pedigree cerrado, derivadas de los híbridos comerciales ACA 2000 y Cóndor. Para ello se midieron el rendimiento en grano y algunas variables relacionadas con el mismo (como longitud de la espiga, peso de 100 semillas y profundidad del grano) para la línea materna y la producción de polen a través de la longitud y número de ramificaciones de la panoja, para la línea paterna en las líneas endocriadas per-se de tipo flint, que presentaban bajo rendimiento en grano y poca producción de polen. La incorporación mediante retrocruza de segmentos heteróticos que tienen un efecto favorable sobre distintos caracteres de importancia agronómica, permitiría contribuir en forma económica en los programas de mejoramiento públicos y privados.
En un trabajo previo se encontró que el segmento heterótico de una de las líneas de maíz paterna denominada "LPBL1A" de endosperma flint fue localizado en las cercanías del centrómero del cromosoma 6, con un 6% de recombinación (Salerno et al., 1992). En coincidencia con estos resultados, una publicación posterior del grupo del "Instituto Sperimentale per la Cerealicoltura", Bergamo-Italia (Ajmone-Marsan et al., 1995), encontró un QTL (Quantitative Trait Loci) para la producción de grano en la región cromosómica cercana al centrómero del cromosoma 6 de maíz. Este QTL posee un Lod Score de 7,4 que explicó un 24,5% del total de la variación fenotípica detectada para la producción de grano y cuyo efecto promedio de sustitución alélica fue de 1 tonelada por hectárea. Estos resultados confirman la importancia de la región del genoma involucrada en nuestro trabajo.
La asociación entre caracteres que involucren el rendimiento en grano es un aspecto importante para tener en cuenta en los programas de mejoramiento, ya que los cambios por selección en un carácter dado pueden afectar positiva o negativamente a otros caracteres (Vencovsky & Barriga, 1992). Hallauer & Miranda (1981) realizaron un resumen de varios estudios en maíz, encontrando que las correlaciones de distintos atributos con el rendimiento fueron mayores para caracteres relacionados con la espiga (como longitud y diámetro de espiga, profundidad del grano, peso del grano, número de hileras de granos por espiga). De estos caracteres, la profundidad del grano fue el más asociado al rendimiento; sin embargo, ningún carácter por sí solo posee las condiciones necesarias para realizar una selección indirecta. A su vez, Brandolini & Brandolini (2001) encontraron correlaciones positivas y significativas entre longitud de espiga - altura de planta, longitud de espiga - diámetro de espiga y altura de planta - profundidad del grano, entre otras.
Algunos estudios demostraron que el número de ramificaciones de la panoja tiene un rango de heredabilidad de 46% a 89%, estableciendo ocho factores genéticos involucrados en la determinación de dicho carácter (Mock & Schuetz, 1974). Valores similares de heredabilidad fueron determinados por Schuetz & Mock (1974) y Geraldi et al. (1985). Se encontraron tres QTLs para el ángulo de las ramificaciones y seis QTLs para el número de ramificaciones de la panoja, asociados a la producción de polen (Mickelson et al., 2002). Berke & Rocheford (1999), encontraron seis QTLs para el ángulo de las ramificaciones, tres QTLs para el número de ramificaciones de la panoja y siete QTLs para peso de la panoja.
Los resultados obtenidos muestran la contribución de los segmentos regulados por sistemas de letales balanceados hallados en las líneas paternas LP BLS14, LPB LS16 y LP BLS1A al vigor híbrido en las líneas maternas LM ACA2000 y LM Cóndor de textura flint, a través del aumento del rendimiento en grano (Figuras 1a y 1b) y del tamaño de la panoja (Figuras 2a y 2b), que permitirían la obtención de híbridos simples de maíz con bajo costo de producción por parte del sector público y privado. Estos resultados concuerdan con otros trabajos donde se detectaron QTLs que contribuyen al fenómeno de heterosis en maíz, encontrándose que a mayor porcentaje de genes marcadores en heterocigosis, el rendimiento en grano era mayor (Stuber et al., 1992; Graham et al., 1997). A su vez, se encontró que después de la F2 de una población de maíz polinizada por cuatro generaciones al azar, sólo 38 de 95 marcadores originales de QTLs ligados para los componentes del grano permanecían significativamente en la población (Dijkhuizen, 1996). Birgham (1998) considera este efecto como dominancia parcial a completa. Si la frecuencia de recombinación entre dos loci es 0,05, luego de tres generaciones de apareamiento al azar, permitiría retener un 86% (1-0,05)³ del ligamiento desequilibrado que estaba presente en la F2 de la población. Si la frecuencia de recombinación es de 0,20, en la misma situación, retendría 51% (1-0,20)³ del ligamiento original. Este autor concuerda con Rhodes et al., (1992) en que se deberá determinar el lugar y la función de cada locus y su contribución a la heterosis, para el dilema "dominancia versus sobredominancia". Por otro lado, la importancia de efectos dominantes fue recientemente confirmada por una alta correlación significativa entre el nivel de heterocigotas y la estabilidad fenotípica, especialmente para rendimiento del grano. Algunas regiones cromosómicas presentaron QTLs sobrepuestos de sobredominancia para peso de plántula, altura de planta, número de granos por planta y rendimiento, sugiriendo efectos pleiotrópicos sobre el vigor de las plantas (Frascaroli et al., 2007).

Figura 1a. Espigas de las líneas progenitoras LM ACA2000 (Foto: LE ACA2000), LP BLS14, la F1 y las retrocruzas R1, R2 y R3.
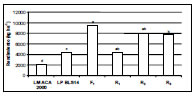
Figura 1b. Rendimiento en grano (kg ha-¹) de las líneas progenitoras LM ACA2000, LP BLS14, la F1 y de las retrocruzas R1, R2. y R3. (LM= Línea materna; LP= Línea paterna). Letras distintas indican diferencias significativas al 5% con el test de Tukey.

Figura 2a. Panojas de las líneas progenitoras LM ACA2000 (Foto: LE ACA2000) y LP BLS14, la F1 y las retrocruzas R1, R2 y R3.
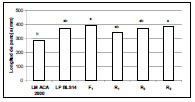
Figura 2b. Longitud de la panoja (mm) de las líneas progenitoras LM ACA2000 y LPBLS14, la F1 y las retrocruzas R1, R2 y R3. Letras distintas indican diferencias significativas al 5% con el test de Tukey.
Estos resultados ponen de manifiesto la importancia de la búsqueda de regiones heteróticas en el genoma de las plantas. Cada región encontrada podría aportar un valor diferencial a la variable que se busca mejorar, ya sea rendimiento en grano, calidad nutricional o resistencia a enfermedades y plagas. En este sentido, nuestro grupo de trabajo detectó marcadores moleculares ligados a QTL para tolerancia a la enfermedad virósica denominada "Mal de Río Cuarto" en Argentina, encontrando diferentes segmentos cromosómicos que fueron mapeados en tres poblaciones los cuales afectan los síntomas de la enfermedad y su tolerancia (Kreff et al., 2006). Esto permitirá a futuro registrar como propiedad intelectual las regiones heteróticas encontradas.
Por último se debe destacar que la metodología de incorporación de segmentos heteróticos emplea conocimientos tradicionales de mejoramiento, permitiendo obtener resultados con menos recursos económicos que los que requieren otras técnicas. De esta manera, la localización de segmentos que confieren valores superiores es un campo de acción enorme en el aumento de la ganancia genética, con un enfoque original a los conceptos tradicionales del vigor híbrido.
BIBLIOGRAFÍA
1. Allard, R. W. & A. D. Bradshaw, (1964). Implications of genotype-environment interactions in applied plant breeding. Crop Science 4:503-508. [ Links ]
2. Ajmone-Marsan, P., Monfredini, G., Ludwig, W.F., Melchinger, A.E., Franceschini, P., Pagnotto, G. & Motto, M.. (1995). In an elite cross of maize a major quantitative trait locus controls one-fourth of the genetic variation for grain yield. Theor Appl Genet 90-3-4:415-424. [ Links ]
3. Apirion, D. & D. Zohary, (1961). Chlorophyll lethal in natural populations of the orchard grass (Dactylis glomerata L.). Genetics 46: 393-399. [ Links ]
4. Band H.T. & P.T. Ives. (1961). Correlated changes in environment and lethal frequency in a natural population of Drosophila melanogaster. Proc. Nat. Acad. Sci. U.S.A. 47: 180-185. [ Links ]
5. Band, H. T. (1963). Genetic structure of populations.II. Viabilities and variances of heterozygotes in constant and fluctuating environments. Evolution [ Links ]
6. Allard, R. W. & A. D. Bradshaw, (1964). Implications of genotype-environment interactions in applied plant breeding. Crop Science 4:503-508. [ Links ]
7. Ajmone-Marsan, P., Monfredini, G., Ludwig, W.F., Melchinger, A.E., Franceschini, P., Pagnotto, G. & Motto, M.. (1995). In an elite cross of maize a major quantitative trait locus controls one-fourth of the genetic variation for grain yield. Theor Appl Genet 90-3-4:415-424. [ Links ]
8. Apirion, D. & D. Zohary, (1961). Chlorophyll lethal in natural populations of the orchard grass (Dactylis glomerata L.). Genetics 46: 393-399. [ Links ]
9. Band H.T. & P.T. Ives. (1961). Correlated changes in environment and lethal frequency in a natural population of Drosophila melanogaster. Proc. Nat. Acad. Sci. U.S.A. 47: 180-185. [ Links ]
10. Band, H. T. (1963). Genetic structure of populations.II. Viabilities and variances of heterozygotes in constant and fluctuating environments. Evolution 17: 307-319. [ Links ]
11. Barucha - Reid, A.T. (1960). Elements of the Theory of Markov processes and their Applications. New York. Ed. Mc Graw-Hill. 468 pp. [ Links ]
12. Berbecel, O. & Eftimescu, M. (1973). Effect of agrometeorogical conditions on maize growth and development. (English translation). P. 10-31. Inst. Meteorol Hydrology. Bucharest- Romania. [ Links ]
13. Berke, T. G. & Rocheford, T. R. (1999). Quantitative trait loci for tassel traits in maize. Crop Sci. 39: 1439-1443. [ Links ]
14. Bingham, E.T. (1998). Role of chromosome blocks in heterosis and estimates of dominance and overdominance. In: Lamkey KR, Staub JE (eds). Concepts and breeding of heterosis in crop plants. Crop Sci. Soc. Am. Madison, Wisconsin, USA.: 71-87. [ Links ]
15. Boggio, R., O. Sorarrain, J. C. Salerno & E. A. Favret, (1997). Theoretical Analysis of Lethal Factors in Plant Populations. Mathematical Biosciences 140: 85-99. [ Links ]
16. Bosso, J. A., O. Sorarrain, & E. A. Favret, (1969). Applications of absorbent Markov chains to sib mating populations with selection. Biometrics 22 (1): 17-26. [ Links ]
17. Allard, R. W. & A. D. Bradshaw, (1964). Implications of genotype-environment interactions in applied plant breeding. Crop Science 4:503-508. [ Links ]
18. Ajmone-Marsan, P., Monfredini, G., Ludwig, W.F., Melchinger, A.E., Franceschini, P., Pagnotto, G. & Motto, M.. (1995). In an elite cross of maize a major quantitative trait locus controls one-fourth of the genetic variation for grain yield. Theor Appl Genet 90-3-4:415-424. [ Links ]
19. Apirion, D. & D. Zohary, (1961). Chlorophyll lethal in natural populations of the orchard grass (Dactylis glomerata L.). Genetics 46: 393-399. [ Links ]
20. Band H.T. & P.T. Ives. (1961). Correlated changes in environment and lethal frequency in a natural population of Drosophila melanogaster. Proc. Nat. Acad. Sci. U.S.A. 47: 180-185. [ Links ]
21. Band, H. T. (1963). Genetic structure of populations.II. Viabilities and variances of heterozygotes in constant and fluctuating environments. Evolution 17: 307-319. [ Links ]
22. Barucha - Reid, A.T. (1960). Elements of the Theory of Markov processes and their Applications. New York. Ed. Mc Graw-Hill. 468 pp. [ Links ]
23. Berbecel, O. & Eftimescu, M. (1973). Effect of agrometeorogical conditions on maize growth and development. (English translation). P. 10-31. Inst. Meteorol Hydrology. Bucharest- Romania. [ Links ]
24. Berke, T. G. & Rocheford, T. R. (1999). Quantitative trait loci for tassel traits in maize. Crop Sci. 39: 1439-1443. [ Links ]
25. Bingham, E.T. (1998). Role of chromosome blocks in heterosis and estimates of dominance and overdominance. In: Lamkey KR, Staub JE (eds). Concepts and breeding of heterosis in crop plants. Crop Sci. Soc. Am. Madison, Wisconsin, USA.: 71-87. [ Links ]
26. Boggio, R., O. Sorarrain, J. C. Salerno & E. A. Favret, (1997). Theoretical Analysis of Lethal Factors in Plant Populations. Mathematical Biosciences 140: 85-99. [ Links ]
27. Bosso, J. A., O. Sorarrain, & E. A. Favret, (1969). Applications of absorbent Markov chains to sib mating populations with selection. Biometrics 22 (1): 17-26. [ Links ]
28. Brandolini, A. and A. Brandolini, (2001). Classification of Italian maize (Zea mays L.) germplasm. Plant Genetic Resources Newsletter No.126: 1-11. [ Links ]
29. Burham, Ch. R. (1993). Balanced lethal from chromosome 6. Maize Genetics Cooperation Newsletter No 67: 101. [ Links ]
30. Carson, H. L. (1967). Permanent heterozygosis. Evolutionary Biology V. I.: 168-193. [ Links ]
31. Cockerham, C.C. (1954). An extension of the concept of partitioning hereditary variance for analysis of covariance's among relatives when epitasis is present. Genetics 39: 859-882. [ Links ]
32. Cockerham, D. M. (1963). Estimation of genetic variances. In W.D. Hanson and H.F. Robinson (ends). Statistical Genetics and Plant Breeding. National Academy of Sciences. National Research Council Publication 982. W:D.: 53-54. [ Links ]
33. Comstock, R.E. & H.F. Robinson, (1948). The components of genetic variance in populations of biparental progenies and their use in estimating the average degree of dominance. Biometrics 4: 254-268. [ Links ]
34. Comstock, R.E. & H.F. Robinson, (1952). Estimation of average dominance of genes. In J.W. Gowen (ed.). heterosis. ISCPress. USA.: 494-516. [ Links ]
35. Crow, J. F. (1948). Alternative hypotheses of hybrid vigour. Genetics 33: 477-487. [ Links ]
36. Crow, J. F. (1999). Dominance and overdominance. In: Coors JG, Pandey, S (eds.). Genetics and exploitation of heterosis in crops. Am. Soc. Agron., Crop Sci. Soc. Am., Soil Sci Soc. Am., Inc, Madison. Wisconsin, USA. : 49-58. [ Links ]
37. Crumpacker, D. W. (1967). Genetic loads in maize (Zea mays L.) and other cross-fertilised plants and animals. Evolutionary Biology 1: 396-415. [ Links ]
38. Dijkhuizen, A., D.W. Dudley & T.R. Rocheford. (1996). Marker-QTL linkages estimated using F2 and random -mated generations. Illinois Corn Breeders School 32: 144-157. [ Links ]
39. Duplesis, D. P. & Dijkhuis, F. J. (1967). The inheritance of lag time between pollen shedding and silking on the yield of maize. S. Afr. J. Agric. Sci. 10. 667-674. [ Links ]
40. Duvick, D. (1977). Genetic rates of gain in hybrid maize yields during the past 40 years. Maydica XXII: 187-196. [ Links ]
41. Duvick, D. (1996). Plant breeding, an evolutionary concept. Crop Science 36: 539-548. [ Links ]
42. Duvick, D. (1999). Commercial Strategies for Exploitation of heterosis. The genetics and exploitation of heterosis in crops. ASA-CSSA-CIMMYT. Chapter 27: 295-304. [ Links ]
43. Duvick, D. (1999). Heterosis: feeding people and protecting natural resources. In: Lamkey KR, Staub JE (eds). Concepts and breeding of heterosis in crop plants. Crop Sci. Soc. Am., Madison, Wisconsin, USA. : 19-29. [ Links ]
44. Duvick, D. (2001). Biotechnology in the 1930. The development of hybrid maize. Nature Reviews genetics 2: 69-74. [ Links ]
45. Emerson, R. A., G. W. Beadle & A. C. Fraser. (1935). A summary of linkage studies in maize. Cornell univ. Agric. Exp. Sta. Mem., No. 180. [ Links ]
46. Favret, E. A. & W. Godeck. (1959). Índice de mutación espontánea en cebada y otras gramíneas. Revista de Investigaciones Agrícolas T. XIII No 3. [ Links ]
47. Falconer, D. S. (1981). Introduction to quantitative genetics. 2nd. edit. Longman editor. 340 pp. [ Links ]
48. Favret, E. A. & G. Ryan. (1966). Possible use of permanent heterozygosis in barley breeding. Barley Newsletter 10: 122-123. [ Links ]
49. Fisher, R.A. (1928). The possible modifications of the response of the wild-type to recurrent mutations. Am. Nat 62:115-126. [ Links ]
50. Frascaroli, E., M. A. Canè, P. Landi, G. Pea, L. Gianfranceschi, M. Villa, M. Morgante & M. E. Pè. (2007). Classical genetic and quantitative trait Loci analyses of heterosis in a maize hybrid between two elite inbred lines. Genetics 176: 625-644. [ Links ]
51. Freire-Maia, N. (1963). Carga genética, o preço da evolução. In C, Pavan and A. B. da Cunha (2d.) Genética. Companhia Editora Nacional, São Paulo, Brasil. 560 pp. [ Links ]
52. Gabriel, K.R. (1971). The biplot graphic display of matrices with applications to principal component analysis. Biometrika 58(3): 453-467. [ Links ]
53. Gallais, A. (1990). Théorie de la selection en amélioration des plantes. Masson Laris. 588 pp. [ Links ]
54. Gardner, C.O. & S. A. Eberhart. (1968). Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22: 439-451. [ Links ]
55. Geraldi, I. O., J. B. Miranda Filho, and R. Vencovsky. (1985). Estimates of genetics parameters for tassel characters in maize (Zea mays L.) and breeding perspectives. Maydica. 30: 1-14. [ Links ]
56. Graham, G.I., D.W. Wolff & C. W. Stuber. (1997). Characterization of a yield quantitative trait locus on chromosome five of maize by fine mapping. Crop Sci. 37: 1601-1610. [ Links ]
57. Griffing, B. (1956). Concept of general and specific combining ability in relation to diallel crossing systems. Australian Jour. Biol. Sci. 9:463-493. [ Links ]
58. Gustafsson, A. (1946). The effect of heterozygosis on variability and vigour. Hereditas 32: 263-286. [ Links ]
59. Gustafsson, A. (1947). The advantageous of deleterious mutations. Hereditas 33: 575. (Abst. [ Links ]).
60. Gustafsson, A. (1953). The cooperation of genotypes in barley. Hereditas 39: 1-18. [ Links ]
61. Harthl, D. L. (1980). Principles of population genetics. Sinauer Assoc. Inc. Sunderland M.A. 488 pp. [ Links ]
62. Hallauer, A.R. & J.B. Miranda. (1981). Quantitative genetics in maize breeding. IsuPress. USA. 468 pp. [ Links ]
63. Infostat . (2004). Infostat/p, versión 2004. Manual del Usuario. Grupo Infostat, FCA, U.N. Córdoba. Primera Edición, Editorial Brujas, Argentina. [ Links ]
64. Jones, D. F. (1945). Heterosis resulting from degenerative changes. Genetics, 30: 527-542. [ Links ]
65. Jones, D. F. (1952). Plasmagenes and chromogenes in heterosis. In J. W. Gowen (ed.), Ames, Iowa, Iowa State College Press. Heterosis. pp. 224-225. [ Links ]
66. Kacser, H., & J. A. Burns. (1981). The molecular basic of dominance. Genetics 97: 639-666. [ Links ]
67. Kempthorne, O. (1954). The correlation between relatives in a random mating population. Proc. Roy. Soc. London, B. 143: 103-113. [ Links ]
68. Kiesselback, T.A. (1951). A half-century of corn research. Am. Sci. 39:629-655. [ Links ]
69. Kreff, E. D. ; M.G. Pacheco, D.G. Díaz, C.G. Robredo, D. Puecher, A. Céliz & J.C. Salerno. (2006). Resistance to Mal de Río Cuarto Virus in Miaze: A QTL Analysis. Journal of Basic and Applied Genetics (BAG), Vol.XVII, N1, Pág. 41-50. [ Links ]
70. Lindstrom, E. W. (1920). Chlorophyll factors in maize. Their distribution on the chromosomes and relation to the problem of inbreeding. J. Heredity 11: 269-277. [ Links ]
71. Lu, H., J. Romero-Severson & R. Bernardo. (2002). Chromosomal regions associated with segregation distortion in maize. Theor. Appl. Genet. 105:622-628. [ Links ]
72. Lu, H., J. Romero-Severson & R. Bernardo. (2003). Theory basic of heterosis explored by simple sequence repeat markers in a randommated maize population. Theor. Appl. Genet. 107:494-502. [ Links ]
73. Luna, J.T. & J. Safont Lis. (1978). El maíz en la Argentina.Vulnerabilidad y recursos genéticos. Ciencia e investigación, Tomo 34, No. 3-4-5-6: 83-90. [ Links ]
74. Mangelsdorf, A. J. (1952). Gene interaction in heterosis. In: Gowen, J.W. (ed.). Heterosis: Iowa State college Press, Ames, Iowa, USA. pp:321-329. [ Links ]
75. Mather, K. and J.L. Jinks. (1971). Biometrical genetics, 2nd. ed. Champman and Hall, London. 382 pp. [ Links ]
76. Mickelson, S. M., Stuber, C. S., Senior, L. and Kaeppler, S. M. (2002). Quantitative trait loci controlling leaf and tassel trait in a B73 x Mo17 population of maize. Crp Sci. 42: 1902-1909. [ Links ]
77. Mihaljevic, R., H. Friedrich Utz & E. Albrecht Melchinger. (2005). No Evidence for Epistasis in Hybrid and Per Se Performance of Elite European Flint Maize Inbreeds from Generation Means and QTL Analyses. Crop Sci 45:2605-2613 [ Links ]
78. Mock, J. J., and S.H. Schuetz. (1974). Inheritance of tassel branch number in maize. Crop Sci. 14: 885-888. [ Links ]
79. Muller, H.J. (1950). Our load of mutations. Am.J.Hum.Gen. 2: 111-176 [ Links ]
80. Otegui, M.E. (1995). Prolificacy and grain yield components in modern Argentinean maize hybrids. Maydica 40: 371-376. [ Links ]
81. Otegui, M.E. & F.H. Andrade. (2000). New relationships between light interception, ear growth and kernel set in maize. In: M.E. Westgate and K. Boote (eds). Physiology and modelling of kernel set in maize. CSSA and ASA Special Publication Nº 29. Baltimore, Maryland, USA. pp 89-102. [ Links ]
82. Redei, G. P. (1962). Single locus heterosis. Z. Vererbungsl 93: 164-170. [ Links ]
83. Rhodes, D., C. Jug, W.J. Yang & Y. Samaras. (1992). Plant metabolism and heterosis. Plant Breed Rev., 10: 53-91. [ Links ]
84. Riman, L. (1963). A synoptic survey of maize genes. Maydica 8: 99-123. [ Links ]
85. Russel, W.A. (1991). Genetic improvement of maize yields. Adv. Agron., 33:245-298. [ Links ]
86. Salerno, J. C. (1981). Utilización de los sistemas letales balanceados en maíz. Acta Jornadas de genética Aplicada del Noroeste Argentino. SAG: 43-51. [ Links ]
87. Salerno, J. C. & E. A. Favret. (1984). Introduction among lethal genes in two lines of maize (Zea mays L.) Genetics 107 (1). 93. (abst. [ Links ]).
88. Salerno, J. C. (1989) Aprovechamiento de los factores letales en el mejoramiento genético. Bol. Genét. 15: 67-72. [ Links ]
89. Salerno, J.C. & D. G. Díaz. (1992). Un Sistema Letal Balanceado en el Cromosoma 6 de Maíz. Acta resumen XXIII Congreso Argentino de Genética, Pergamino, SAG: pp. 35. [ Links ]
90. Salerno, J.C. & Favret E. A. (1994).17 Años de letales balanceados en maíz. Mendeliana. Vol. XI No.1: 82-85. [ Links ]
91. Salerno J. C., D. G. Díaz, C. Robredo, R. Ríos, A. G. Reid, R. Boggio Ronceros & O. Sorrarain. (1997). Lethal genes associated with grain yield in inbred lines of maize. CIMMYT. Proceeding. The genetics and Exploitation of Heterosis in Crops. ASA-CSSA-CIMMYT. México: pp. 134-135. [ Links ]
92.Salerno, J.C., D. G. Díaz, C. Robredo, R. Boggio & O. Sorarrain. (1998). Explotación de la carga genética en la producción de semilla híbrida en maíz. IAMFE-Simposio Internacional de Experimentación de la Maquinaría Agrícola-Castelar: 256-262. [ Links ]
93. Salerno, J. C., R. Boggio, R. & O. Sorarrain. (1999). Análisis teórico de rendimiento en plantas reguladas por factores letales. Revista de Agricultura. Piracicaba. San Pablo, Brasil. Vol.74-No. 2-Set: 137-156. [ Links ]
94. Salerno, J. C., D. G. Díaz, C. Robredo, R. Boggio & O. Sorarrain. (2000). La Carga genética en el mejoramiento genético del maíz. Actas de la XVIII Reunión Latinoamericana del Maíz. CIMMYT-EMBRAPA. Sete Lagoas, Minas Gerais, Brasil. Pp: 211-218. [ Links ]
95. Salerno, J. C., M. Kandus, R. Boggio, O. Sorarrain, C. Gonzalez & D. Almorza. (2007). Genetics and statistical association between lethal alleles and quantitative yield factors in maize (Zea mays L.). Journal of Basic and Applied Genetics (BAG), Vol. XVIII, No. 1:7-13. [ Links ]
96. Schnell, F.W. (1963). The covariance between relatives in the presence of linkage. In W.D. Hanson and H.F. Robinson (eds). Statistical Genetics and Plant Breeding. National Academy of Sci. NRC. Publicación 982. W.D.C. pp: 468-483. [ Links ]
97. Schuetz, S. H., & J. J. Mock, (1978). Genetics of tassel branch number in maize and its implications for a selection program for small tassel size. Theor. Appl. Gene. 53: 265-271. [ Links ]
98. Smith, J. & O. Smith. (1989). The description and assessment of distance between inbred lines of maize: I. The use of morphological traits and descriptors. Maydica 34 (2): 141-150. [ Links ]
99. Sorarrain, O. M. , R. Boggio, & J. C. Ocampo. (1980). Continuous and discontinuous Markov chains for mastitis infection. Math. Biosci. (52): 277-287. [ Links ]
100. Sorarrain, O. M., R. Boggio, J. L. Pousa & E. A. Favret. (1983). Application of absorbent Markov Chains for a Selfing Model of two Independent Loci. J. Theor. Biol. (1983) 103: 173-180. [ Links ]
101. Stuber, C. W., S. E. Lincoln, D. W. Wolff, T. Helentjaris & E. S. Lander. (1992). Identification of genetic factors contributing to Heterosis in a hybrid from two elite maize inbred lines using molecular markers. Genetics 132: 823-839. [ Links ]
102. Stuber, C. W. (1994). Heterosis in plant breeding. Plant Breeding Reviews, No 12: 227-251. [ Links ]
103. Stuber, C. W. (1995). Mapping and manipulating quantitative traits in maize. Trends Genet. 11: 477-481. [ Links ]
104. Vencovsky, R. & P. Barriga. (1992). Genética biométrica no fitomelhoramento. Ribeirão Preto: SBG. 496 P p. [ Links ]
105. Wallace, B. (1970). Genetic load. Its Biological and Conceptual Aspects. Prentice-Hall, Inc. Englewood Cliffs, N.J. Chapter 8: 52 pp. [ Links ]
106. Weijer, J. 1952. A catalogue of genetic maize types together with a maize bibliography. Bibl. Genetics 50: 294. (abst. [ Links ]).
107. Wentz, J. B., & D. F. Goodsell. (1929). Recessive defects and yield in corn. J. Agric. Res. 38:0505-510. [ Links ]
108. Whaley, W. (1964). Physiology of gene action in hybrids. Heterosis. Gowen ed. Hafner, N.Y.: pp. 98-115. [ Links ]
109. Wright, S. (1968, 1969, 1977, 1978). Evolution and the Genetics of Populations.-Vol. 1,2,3,4. University of Chicago Press. 469 pp., 512 pp., 613 pp., 580 pp. [ Links ]
Received: 20/05/2010
Accepted: 17/07/2010

