










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkBAG. Journal of basic and applied genetics
On-line version ISSN 1852-6233
BAG, J. basic appl. genet. vol.22 no.1 Ciudad Autónoma de Buenos Aires Jan./June 2011
Antecedentes y nuevos aportes en el estudio del Cromosoma Y en poblaciones humanas sudamericanas
Graciela Bailliet1, Virginia Ramallo1, Marina Muzzio1, 2, María R Santos1, 2, Josefina MB Motti1, 2, Néstor O Bianchi1, Claudio M1, 2 Bravi
1 Instituto Multidisciplinario de Biología Celular (IMBICE). CCT-CONICET-La Plata; CICPBA. Calle 526 e/ 10 y 11. CC 403. 1900 La Plata, Argentina. Tel: 54 221 4210112. Fax 54 221 4253320.
2 Facultad de Ciencias Naturales y Museo. Universidad Nacional de La Plata. Calle 60 y 122 La Plata (C. P. 1900) (0221) 423-6773/ 425-8252/ 423-2734 gbailliet@imbice. org. ar
RESUMEN
Los avances en el conocimiento del genoma humano han llegado también a una situación comprensiva en relación a la región específica del cromosoma Y humano. En los últimos años se han reconocido al menos 600 polimorfismos bialélicos (SNP), a partir de los cuales se ha construido una sólida filogenia y se ha normalizado la nomenclatura. En el presente trabajo se han estudiado 940 muestras de varones no relacionados de 10 poblaciones aborígenes de Argentina, Chile y Paraguay; y 12 poblaciones urbanas de Argentina. Se han analizado 9 marcadores bialélicos para reconocer los clados mayores de la filogenia. Fue posible distinguir los haplogrupos Q* y Q3 autóctonos, y los haplogrupos AB, DE, F, K, P y R alóctonos. El Fst fue de 0, 17, mostrando un alto nivel de diferenciación entre poblaciones. Las poblaciones aborígenes pudieron ser distinguidas de las urbanas en el análisis de MDS cuyo stress de 0, 0297 evidenció su buen ajuste. Las poblaciones aborígenes autoidentificadas y las poblaciones jujeñas analizadas mantuvieron una proporción importante de cromosomas propios de América, portadores de los haplogrupos Q3, en el caso de las poblaciones de Paraguay la elevada frecuencia de Q*, las distinguió de los demás grupos. Las poblaciones urbanas se distinguieron de las aborígenes por la predominancia de los haplogrupos alóctonos provenientes de varones inmigrantes europeos o de otras regiones geográficas.
Palabras clave: Linajes paternos; Haplogrupos; SNP; Ancestralidad genética; Poblaciones humanas.
ABSTRACT
Advances in the knowledge of the human genome have also impacted in the comprehension of the specific region of the human Y-chromosome. In the last years, almost 600 bialelic polymorphisms (SNP) have been described, allowing a consistent phylogeny to be constructed based on them, and the normalization of nomenclature. In the present work we have analyzed 940 samples from males of 10 aboriginal populations from Argentina, Chile and Paraguay; and 12 urban populations from Argentina. Nine SNP that characterize mayor clades of the phylogeny have been analyzed, enabling the recognition of the Q* and Q3 Native American haplogroups, and AB, DE, F, K, P y R foreigner haplogroups. Fst was 0. 17, showing a high level of differentiation between populations. The MDS analysis showed a differentiation between aboriginal populations and urban, the stress was 0. 0297 meaning a good adjustment. The self-identifed aboriginals and Jujuy populations showed a high proportion of Y-chromosomes from Q3 haplogroups, the Paraguayan populations differentiated from the others due to their high proportion of Q* haplogroups. The urban populations were characterized by the foreign haplogroups entering in those populations by migrations from Europe and other regions.
Key words: Parental lineages; Haplogroups; SNP; Genetic ancestrally; Human populations.
INTRODUCCIÓN
La búsqueda de marcadores polimórficos en la región específica del cromosoma Y, se inició a mediados en los 90's, ante la necesidad de confrontar la información aportada por el ADN mitocondrial para los linajes matrilineales. El Cromosoma Y, tiene una tasa mutacional intermedia entre la de los autosomas y el cromosoma X, su herencia es patrilineal exclusiva y su bajo número específico, ¼ en relación a los autosomas y 1/3 en relación al cromosoma X, lo hace susceptible a sufrir en forma particularmente importante a la deriva genética (Jobling y Smith, 2003). Los primeros marcadores genéticos que pudieron ser estudiados por las técnicas de PCR fueron un microsatélite tetranucleotideo (DYS19, Roewer y col., 1992), un inserto Alu (YAP+, M1, Hammer y col., 1994), un polimorfismo de base única (SNP, "single nucleotide polimorphism") (M2) asociado a cromosomas YAP* Alu (Seielstad y col 1994) y variantes del sistema alfoide (Santos y col., 1995). Desde entonces la búsqueda de polimorfismos se intensificó, y Underhill y colaboradores publicaron la primera filogenia compilando 166 SNP y su correlación con la distribución geográfica continental (Underhill, et al, 2000). Posteriormente se normalizó la nomenclatura y de esta manera pudieron ser comparables los trabajos entre laboratorios (Y Chromosome Consortium 2002). Todos estos antecedentes demostraron que, debido a sus características específicas, el Cromosoma Y, era susceptible de mostrar diferenciación continental y ser sumamente lábil a la deriva genética. De manera de poder distinguirse a los linajes africanos, europeos, asiáticos y americanos.
Para América los primeros antecedentes provinieron de la caracterización del sistema alfoide y el microsatélite DYS 19. En estos estudios tempranos se evidenció que gran parte de los cromosomas Y de poblaciones nativas americanas eran portadores de las variantes II A, para los citados loci (Pena y col, 1995; Santos et al., 1996 a y b). En 1996, Underhill y col. describieron el SNP DYS199 (actualmente denominada M3), se comportaba de forma similar a la descrita en el trabajo de Pena et al.(1995). En un trabajo de nuestro grupo se correlacionaron los 3 sistemas, y se describió la frecuencia de 6 sitios polimórficos, entre los cuales se contaba un nuevo polimorfismo (actualmente denominado SRY2627) (Bianchi et al., 1997). Estos trabajos demostraron que el poblamiento de América sufrió un fuerte efecto fundador en relación a los linajes paternos, por el cual la mayoría de los cromosomas Y pertenecieron a un único linaje. Mediante el estudio de microsatélites específicos del cromosoma Y se definieron los haplotipos, uno de estos fue identificado como ancestral para la diversificación de lo cromosomas Y portadores de M3. Mediante métodos de máxima parsimonia, distintos autores han calculado el tiempo de divergencia de los linajes, a saber: 22. 770 años (13, 500 - 58. 700) (Bianchi et al.1998); 11. 456 (9. 423-13. 797) (Ruiz-Linares y col. 1999); 7, 570 años (SE 681) (Bortolini et al., 2003). Las diferencias temporales en los tres trabajos dependieron fundamentalmente del ajuste del cálculo de las tasas mutacionales y en la edad generacional considerada, Bianchi y col. y Ruiz Linares y col. consideran 27 años y Bortolini y col. 25 años.
A pesar de que los cromosomas portadores de M3, son mayoritarios en las poblaciones americanas, se han hecho denodados esfuerzos para determinar si existieron otros haplogrupos que hayan entrado al continente americano provenientes de Asia. En 1999, Bergen y col. describieron el haplogrupo C, caracterizado por un polimorfismo en el gen RPS4Y, que solo ha sido encontrado, hasta el momento, en América del Norte. En 2003, Seielstad y col. definieron al haplogrupo Q a través de un polimorfismo en el locus M242, todos los cromosomas M3 muestran el alelo derivado para M242, y en este caso estamos en presencia del haplogrupo Q3 (Y Chromosome Consortium 2002). Pero en algunas oportunidades Q tiene el alelo ancestral para M3, y en este caso se denomina paragrupo Q*. El paragrupo Q* fue descrito como otro linaje fundador para América (Seielstad et al., 2003), en Norte América es el segundo haplogrupo en frecuencia cuando no está presente C (Zegura et al., 2004; Bolnick y col., 2006); en Sudamérica tiende a tener frecuencias menores (Bortolini et al., 2006, Bailliet et al., 2009). Q ha acumulado 17 polimorfismos a lo largo de su historia (Karafet et al., 2009), pero todo parece indicar que los linajes americanos son muy cercanos a los linajes Q3, compartiendo al menos 3 de los polimorfismos (Karafet et al., 2008; Bailliet et al., 2009)
Las evidencias presentes hacen suponer que la divergencia entre Q* y Q3 (Y Chromosome Consortium 2002; Karafet et al., 2008) se produjo durante o poco antes del cruce del estrecho de Behring en un cromosoma Q*, en el momento de colonizar el continente o en áreas cercanas al estrecho de Behring (Lell et al., 2002). En tanto el origen de Q* pudo haber precedido la expansión desde Asia al resto del mundo (Bortolini et al., 2003; Seielstad et al., 2003).
En las poblaciones americanas también es posible observar haplogrupos alóctonos. Los haplo-grupos más frecuentes son R y F y en menor frecuencia los haplogrupos DE, K y P (Altuna et al., 2009; Bravi et al., 2000; 1997 a y b; Ramallo et al., 2009 a y b).
En el presente trabajo se caracterizarán poblaciones autodenominadas autóctonas y poblaciones urbanas de Argentina y algunos países limítrofes.
MATERIALES Y MÉTODOS
Se analizaron 940 muestras de varones tomadas al azar y provenientes de donantes voluntarios de 10 poblaciones autodenominadas aborígenes: Ayoreo (9) y Lengua (29) de Paraguay; Huilliche (26) y Pehuenche (18) de Chile; Wichi (118), Toba (5), Chorote (9), Mocoví (27), Mapuche (26), Tehuelche (20). Doce poblaciones urbanas de Argentina: Susques (19), Cochinoca (6), Rinconada (6), San Salvador de Jujuy (87), Catamarca (112), Salta (84), Tucumán (17), La Rioja (87), Mendoza (75), Tartagal (108), Aguaray (13), Azampay (44).
Los SNP (Single Nucleotide Polimorfisms) y una inserción de una región Alu, definen haplo-grupos. Con éstos se ha construido una filogenia confiable que reúne a todos los linajes paternos del mundo (Y Chromosome Consortium, 2002; Karafet et al., 2008). Los haplogrupos tienen una distribución geográfica conocida, lo que permite asignar un posible origen geográfico a los linajes que se encontraron en este estudio. Se utilizaron 9 marcadores: YAP (Hammer, 1994), M168 (Underhill et al.2000), M89 (Underhill et al.2000), M9 (Underhill et al.2000), P27 (Karafet et al.1999), M207 (Su et al.1999), M173 (Su et al.1999), M242 (Seielstad et al, 2003), M3 (Underhill et al.1996).
Se calculó el Indice de Diferenciación Fst (Excoffer et al., 1992) a través del programa Arlequín (Schneider, Roessli, Excoffer 2000). El análisis de MDS se realizó a través del programa NTSYS 2. 11S a partir de la Distancia Da (Nei, 1972) (Exeter Software).
RESULTADOS
Linajes autóctonos:
El haplogrupo Q3 fue encontrado en frecuencias > 80% en las poblaciones Wichi, Toba, Chorote, Pehuenche y Susques. En frecuencias <10% en las poblaciones urbanas de Catamarca, La Rioja y Mendoza. Se encontraron frecuencias intermedias en el resto de las poblaciones (Tabla I).
Tabla 1. Frecuencia de haplogrupos en las poblaciones estudiadas.
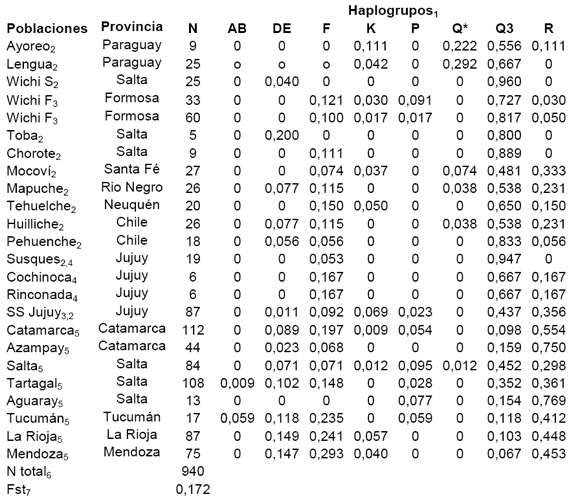
(N) tamaño de la muestra. 1)Y Chromosome Consortium, 2002. 2) Bianchi y col., 1997, 1998. 3) Ramallo y col. 2009b. 4) Dipierri y col., 1998. 5) Ramallo y col., 2009a. 6) N total: número de individuos analizados. 7) Fst (Excoffer y col., 1992).
El paragrupo Q* sólo fue descrito en 6 de las 22 poblaciones analizadas, las máximas frecuencias se encontraron en Lengua y Ayoreo (>22%) de Paraguay. En Mocovies, Huilliches, Mapuches y Salta en frecuencias < 10% (Tabla I) (Bailliet et al., 2009).
Linajes alóctonos:
Los haplogrupos con mayor representación son R y F. R fue importante en las poblaciones urbanas, llegando a mostrar frecuencias mayores al 70% en Aguaray y Azampay. En las poblaciones nativas las frecuencias son menores del 30% e intermedias en el resto de las poblaciones (Tabla I). F estuvo en frecuencias < 30% en Mendoza, La Rioja y Tucumán, en tanto que en las poblaciones aborígenes, en ocasiones, alcanza frecuencias relativas mayores a R, como en Wichi de Formosa, Chorotes y Huilliches de Chile (Tabla I).
Los haplogrupos DE estuvieron en la frecuencia máxima de 14% en Mendoza y La Rioja, en todas las demás poblaciones no superaron el 10%. Los haplogrupos K y P presentaron frecuencias menores al 10% (Tabla I).
Sólo se encontraron 2 cromosomas Y pertenecientes a los haplogrupos AB, en Aguaray y Tucumán (Tabla I).
El 17, 23 % de la variación ocurrió entre poblaciones, el coeficiente de diferenciación entre poblaciones (Fst= 0, 17), fue elevado.
La figura 1 es una representación bidimensional de la matriz de distancias Da (Nei 1972). El stress fue de 0, 0297 que representó un buen ajuste. En la figura se destaca que el primer eje (R1) explicó el 100% de la variación, y separó a todas las poblaciones aborígenes de las poblaciones urbanas, con excepción de las poblaciones jujeñas de SS Jujuy, Rinconada, Cochinoca y Susques; esta porción del gráfico está determinada por las frecuencias elevadas de los haplogrupos Q3, Q* y K. Las 3 muestras de Wichis, Susques, Chorotes, Pehuenches y Rinconada y Cochinoca (estas dos últimas fueron indistinguibles) representan un grupo, el otro grupo lo integran los Mapuches, Tehuelches, Huilliches, Mocovíes y SS de Jujuy. Las muestras Lengua y Ayoreo se posicionaron alejadas en relación a las muestras precedentes, probablemente infuenciadas por presentar la mayor proporción del paragrupo Q*. La otra mitad del gráfico está infuenciada por los haplogrupos foráneos AB, DE, F, P y R son los determinantes de las posiciones de las poblaciones urbanas y semiurbanas a excepción de las ya mencionadas localidades jujeñas. Las muestras Salta, Aguaray y Azampay conforman un grupo; La Rioja y Mendoza conforman otro grupo; finalmente Tartagal, Catamarca y Tucumán se posicionaron sin configurar ningún grupo (Fig. 1).

Figura 1. Escalamiento bidimensional de la matriz de distancia de Nei 1972. O Poblaciones. ¨Haplogrupos; Stress=0. 02970
DISCUSIÓN
En la actualidad, las poblaciones autoidentificadas aborígenes siguen manteniendo un número importante de linajes paternos aborígenes. Aunque lo mismo cabe destacar en contextos urbanos mezclados, las poblaciones jujeñas se distinguieron del resto por su alta contribución amerindia. Deriva de las características etnográficas y la historia de Jujuy por ser una de las áreas más densamente pobladas en épocas precolombinas y opuso una fuerte resistencia a la colonización (Hernández, 1992; Pucci, 1998). Por otro lado, existen evidencias de una menor proporción de mezcla genética con la altura, en cierta manera la altitud actuó como una barrera o un efecto disuasorio para los habitantes de origen europeo (Dipierri et al., 1997, 1998, 2000).
En cuanto a la relación entre las poblaciones Mapuches, Huilliches y Tehuelches, también es posible interpretar este resultado en un contexto histórico. Los Mapuches de Argentina y los Huilliches de Chile tienen el mismo origen y el contacto entre Mapuches y Tehuelches ha sido bien documentado (Martínez Sarasola, 1992).
Linajes autóctonos para América:
Q3 (portador de M3) es el mayoritario y el más ampliamente distribuido en América (Underhill et al, 1996; Bianchi et al, 1998, Bortolini et al., 2003) y es considerado autóctono para América.
Q* (portadores de M-242, exclusivamente) filogenéticamente ancestral al haplogrupo mayoritario Q3 (Seielstad et al., 2003, Bortolini et al, 2003). Tiene una frecuencia muy baja en Sudamérica (< 6%, salvo algunas excepciones) (Bailliet et al.2009). A través del trabajo de Karafet, et al. (2008), pudo evidenciarse que el clado Q* ha acumulado 17 mutaciones, pero hasta el momento todos los haplogrupos Q* de la presente muestra presentan el estado derivado para M346 C-G (Sengupta et al., 2006; Bailliet y col., 2009). Según los resultados de Karafet, et al.(2008), es muy probable que todos estos cromosomas también tengan los estados derivados de P36. 2 C-T y MEH2 G-T (Y Chromosome Consortium, 2002).
C (portador de RPS4Y) filogenéticamente muy lejano a los 2 precedentes (Bergen y col., 1999). No ha sido encontrado en América del Sur (Bortolini et al., 2003), salvo por alguna excepción en los Wayuu (Zegura et al.2004).
Q* y C, se encuentran en otras regiones geográficas, principalmente en Asia (Seielstad et al., 2003; Zerjal et al., 2002).
Linajes alóctonos:
A-B: son casi exclusivos de Africa Sub-Sahariana, los cromosomas A son muy frecuentes entre los Khoisán, en Etiopía y Sudan, mientras que B fue encontrado en su frecuencia máxima en Pigmeos (Hammer et al.2001; Underhill et al.2001; Jobling y Tyler-Smith 2003). Esta es la primera vez que AB se describe en Argentina, la presencia africana ha sido inicialmente reconocida a través de los cromosomas Yap+ (Bravi et al., 2000).
F: Es el 3º haplogrupo en frecuencia en las poblaciones de Argentina estudiadas hasta el momento en nuestro laboratorio. Es propio de Europa y de Medio Oriente (Jobling and Tyler-Smith 2003; Sengupta y col. 2006, Hammer y col. 2000, 2001; Underhill et al.2000).
K: es un haplogrupo minoritario en las muestras de Sudamérica. Comprende a los subhaplogrupos de origen asiático (Su et al.2000; Hammer et al.2001; Su et al.1999; Underhill et al.2001).
R: es el haplogrupo más frecuente en Europa (Jobling y Tyler Smith 2003). En Argentina es el haplogrupo más abundante en las poblaciones urbanas (Ramallo y col., 2009a). En América, gran parte de los linajes alóctonos se referen al Clado R, específicamente R1b, estos linajes pueden explicarse por la fuerte migración europea del último siglo, especialmente el ingreso de españoles e italianos. Algo similar se observa en otros países sudamericanos como Brasil (Bortolini y col., 2003), Méjico (Rangel-Villalobos et al., 2008) y en USA (Zegura et al., 2004; Bolnick et al., 2006). En el subhaplo-grupo R1b el polimorfismo SRY 2627 define un subclado, en muestras urbanas de la ciudad de La Plata fue descrito en el 11% (Bianchi et al., 2007).
AGRADECIMIENTOS
Agradecemos profundamente a todos los donantes voluntarios que hicieron posible este trabajo de investigación. Ofrecemos nuestro reconocimiento por los aportes críticos de Susana Salceda, José E. Dipierri, Emma L. Alfaro. El proyecto fue financiado por CONICET, FONCyT, CICPBA y Fundación Antorchas.
BIBLIOGRAFÍA
1. Altuna, M. E., García, A., Ramallo, V., Bailliet, G., Modesti, N. M., Demarchi, D. A. (2009). Origin of paternal lineages in an admixed population of Northern Argentina (La Esperanza, Jujuy). Forensic Science International: Genetics Supplement Series 2:451- 452. [ Links ]
2. Bailliet, G., Ramallo, V., Muzzio, M., García, A., Santos, M. R., Alfaro, E. L., Dipierri, J. E., Salceda, S., Carnese, F. R., Bravi, C. M. Bianchi, N. O., Demarchi, D. A. (2009). Brief Communication: Restricted Geographic Distribution for Y-Q* Paragroup in South America. Am. J Phys Anthropol. 140:578-582 (2009) [ Links ]
3. Bergen, A. W., Wang, C. Y., Tsai, J., Jefferson, K., Dey, C., Smith, K. D., Park, S. C., Tsai, S. J., Goldman, D. (1999). An Asian-Native American paternal lineage identifed by RPS4Y resequencing and by microsatellite haplotyping. Ann Hum Genet 63:63-80 [ Links ]
4. Bianchi, N., Bailliet, G., Bravi, C. M., Pena SD, Rothhammer F. (1997). Origin of Amerindian Y-chromosome as inferred by the analysis of six polymorphic markers. Am J Phys Anthropol 102:79-89. [ Links ]
5. Bianchi, N. O., Catanesi, C. I., Bailliet, G., Martinez-Marignac, V. L., Bravi, C. M., Vidal-Rioja, L. B., Herrera, R. J., Lopez-Camelo, J. S. (1998). Characterization of ancestral and derived Y-chromosome haplotypes of New World native populations. Am. J Hum Genet 63:1862-1871. [ Links ]
6. Bolnick, D. A., Bolnick, D. I., Smith, D. G. (2006). Asymmetric male and female genetic histories among Native Americans from Eastern North America. Mol. Biol. Evol. 23(11): 2161-2174 [ Links ]
7. Bortolini, MC., Salzano, F. M., Thomas, M. G., Stuart, S., Nasanen, S. P. K., Bau, C. H. D., Hutz, M. H., Layrisse, Z., Petzl-Erler, M. L., Tsuneto, L. T., Hill K., Hurtado, A. M., Castro-de-Guerra, D., Torres, M. M., Groot, H., Michalski, R., Nymadawa, P., Bedoya, G., Bradman, N., Labuda, D., Ruiz-Linares, A. (2003). Y-chromosome evidence for differing ancient demographic histories in the Americas. Am J Hum Genet 73(3):524-39. [ Links ]
8. Bravi, C. M., Bailliet, G., Crisci, J. V., Bianchi, N. O. (1997a). Evolution of YAP+ human Y-chromosomes J. Genet. Mol. Biol. 8: 65-69. b [ Links ]
9. Bravi, C. M., Bailliet, G., Martìnez-Marignac, V. L., Bianchi, N. O. (2000). Origin of YAP+ lineages of the human Y-chromosome. Am. J. Phis. Antrop. 112 (2):149-158. [ Links ]
10. Bravi, C. M., Sans, M., Bailliet, G., Martinez-Marignac, V. L., Bianchi, N. O. (1997b). Characterization of mitochondrial and Y-chromosome haplotypes in a uruguayan population of African ancestry. Hum. Biol. 69 (5):641-652. [ Links ]
11. Dipierri, J. E., Alfaro, E., Martinez-Marignac, V. L., Bailliet, G., Bravi, C. M., Cejas, S., Bianchi, N. O. (1998). Paternal directional mating in two amerindian subpopulations from the northwest of Argentina". Hum. Biol. 70 (6):1001-1010. [ Links ]
12. Dipierri, J. E., Alfaro, E., Peña, J. A., Jacques, C. and Dogoujon, J. M. (2000). GM, KM inmunoglobulin allo-types and other serum genetic markers ( HP, GC, PI and TF) among south American populations living at different altitude (Jujuy province, Argentina) Admixture Estimates. Hum. Biol. 72(2):305-319. [ Links ]
13. Dipierri, J. E., Alfaro, E., Vullo, C., Gutiérrez, N. (1997). El polimorfismo HLA en el noroeste argentino. III Jornadas Nacionales de Antropología Biológica, Rosario Argentina. [ Links ]
14. Excoffer L, G. L, and Schneider S. (2005). Arlequin ver. 3. 0: An integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online 1. [ Links ]
15. Schneider, S., Roessli, D., Excoffer, L. (2000). Arlequin ver. 2. 000: A software for population genetics data analysis. Genetics and Biometry Laboratory. Unversity of Geneva. Switzerland. [ Links ]
16. Hammer, M. F. (1994). Arecent insertion of an Alu element on the Y chromosome is a useful marker for human population studies. Mol. Biol. Evol. 11:749- 761. [ Links ]
17. Hammer, M. F., Blackmer, F., Garrigan, D., Nachman, M. W., Wilder, J. A. (2003). Human population structure and its effects on sampling Y chromosome sequence variation. Genetics 164: 1495-1509 [ Links ]
18. Hammer, M. F., Karafet, T. M., Redd, A. J., Jarjanazi, H., Santachiara-Benerecetti, S., Soodyall, H., Zegura, S. L. (2001). Hierarchical patterns of global human Y-chromosome diversity. Mol. Biol. Evol. 18: 1189- 1203 [ Links ]
19. Hammer, M. F., Redd, A. J., Wood, E. T., Bonner, M. R., Jarjanazi, H., Karafet, T., Santachiara-Benerecetti, S., Oppenheim, A., Jobling, M. A., Jenkins, T., et al. (2000). Jewish and Middle Eastern non-Jewish populations share a common pool of Y-chromosome biallelic haplotypes. Proc. Natl. Acad. Sci. 97: 6769- 6774. [ Links ]
20. Hernández, I. (1992). Los indios de la Argentina. Colecciones Mapfre 1492, Madrid. [ Links ]
21. Jobling, M. A. and Tyler-Smith, C. (2003). The human Y chromosome: An evolutionary marker comes of age. Nat. Rev. Genet. 4: 598-612. [ Links ]
22. Karafet, T. M., Mendez, F. L., Meilerman, M. B., Underhill, P. A., Zegura, S. L., Hammer, M. F. (2008). New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree. Genome Res. 18(5):830-8. [ Links ]
23. Karafet, T. M., Zegura, S. L., Posukh, O., Osipova, L., Bergen, A., Long, J., Goldman, D., Klitz, W., Harihara, S., de Knijff, P., Wiebe, V., Griffths, R. C., Templeton, A. R. y Hammer, M. F. (1999). Ancestral Asian Source(s) of New World Y-Chromosome Founder Haplotypes. Am. J. Hum. Genet. 64:817-831, [ Links ]
24. Lell, J. T., Sukernik, R. I., Starikovskaya, Y. B., Su, B., Jin, L., Schurr, T. G., Underhill, P. A., Wallace, D. C. (2002). The dual origin and Siberian affinities of Native American Y chromosomes. Am J Hum Genet 70:192-206. [ Links ]
25. Martinez Sarasola C . (1992). In: EMECE (ed.): Nuestros paisanos los indios. Vida, historia y destino de las comunidades indígenas de la Argentina. [ Links ]
26. Nei, M. (1972). Genetic distance between populations. Amer. Naturalist, 106:283-292. [ Links ]
27. Pena, S. D. J., Santos, F. R., Bianchi, N. O., Bravi, C. M., Carnese, F. R., Rothhammer, F., Gerelsaikhan, T., Munkhtuja, B., Oyunsuren, T. (1995). A major founder Y-chromosome haplotype in Amerindians. Nat Genet 11:15. [ Links ]
28. Pucci R. (1998). El tamaño de la población aborigen del Tucumán en la época de la conquista: balance de un problema y propuesta de una estimación. Población y Sociedad Nº 5: 239-270. [ Links ]
29. Ramallo, V., Mucci, J. M., García, A., Muzzio, M., Motti, J. M. B., Santos, M. R., Perez, M. E., Alfaro, E. L., Dipierri, J. E., Demarchi, D. A., Bravi, C. M., Bailliet, G.. (2009a). Comparison of Y chromosome haplogroup frequencies in 8 Provinces of Argentina. Forensic Science International: Genetics Supplement Series 2:431-432 [ Links ]
30. Ramallo, V., Muzzio, M., Motti, J. B. M., Salceda, S., Bailliet, G. (2009b). Linajes masculinos y su diversidad en comunidades wichi de Formosa. Revista del Museo de Antropología 2: 67-74 [ Links ]
31. Rangel-Villalobos, H., Muñoz-Valle, J. F., González-Martín, A., Gorostiza, A., Magaña, M. T., Páez-Riberos, L. A. (2008). Genetic admixture, relatedness, and structure patterns among Mexican populations revealed by the Y-chromosome. Am J Phys Anthropol. 135(4):448-61. [ Links ]
32. Roewer, L., Arnemann, J., Spurr, N. K., Grzeschik, K. H. and Epplen, J. T. (1992). Simple repeat sequences on the human Y chromosome are equally polymorphic as their autosomal counterparts. Hum. Genet., 89, 389-394. [ Links ]
33. Ruiz-Linares, A., Ortiz-Barrientos, D., Figueroa, M., Mesa, N., Munera, J. G., Bedoya, G., Velez, I. D., Garcia, L. F., Perez, Lezaun, A., Bertranpetit, J., Feldman, M. W., Goldstein, D. B. (1999). Microsatellites provide evidence for Y chromosome diversity among the founders of the New World. Proc Natl Acad Sci USA 96:6312-6317. [ Links ]
34. Santos, F. R., Pena, S. D. J., and Tyler-Smith, C. (1995b). PCR haplotypes for the human Y chromosome based on alphoid satellite variants and heteroduplex analysis. Gene 165:191-198. [ Links ]
35. Santos, F. R., Bianchi, N. O., and Pena, S. D. J. (1996a). Worldwide distribution of human Y chromosome haplotypes. Genome Res. 6:601-611. [ Links ]
36. Santos, F. R., Rodríguez-Delfin, L., Pena, S. D. J., Moore, J., and Weiss, K. M. (1996b). North and South Amerindians may have the same major founder Y chromosome haplotype. Am. J. Hum. Genet. 58:1369-1370. [ Links ]
37. Seielstad, M., Yuldasheva, N., Singh, N., Underhill, P., Oefiner, P., Shen, P., Wells, R. S. (2003). A novel Y-chromosome variant puts an upper limit on the timing of frst entry into the Americas. Am J Hum Genet 73(3):700-5 [ Links ]
38. Seielstad, M., Herbert, J. M., Lin, A. A., Underhill, P. A., Ibrahim, M., Vollrath, D., and Cavalli-Sforza, L. (1994). Construction of human Y-chromosomal haplotypes using new polymorphic A to G transition. Hum. Mol. Genet. 3:2159-2161. [ Links ]
39. Sengupta, S., Zhivotovsky, L. A., King, R., Mehdi, S. Q., Edmonds, C. A., Chow, C. E., Lin, A. A., Mitra, M., Sil, S. K., Ramesh, A., Usha Rani, M. V., Thakur, C. M., Cavalli-Sforza, L. L., Majumder, P. P., Underhill, P. A. (2006). Polarity and temporality of high-resolution Y- chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic infuence of Central Asian pastoralists. Am J Hum Genet 78:202-221. [ Links ]
40. Su, B., Xiao, J., Underhill, P., Deka, R., Zhang, W., Akey, J., Huang, W., Shen, D., Lu, D., Luo, J., Chu, J., Tan, J., Shen, P., Davis, R., Cavalli-Sforza, L., Chakraborty, R., Xiong, M., Du, R., Oefiner, P., Chen, Z., Jin, L. (1999). Y-chromosome evidence for a northward migration of modern humans into Eastern Asia during the last Ice Age. Am J Hum Genet 65:1718-1724. [ Links ]
41. Su, B., Jin, L., Underhill, P., Martinson, J., Saha, N., McGarvey, S. T., Shriver, M. D., Chu, J., Oefiner, P., Chakraborty, R., et al. (2000). Polynesian origins: Insights from the Y chromosome. Proc. Natl. Acad. Sci. 97: 8225-8228. [ Links ]
42. Underhill, P. A., Jin, L., Zemans, R., Oefiner, P. J. & Cavalli-Sforza, L. L. (1996). A pre-Columbian Y chromosome-specific transition and its implications for human evolutionary history. Proc. Natl Acad. Sci. USA 93, 196-200. [ Links ]
43. Underhill, P. A., Shen, P., Lin, A. A., Jin, L., Passarino, G., Yang, W. H., Kauffman, E., Bonne-Tamir, B., Bertranpetit, J., Francalacci, P., Ibrahim, M., Jenkins, T., Kidd, J. R., Mehdi, S. Q., Seielstad, M. T., Wells, R. S., Piazza, A., Davis, R. W., Feldman, M. W., Cavalli-Sforza, L. L., Oefiner, P. J. (2000). Y chromosome sequence variation and the history of human populations. Nat Genet 26:358-361. [ Links ]
44. Underhill, P. A., Passarino, G., Lin, A. A., Shen, P., Mirazon, L. M., Foley, R. A., Oefiner, P. J., Cavalli-Sforza, L. L. (2001). The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations. Ann Hum Genet 65:43-62. [ Links ]
45. Y Chromosome Consortium. (2002). A nomenclature system for the tree of human Y-chromosomal binary haplogroups. Genome Res 12:339-348. [ Links ]
46. Zegura, S. L., Karafet, T. M., Zhivotovsky, L. A., Hammer, M. F. (2004). High-resolution SNPs and microsatellite haplotypes point to a single, recent entry of Native American Y chromosomes into the Americas. Mol. Biol. Evol. 21: 164-175. [ Links ]
47. Zerjal, T., Wells, R. S., Yuldasheva, N., Ruzibakiev, R., Tyler-Smith, C. (2002). A genetic landscape reshaped by recent events: Ychromosomal insights into central Asia. Am J Hum Genet 71:466-482. [ Links ]
Received: 26/05/2010
Accepted: 29/08/2011

