










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBAG. Journal of basic and applied genetics
versión On-line ISSN 1852-6233
BAG, J. basic appl. genet. vol.22 no.2 Ciudad Autónoma de Buenos Aires dic. 2011
ARTÍCULOS ORIGINALES
Estructura genético-isonímica del noroeste Argentino
José E. Dipierri, Emma L. Alfaro, Graciela Bailliet, Claudio Bravi, Maria E. Albeck, Marina Muzzio, Virginia Ramallo, Josefina María Brenda Motti
Instituto de Biología de la Altura. Universidad Nacional de Jujuy.
Laboratorio de Genética Molecular Poblacional, IMBICE, CCT CONICET La Plata.
Facultad de Humanidades y Ciencias Sociales. Universidad Nacional de Jujuy. CONICET
Avda. Bolivia 1661. (4600) San Salvador de Jujuy
dipierri@inbial.unju.edu.ar
jedipierri49@yahoo.com
ABSTRACT
From a collaborative effort of several institutions and researchers the aim of this work was to further studies on the population structure of Northwest Argentina (NWA) using both the analysis of the distribution and frequency of surnames by the isonymic method and genetic markers. To this end, blood markers (ABO, HLA), Y chromosome, mtDNA and total genomic analysis were used, allelic and haplogroup frequencies were determined, intra-and interpopulation genetic diversity was analyzed and dendrograms were constructed. To assess NWA miscegention, which started in the second half of the XVII century, rates were calculated using as a reference allele frequencies of (European, Amerindian and African) parental populations. Surnames are suitable material for analyzing the structure of populations in terms of consanguinity and kinship and their ethnic origin. This paper analyzes the inbreeding coefficient by random isonymy, isonymic distances calculated as from the identity of surnames and the richness or abundance of surnames that can weigh genetic drift by isonymy. The genetic structure of NWA population shown by isonymy was that it is characterized by low interpopulation differentiation at large urban center level, isolation by distance with a large genetic differentiation among departmental subpopulations, high consanguinity level in isolated departments that are not part of the capital cities in the region, and varying degrees of miscegenation according to the markers considered, but which generally revealed a predominant Amerindian contribution followed by European and African participation. These results are consistent with the demographic attributes, historical background of settlement in the region and geographical particularities of the NWA, which would lead to conclude that the population structure of the NWA would be determined by the migration status and relative isolation of its populations.
Key words: Molecular markers; Population structure; Northwest Argentina.
RESUMEN
El objetivo de este trabajo es profundizar el estudio de la estructura poblacional del NOA recurriendo al análisis de la distribución y frecuencia de apellidos por el método isonímico y a la utilización de marcadores genéticos a partir de un esfuerzo colaborativo de varias instituciones e investigadores. Para ello se utilizaron marcadores sanguíneos (ABO, HLA), cromosoma Y, ADNmt y análisis genómico total y se determinaron frecuencias alélicas y de haplogrupos, se analizó la diversidad genética intra e interpoblacional y se construyeron dendrogramas. Para evaluar el mestizaje en el NOA iniciado en la segunda mitad del siglo XVII se calcularon porcentajes de miscegenación usando como referencia frecuencias alélicas de las poblaciones parentales (europea, amerindia y africana), familias lingüísticas y poblaciones amerindias. Los apellidos constituyen un material adecuado para analizar la estructura de las poblaciones en términos de consanguinidad y parentesco y el origen étnico de las mismas. En este trabajo se analiza el coeficiente de consanguinidad por isonimia al azar, las distancias isonímicas calculadas a partir de la identidad de apellidos y la riqueza o abundancia de apellidos que permite ponderar la deriva génica por isonimia.
La estructura genético-isonímica del NOA se caracterizaría por una escasa diferenciación interpoblacional a nivel de los grandes centros urbanos, aislamiento por distancia, con una gran diferenciación genética entre sus subpoblaciones departamentales, alta consanguinidad en los departamentos más aislados no integrantes de las ciudades capitales de provincia y grados variables de miscegenación de acuerdo a los marcadores considerados pero que en general revelan una contribución amerindia preponderante, seguida por la europea y africana. Estos resultados son coherentes con los atributos demográficos, los antecedentes históricos del poblamiento de la región y las particularidades geográficas del NOA, lo que permitiría concluir que la estructura poblacional del NOA se encontraría determinada por la condición migratoria y el aislamiento relativo de sus poblaciones.
Palabras clave: Apellidos; Marcadores moleculares; Estructura de población; NOA
LAS ESTRUCTURAS DE POBLACIÓN
Los estudios de población convocan la atención de muchas disciplinas (demografía, economía, sociología, genética, etc.) lo que convierte a la población, por estar asociada y existir en marcos espacio-temporales concretos, en un objeto de estudio interdisciplinario. Cada disciplina puede delinear correctamente niveles de población o estructuras dentro de las poblaciones. Éstas pueden ser tan inclusivas como la población mundial o tan limitada como la familia (Macbeth y Collison, 2002). En 1972 Harrison y Boyce describen los patrones de agrupamiento jerárquico observados dentro de las poblaciones subdivididas como estructuras, biológicas y/o culturales. En algunas disciplinas, la configuración de este patrón resulta fácil de delimitar. Demográfica o geográficamente el patrón puede ser desplegado desde el hogar al país, pasando por la ciudad, los departamentos, las provincias o sus equivalentes administrativos. Genéticamente, una población puede ser subdividida en subpoblaciones y éstas en demes; socialmente, desde la familia a la comunidad, al grupo étnico o idiomático, etc.
Sin embargo, el concepto de estructura de población es utilizado de manera imprecisa porque el mismo difiere de acuerdo a cada enfoque disciplinario (Castro de Guerra, 1987). En demografía se refiere a la composición por sexo y edad, junto a la consideración de la frecuencia de nacimientos, muertes y migraciones sucedidas en un período de tiempo determinado. En genética poblacional este concepto tiene varias acepciones. Para Vogel y Motulsky (1982) la estructura genética de una población se refiere a la composición genética de la misma, en términos de la distribución de alelos y genotipos, por los efectos combinados de la mutación, selección, migración y deriva génica. Yasuda y Morton (1967) definen la estructura genética como la totalidad de las desviaciones de la panmixia y la misma resultaría de los efectos de la consanguinidad, migración, apareamiento direccional, deriva génica y tomaría en cuenta además el tamaño y distribución de las poblaciones subdividas (Lasker y Mascie-Taylor, 1993).
Desde la perspectiva disciplinar, el estudio de la estructura poblacional podría parecer en todo momento operativo. Las poblaciones se abordarían desde el punto de vista de cada disciplina, sus métodos y supuestos teóricos. Sin embargo, este enfoque clásico resulta en cierta forma limitante. A menos que el objetivo de estos estudios sea meramente descriptivo, las diferentes estructuras de población no deberían considerarse aisladamente, sino en forma conjunta intentando comprender sus interrelaciones (Castro de Guerra, 1987). La complejidad de la población como objeto de estudio justifica aún más la interdisciplinariedad de su abordaje, la necesidad de un enfoque global que incluya junto a la consideración de los factores disciplinares específicos, otros provenientes de disciplinas relacionadas. Este enfoque sistémico debería contribuir a integrar las múltiples interrelaciones entre los diversos fenómenos (demográfico, social, biológico, económico, etc.) que caracterizan a las poblaciones humanas.
LA REGIÓN NOA
Los conceptos de espacio, territorio y región se encuentran estrechamente interrelacionados; abarcan muchas disciplinas y no han alcanzado aún consenso teórico y metodológico (Ortega Valcarcel, 2000). El término región es utilizado en una variedad de formas para denotar el compartimiento de un espacio de significancia formal, funcional o perceptiva.
Uno de los modelos aplicados al análisis de la dinámica económica del espacio territorial es el de centro-periferia, aplicado en la mayoría de los países en vías de desarrollo. Según este modelo, la transformación económica se organiza sobre la base de una estructura polarizada, en donde las regiones son subsistemas, relacionados entre sí jerárquicamente, al sistema nación (Hilhorst, 1971). A partir de este modelo se deducen dos tipos de regionalización, la polarizada y la homogénea. El modelo de regiones polarizadas fue aplicado a 6 provincias del Noroeste Argentino (NOA) por el CFI (1963) para determinar las regiones económicas de Argentina (Hilhorst, 1971). Los espacios geoeconómicos definidos a partir del concepto de región polarizada están ligados al de región natural o región geográfica, un espacio del territorio diferenciado o delimitado, continuo y homogéneo (Ortega Valcárcel, 2000). La región se comportaría como una unidad estructurada que le otorga singularidad e individualidad (Boisier, 1992).
No existe consenso sobre las provincias que integran el NOA. Mientras algunos autores (Pontussi, 1995) coinciden con la clasificación del CFI (1963), otros (Boleda, 1993; Boleda, 1998; Bolsi y Pucci, 1997) incluyen a La Rioja porque pese a sus diferencias, las seis provincias mantienen una homogeneidad demográfica que justifica su inclusión en un conjunto espacial supraprovincial. Además de las características económicas (CFI, 1963; Hilhorst, 1971) y demográficas (Boleda, 1993; Boleda, 1998; Bolsi y Pucci, 1997), los antecedentes históricos (Bazán, 1986), etnohistóricos (Lorandi y Otonello, 1992) y culturales (Ruiz, 1993) contribuyen también a la singularidad del NOA como región. Biogeográficamente el NOA integra la Región Neotropical y dentro de ésta comparte los Dominios Amazónico (Provincia Yungas), Chaqueño (Provincias Prepuneña y Chaqueña) y Andino-Patagónico (Provincias Altoandina y Puneña) (Cabrera y Willink, 1973).
Las poblaciones nativas del NOA se adscriben a las Áreas Culturales Chaco Austral y Noroeste (macro Área Andina) (Hernández, 1992), esta última la más importante del país y con mayor trascendencia en el desarrollo de la población actual del NOA por su importancia numérica en tiempos prehispánicos y por su permanencia debida a la resistencia que ofrecieron al proceso de desnaturalización iniciado con la conquista (Hernández, 1992; Pucci, 1998). Basándose en el momento de ingreso de las poblaciones alóctonas continentales y extracontinentales se puede dividir el poblamiento del NOA en tres fases: a) precolonial, con una profundidad temporal de al menos 9200±600 BP por datación del sitio de Inca Cueva (Jujuy) (Aschero, 1980); b) colonial, marcada por el ingreso de población europea a partir de 1542 y africana durante el siglo XVIII. El Censo Virreinal de 1778 da cuenta de la composición étnica del NOA en este momento del poblamiento (Tabla I); c) formación del estado nacional caracterizado por el ingreso masivo (1857-1926) de migrantes europeos y de otros continentes que se distribuyen heterogéneamente en el NOA, concentrándose en Tucumán y capitales provinciales.
Tabla I. Composición étnica de la población del noroeste argentino según el censo de Carlos III de 1778.
Fuente: Comadrán Ruiz, 1969
A partir de un esfuerzo colaborativo de varias instituciones e investigadores se ha podido profundizar el análisis de la estructura poblacional del NOA iniciado en los 90 (Dipierri y Alfaro, 1994) recurriendo al análisis de la distribución y frecuencia de apellidos por el método isonímico y a la utilización de marcadores genéticos. Este trabajo se propone revisar los principales resultados obtenidos con estas investigaciones.
LOS GENES DEL NOA
En el caso de los marcadores genéticos los datos son representativos de poblaciones urbanas y las muestras de sangre fueron proporcionadas por donantes de sangre y de órganos en hospitales públicos del NOA. Se utilizaron marcadores sanguíneos (ABO, HLA) (Morales et al., 2000; Alfaro et al., 2004; 2005), ADNmt (Dipierri et al., 1998; Tamm et al., 2007; Bravi, 2005), cromosoma Y (Dipierri et al., 1998; Muzzio et al., 2010; Muzzio, 2010; Bailliet et al., 2005; Ramallo et al., 2005; 2009) y análisis genómico total (678 microsatélites autosómicos y 29 ligados al X) (Wang et al., 2008). Más detalles de la metodología utilizada para la determinación de los mismos puede consultarse en la bibliografía consignada previamente.
Se determinaron frecuencias alélicas y de haplogrupos, se analizó la diversidad genética intra e interpoblacional y se construyeron dendrogramas. Para evaluar el creciente mestizaje en el NOA iniciado en la segunda mitad del siglo XVII se calcularon porcentajes de miscegenación usando como referencia frecuencias alélicas de las poblaciones parentales (europea, amerindia y africana), familias lingüísticas y poblaciones amerindias (Wang et al., 2008).
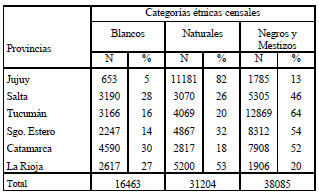
Referente al sistema ABO, estudiado en 95.562 individuos (2.7% población del NOA), el alelo más frecuente en todas las provincias fue el 0. El alelo A presenta una frecuencia superior al 15% en Tucumán, Santiago del Estero, Catamarca y La Rioja, mientras que el B es mayoritario en Tucumán y Santiago del Estero (Tabla II) (Morales et al., 2000). La baja frecuencia continental de A y B en centro y suramerindios apoya la hipótesis de que estos grupos étnicos precolombinos eran portadores solo del alelo 0 y que los alelos A y B podrían haber sido introducidos por las poblaciones alóctonas o resultar de una fuerte selección y/o pérdida por deriva (Dipierri et al., 1999). La tipificación en momias de alelos A sugiere que las poblaciones suramerindias precolombinas portaban este alelo y quizás el B, pero en muy baja frecuencia. Sin embargo, estos hallazgos en momias son muy controvertidos (Salzano y Callegari Jacques, 1988; Llop y Rothhhammer, 1988).
El sistema ABO (Tabla II) indicaría una baja diversidad genética interpoblacional entre las poblaciones urbanas capitalinas del NOA ya que el 99.9% de la variación encontrada resulta de diferencias intrapoblacionales (Morales et al., 2000). Con el sistema HLA puede también observarse una remarcable homogeneidad genética entre las provincias del NOA y una estructura genética similar con un exceso de heterocigosis (Alfaro et al., 2005). En cuanto a la variabilidad alélica del sistema HLA las poblaciones urbanas del NOA, al igual que otras poblaciones amerindias, se caracterizan por un limitado número de alelos, más notorio en el locus A que en el B (Salzano y Callegari Jacques, 1988). En efecto, solo 4 alelos del locus A (A2, A24, A28, A31) se presentan en el 60% del total de individuos analizados en el NOA, y ellos son marcadores típicamente amerindios (Rothhammer et al., 1997) (Tabla II). En el locus B la misma proporción está representada por 6 especificidades, B5, B62, B60, B51, B44 y B7, de las cuales las 4 primeras se encuentran frecuentemente en Amerindios (Alfaro et al, 2004; 2005). La introgresión de genes de población africana ha sido demostrada para la provincia de Jujuy con los haplogrupos del sistema GM, cuyas frecuencias se distribuyen de forma inversa a la altura geográfica (Dipierri et al., 2000).
Tabla II. Frecuencias alélicas y de haplogrupos en provincias del NOA
Fuentes: ¹Morales et al., 2000; ²Alfaro et al., 2004; ³Muzzio et al., 2010; Muzzio. 2010; Bailliet et al., (2005); ⁶Ramallo et al., (2005; 2009); Dipierri et al., 1998; Tamm et al., 2007; Bravi, 2005.
Los haplogrupos del cromosoma Y muestran mayor diversidad étnica interpoblacional (Tabla II). Mientras que en Salta y Jujuy el haplogrupo predominante es el Q1a3a (Bailliet et al., 2009), linaje mayoritario entre nativos americanos, en Tucumán y Catamarca el haplogrupo más frecuente es el R originario de Europa. En estas dos provincias el haplogrupo DE, frecuente en población africana, alcanza su máximo valor (Tabla II) (Muzzio et al., 2010, Muzio, 2010; Bailliet et al., 2005; Ramallo et al., 2005; 2009). Por el contrario con el ADNmt solo se identificaron 4 haplogrupos presentes en todas las poblaciones americanas y compartidos con poblaciones del este y centro de Asia, lo que indica la pobre diversidad filética de linajes maternos en el NOA semejante a la encontrada en el resto de América (Tabla II) (Bravi, 2005).
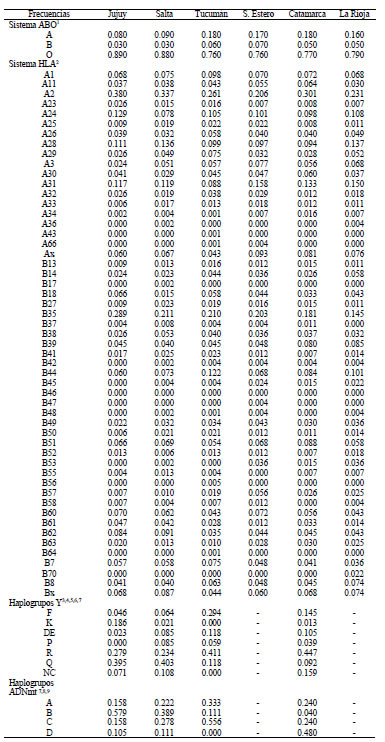
El análisis de agrupamiento de las 6 provincias de NOA utilizando los marcadores sanguíneos (Fig. 1A) o combinados con los linajes maternos y paternos para 4 provincias (Jujuy, Salta, Tucumán y Catamarca) (Fig. 1B) indica relaciones biológicas semejantes. Se identifican dos grandes agrupamientos, por un lado Jujuy y Salta, por otro Santiago del Estero junto a Catamarca y La Rioja, quedando Tucumán en una posición intermedia. Este agrupamiento reflejaría la localización diferencial de las poblaciones alóctonas extracontinentales y la mayor integración cultural de Salta y Jujuy al mundo andino.
Figura 1. Distancias genéticas entre las provincias del NOA calculadas a partir de frecuencias alélicas y de haplogrupos de la Tabla I.
Si bien se observan diferencias entre los porcentajes de aportes continente-específicos de origen africano, europeo y americano al genoma del NOA calculados con los marcadores genéticos de la Tabla III y el análisis genómico total, todos los marcadores indican la misma tendencia caracterizada por un predominio promedio del tronco parental amerindio (43.2%), seguido por el europeo (38%) y el africano (13.3%). Con el análisis genómico total puede apreciarse la afinidad biológica diferencial de Salta, Tucumán y Catamarca con las poblaciones parentales (Fig. 2) y con los grandes troncos lingüísticos (Fig. 3) y poblaciones nativas americanas (Tabla IV). El análisis genómico total también permite evidenciar una contribución sexual diferencial al proceso de mestizaje del NOA. En efecto las proporciones de ancestros africanos y amerindios en las provincias de Tucumán y Catamarca calculadas con los microsatélites del X son mayores que las estimadas con los marcadores autosómicos, situación que se acompaña de una reducción concomitante de los ancestros europeos. Este patrón es consistente con un mestizaje caracterizado predominantemente por la unión de hombres europeos con mujeres autóctonas. Este sesgo sexual en la mezcla europea-nativa ha sido comprobado con los polimorfismo del Y en población jujeña (Dipierri et al., 1998) y múltiples poblaciones mestizas latinoamericanas (Wang et al., 2008).
Tabla III. Porcentajes de mezcla genética por sistemas genéticos y poblaciones parentales en provincias del NOA
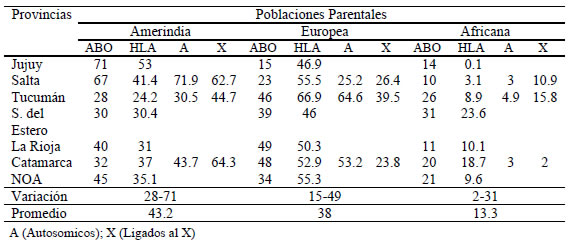
Fuentes: Wang et al., 2009

Figura 2. Distribución del mestizaje por individuos de Salta, Tucumán y Catamarca. La posición de cada punto en el triángulo indica la proporción de ancestría africana, europea y nativa americana
Fuentes: Wang et al., 2009
Tabla IV: Ancestría promedio de Salta, Tucumán y Catamarca con respecto a poblaciones nativas americanas consideradas individualmente (se incluyen los componentes europeo y africano)
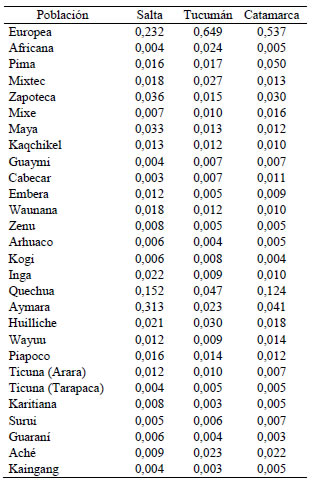
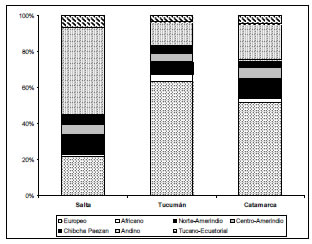
Figura 3. Ancestría promedio de Salta, Tucumán y Catamarca estimada con respecto a grandes troncos lingüísticos nativos americanos (se incluyen los componentes europeo y africano)
LOS APELLIDOS DEL NOA
Los apellidos constituyen un material adecuado para analizar la estructura de las poblaciones en términos de consanguinidad y parentesco y el origen étnico de las mismas. Crow y Mange (1965) sentaron las bases de esta metodología al proponer el coeficiente de consanguinidad por isonimia (F). Posteriormente se desarrollaron otros indicadores: coeficiente de parentesco por isonimia (Lasker, 1977), distancias isonímicas (Lasker, Nei, Euclídea) calculadas a partir de la identidad de apellidos y α de Fisher o riqueza o abundancia de apellidos que permite ponderar la deriva génica por isonimia. Esta metodología fue aplicada para analizar la estructura genética por isonimia de la Argentina (Dipierri et al., 2005a) y del NOA (Dipierri et al., 2007) y sus resultados pueden consultarse en:
http://web.unife.it/progetti/genetica/pdata/Isonymy_Argentina.txt
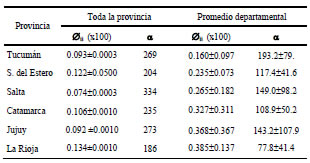
Los apellidos analizados provinieron de 2.576.548 individuos incluidos en los Padrones Electorales (2001) de los 117 Departamentos que integran el NOA. La consanguinidad por isonimia y α se correlacionan negativamente (Dipierri, 2004) por consiguiente valores elevados de consanguinidad y concomitantemente bajos de α indican que la población es endogámica, se encuentra conformada por pocos apellidos diferentes con una elevada frecuencia y que sobre ella se ha producido un fuerte efecto de deriva. La provincia más consanguínea y con menor riqueza de apellidos es La Rioja (Tabla V). Los valores más altos de consanguinidad se observaron en departamentos de la zona andina y los más bajos, como era de esperar, en las capitales provinciales (Fig. 4). La consanguinidad promedio de los 117 departamentos del NOA (0.284± 0.2) duplica al de Argentina (0.135±0.143) y también el promedio departamental de todas las provincias del NOA, excepto Tucumán, situación que se interpreta como el mayor aislamiento relativo de tipo geográfico particularmente de La Rioja y Catamarca (Tabla 5). El α promedio del NOA (130.9±79.8) es inferior al de toda la Argentina (602), al promedio provincial (422) y departamental (358) (Dipierri et al., 2003). Esto estaría indicando que en las provincias del NOA existiría una menor heterogeneidad o riqueza de apellidos con respecto al resto de país, riqueza que oscila entre un α de 17 (Susques y Antofagasta de la Sierra) y 374 (Capital, Tucumán).
Tabla V: Consanguinidad por isonimia (ii) y riqueza de apellidos (α ) por provincias y departamentos del NOA
Fuente: Dipierri, 2004
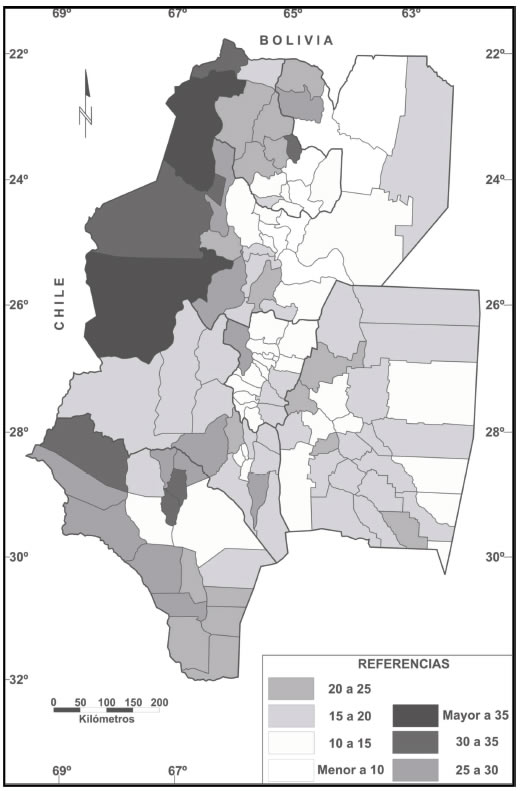
Figura IV: Distribución de la consanguinidad por isonimia ( ii x100) por departamentos del NOA
La variación de las frecuencias de apellidos entre departamentos de una provincia o microdiferenciación interpoblacional puede ser ponderada por el indicador FST propuesto por Relethford (1988). La provincia con mayor microdiferenciación fue Catamarca (0.0369323), seguida por La Rioja (0.0322842), Salta (0,0022710), Jujuy (0,0020552), Santiago del Estero (0,0018646) y Tucumán (0,0010821). Catamarca y La Rioja son las que presentan, coincidentemente, la consanguinidad y el α departamental promedio más alto y más bajo del NOA respectivamente.
La correlación positiva de las distancias genéticas por isonimia con las geográficas entre los 117 departamentos del NOA corresponde al modelo de aislamiento por distancia de Malécot (1948), por lo que la diferenciación genética entre las poblaciones de la región a nivel departamental se produciría por la exogamia de individuos entre poblaciones vecinas a través de migraciones de corta distancia (Cavalli-Sforza et al., 1994). Las distancias genéticas por isonimia permiten diferenciar en el espacio del NOA 10 conglomerados coincidentes con la subdivisión geográfica de la región y con el heterogéneo desarrollo socioeconómico y de las comunicaciones viales registradas en el área.
Estos resultados indican que grandes porciones del territorio regional, especialmente de los subconjuntos departamentales más aislados, no relacionados con las capitales provinciales ni con las ciudades secundarias; concentrarían las poblaciones más consanguíneas y con menor diversidad de apellidos (Dipierri, 2004) (Fig. 4).
Utilizando los apellidos como discriminante étnico es posible examinar la etnogénesis de las poblaciones y en este sentido los apellidos del NOA se muestran muy efectivos para analizar la contribución de las poblaciones parentales a la población actual y su distribución espacial. Observando el origen étnico de los 7 apellidos más frecuentes a nivel provincial, todos son patronímicos de origen español excepto el apellido Mamaní en la Provincia de Jujuy. Los apellidos autóctonos (Erdman, 1964), entre los 7 más frecuentes, se presentan en muchos departamentos de las provincias de Jujuy y Salta y sólo en uno o dos departamentos de las provincias de Catamarca y Tucumán, no observándose ni en La Rioja ni en Santiago del Estero. Los 100 apellidos más frecuentes de Argentina son todos patronímicos españoles, en cambio en el NOA entre los 100 más frecuentes los apellidos autóctonos Mamani y Tolaba ocupan el puesto 22 y 78 con 14395 y 6076 individuos respectivamente (Dipierri, 2004). La distribución de los apellidos por su origen lingüístico y/o geográfico es coherente con la información etnohistórica relativa a la contribución del indigenado en la conformación de las poblaciones del NOA y al proceso de desestructuración diferencial a la que fueran sometidas las poblaciones nativas a partir del siglo XVI (Lorandi y Otonello, 1992). En la Fig. 5 puede observarse el porcentaje de individuos de Salta y Jujuy de acuerdo al origen autóctono o foráneo de los apellidos, y dentro de los foráneos de acuerdo a su origen geográfico. En ambas provincias predominan los individuos portadores de apellidos foráneos y dentro de éstos de aquellos provenientes de sur de Europa (Albeck et al., 2007; 2010; Alfaro et al., 2005). No obstante, esta distribución es coherente con la proporcionada por los datos demográficos (Tabla I) y genéticos (Tablas II y IV, Figs. 2 y 3).
Dado que la transmisión de los apellidos responden al mismo tipo de herencia que los genes, se pueden establecer una asociación entre ambos linajes. En la provincia de Jujuy se analizó la relación entre los alelos del sistema AB0 y la clasificación de los individuos de acuerdo al origen de los apellidos (autóctonos y foráneos) por regiones geográficas distribuidas en un gradiente altitudinal. En todos los niveles de altura la frecuencia del alelo 0 fue mayor en los individuos con apellidos autóctonos y la de los alelos A y B entre los individuos con apellidos foráneos (Dipierri et al., 1999). La misma relación pudo establecerse con los alelos D/d del sistema Rh. El alelo d fue más frecuente en apellidos foráneos y en las tierras bajas, hallazgo que coincide con el hecho de que de acuerdo a Cavalli-Sforza et al. (1994), el haplotipo cde del sistema Rh constituye un excelente indicador de introgresión de genes caucasoides (Morales et al., 2003). Dipierri et al. (1998) utilizando los marcadores holándricos α , DYS199, DYS19, YAP y pSRY analizaron la correlación entre estos polimorfismos y los apellidos de los individuos de poblaciones jujeñas situadas a diferentes niveles altitudinales, observando que el haplotipo αhII-DYS199*T-DYS19*A, específico de amerindios (Bianchi et al., 1998), se encontraba presente en el 35.5% y el 16.7% de los individuos con apellidos autóctonos y foráneos respectivamente. Una correlación opuesta se observó con los haplotipos no amerindios, europeos específicamente (Bianchi et al., 1998). Este trabajo demostró una introgresión de patrilinajes (haplotipos del cromosoma Y) españoles inversamente proporcional a la altura geográfica y la presencia exclusiva de haplotipos mitocondriales amerindios en todos los niveles altitudinales, lo que demuestra un apareamiento paterno direccional en las poblaciones de la Quebrada de Humahuaca. Muzzio (2010) al analizar en varias poblaciones del NOA la relación entre apellidos y haplogrupos del cromosoma Y determina que el "riesgo" relativo de un individuo con un apellido americano para presentar un haplogrupo nativo americano es 3 veces mayor que la probabilidad de que un individuo extranjero presente un haplogrupo foráneo.
CONCLUSIONES
Las poblaciones humanas conforman distintas estructuras parcialmente flexibles, lábiles e interdependientes. El análisis de la interrrelación entre estas estructuras jerárquicas en el NOA, en este caso genética e isonímica, permitió profundizar no solamente su conocimiento, sino también la dinámica poblacional y microevolución de esta particular región geográfica argentina. La estructura genético-isonímica del NOA se caracterizaría por una escasa diferenciación genética interpoblacional a nivel de los grandes centros urbanos, aislamiento por distancia, con una gran diferenciación genética entre sus subpoblaciones departamentales, alta consanguinidad en los departamentos más aislados no integrantes de las ciudades capitales de provincia y grados variables de miscegenación de acuerdo a los marcadores considerados pero que en general revelan una contribución amerindia preponderante, seguida por la europea y africana. Las estructuras genética e isonímica son coherentes con los atributos demográficos, los antecedentes históricos del poblamiento de la región y las particularidades geográficas del NOA. Esta coherencia permitiría concluir que la estructura poblacional del NOA se encontraría determinada por la condición migratoria y el aislamiento relativo de sus poblaciones (Dipierri et al., 2005b).
BIBLIOGRAFÍA
1. Albeck M.E., Alfaro E.L., Dipierri J.E. (2007) Apellidos y estructura Sociodemográfica: cambio y continuidad en Casabindo (siglo XVII al XX). En: Boleda M, Mercado MC (Compiladores). Seminario Internacional de Población y Sociedad en América Latina (SEPOSAL), 8-10 Junio, Tomo I, pp. 332-346. [ Links ]
2. Albeck M.E., Alfaro E.L., Dipierri J.E. (2010) Origen, diversidad y frecuencia de los apellidos en Salta (República Argentina). Manuscrito. [ Links ]
3. Alfaro E., Dipierri J., Gutiérrez N., Vullo C (2004). Frecuencias génicas y haplotípicas del sistema HLA en el Noroeste Argentino. Antropo 6:15-23. [ Links ]
4. Alfaro E.L., Albeck M.E., Dipierri J.E. (2005). Apellidos en Casabindo entre los siglos XVII y XX. Continuidades y cambio. Andes 16: 147-166. [ Links ]
5. Alfaro E.L., Dipierri J.E., Gutiérrez N.J., Vullo C.M. (2005) Genetic structure and admixture in urban populations of the Argentina North-West. Ann Hum Biol 32(6):724-737. [ Links ]
6. Aschero C. (1980) Comentarios acerca de un fechado radiocarbónico del sitio de Inca Cueva IV (Dpto. de Humahuaca-Jujuy) RSAA Tomo XIV. [ Links ]
7. Bailliet G., Ramallo V., Alfaro E.L., Dipierri J.E., Bianchi N.O. (2005) Caracterización de linajes paternos en muestras provenientes de tres provincias del NOA. BAG (Actas del XXXIV Congreso Argentino de Genética), XVII:181. [ Links ]
8. Bailliet G., Ramallo V., Muzzio M., García A., Santos M.R., Alfaro E.L., Dipierri J.E., Salceda S., Carnese F.R., Bravi C.M., Bianchi N.O., Demarchi D.A. (2009) Brief communication: Restricted geographic distribution for Y-Q* paragroup in South America. Am J Phys Anthropol 140(3):578-582. [ Links ]
9. Bazán A.R. (1986) La Historia del Noroeste Argentino. Editorial Plus Ultra. Bs.As. Argentina. [ Links ]
10. Bianchi N.O., Catanesi, C.I., Bailliet, G., Martinez-Marignac V.L., Bravi C.M., Vidal-Rioja L.B., Herrera R.J., Lopez-Camelo J. (1998) Characterisation of ancestral and derive Y-chromosome haplotypes of New World Native Populations. Am J Hum Genet 63:1862-1871. [ Links ]
11. Boisier S. (1992) El difícil arte de hacer región. Centro de Estudios Regionales Andino Bartolomé de las Casas. Cuzco, Perú [ Links ].
12. Boleda M. (1993) La población del Noroeste Argentino. Historia y actualidad. Legasa. Buenos Aires. [ Links ]
13. Boleda M. (1998) Ciudades del Noroeste Argentino. Alianza Editorial, Buenos Aires. [ Links ]
14. Bolsi A.S., Pucci R (1997) Problemas poblacionales del Noroeste Argentino. Instituto de Estudios Geográficos. Facultad de Filosofía y Letras. Junta de Andalucía. Universidad Nacional de Tucumán. [ Links ]
15. Bravi C.M. (2005) Análisis de linajes maternos en poblaciones nativas americanas. Tesis de Doctorado, Facultad de Ciencias Naturales y Museo, UNLP. [ Links ]
16. Cabrera A.L., Willink A (1973) Biogeografía de América Latina. Monografía Nº 13. OEA. [ Links ]
17. Castro de Guerra D. (1987) Uso de apellidos para determinar estructura de población en pueblos negros de la costa norcentral de Venezuela. Tesis de Grado Magister Scientiarum en Biología. Centro de Estudios Avanzados. IVIC, Caracas. [ Links ]
18. Cavalli-Sforza L.L., Menozzi P. y Piazza A. (1994) The history and geography of human genes. Princeton University Press, New Jersey. [ Links ]
19. C.F.I. (Consejo Federal de Inversiones) (1963) Relevamiento de la estructura económica de la Argentina. Centro de Investigaciones Torcuato di Tella. Buenos Aires. [ Links ]
20. Comadrán Ruiz (1969) Evolución demográfica durante el periodo hispano (1535-1810). EUDEBA. Buenos Aires. [ Links ]
21. Crow J.E., Mange A.P. (1965) Measurements of inbreeding from the frequency of marriages between persons of the same surnames. Eugenic Quarterly 12:190:203 [ Links ]
22. Dipierri J.E. (2004) Apellidos del Noroeste Argentino: Distribución, Isonimia, Estructura y Dinámica Poblacional. Tesis de Maestría. Maestría en Teoría y Metodología de las Ciencias Sociales. Facultad de Humanidades y Ciencias Sociales. Universidad Nacional de Jujuy. [ Links ]
23. Dipierri J.E., Alfaro E., Dugoujon J.M., Constans J. (2000) GM and KM inmunoglobulin allotypes and other serum genetic markers (Hp, Gc, Pi and Tf) among South American living at different altitudes (Jujuy Province, Argentina): Admixtures estimates. Hum Biol 72(2):305-319. [ Links ]
24. Dipierri J.E., Alfaro E., Martinez Marignac V., Bailliet G., Bravi C., Cejas S., Bianchi N.O. (1998) Paternal directional matting in two Amerindian subpopulations located a different altitudes in Northwest Argentina. Hum Biol 70(6):1000-1010. [ Links ]
25. Dipierri J.E., Alfaro E.L., Scapoli C., Mamolini E., Rodríguez Larralde A., Barrai I. (2005a) Surnames in Argentina: A population study through isonymy. Am J Phys Anthropol 128(1):199-299. [ Links ]
26. Dipierri J.E., Alfaro, E., Bejarano, I.F. (1999) Surnames, ABO system and miscegenation in highland populations of Province of Jujuy (Nortwest Argentine). Homo 50(1):14-20. [ Links ]
27. Dipierri J.E., Rodríguez Larralde A., Alfaro E.L., Andrade A., Chaves E., Barrai I. (2005b). Distribución de apellidos y migración en el noroeste argentino. Antropo 10:35-50. [ Links ]
28. Dipierri J.E., Rodríguez Larralde A., Alfaro E.L., Barrai I. (2007) Isonymic structure of the Argentine Northwest. Ann Hum Biol 34(4):498-503. [ Links ]
29. Dipierri, J.E., Alfaro E. (1994) Aspectos genéticos y bioantropológicos de las poblaciones nativas del Noroeste de la República Argentina. Mendeliana (Suplemento) XI (1):12-15 [ Links ]
30. Erdman L.E. (1964) Patronimia del Noroeste Argentino. Rev. Instituto de Antropología (Universidad Nacional de Córdoba), Tomo III:121-134. [ Links ]
31. Harrison G.A., Boyce AJ (1972) The structure of human populations. Clarendon Press Oxford. [ Links ]
32. Hernández I. (1992) Los indios de la Argentina. Colecciones Mapfre 1492, Madrid. [ Links ]
33. Hilhorst J.G.M. (1971) Planeamento regional. Enfoque sobre sistemas. Biblioteca de Ciências Sociais. Zahar Editores. Río de Janeiro. Brasil. [ Links ]
34. Lasker G.W. (1977) A coefficient of relationship by isonymy: a method for estimating the genetic relationship between populations. Hum Biol 49:489-493. [ Links ]
35. Lasker G.W., Mascie-Taylor CGN (1993) Research strategies in human biology: field and survey studies. Cambridge University Press. [ Links ]
36. Llop E., Rothhammer F. (1988) A note on presence of blood groups A and B in Pre-Columbian South America. Am J Physc Anthropol 75:107-111. [ Links ]
37. Lorandi A.M., Otonello M. (1992) Argentina: completando la historia. Ciencia e Investigación. Tomo 45 (2):84-99. [ Links ]
38. Macbeth H., Collison P. (2002) Human Populations Dynamics. Cross-Disciplinary Perspectives. Cambridge University Press. [ Links ]
39. Malécot G. (1948) Les mathématique de l'Hérédité. Paris. Masson. [ Links ]
40. Martinez-Marignac V., Bailliet G., Dipierri J.E., Alfaro E., López-Camelo J.S. y Bianchi N.O. (2001) Variabilidad y antigüedad de linajes holándricos en poblaciones Jujeñas. Rev. Arg. Antrop. Biol. 3(1): 49-63. [ Links ]
41. Morales J., Alfaro E., Dipierri J., Bejrano I. (2003). Apellidos y sistema Rh (D/d) en poblaciones jujeñas. Rev Arg Antrop Biol 5(2):7-13. [ Links ]
42. Morales J., Dipierri J.E., Alfaro E., Bejarano I.F. (2000) Distribution of the ABO system in the Argentine Northwest: Miscegenation and genetic diversity. Interciencia 25(9):432-435. [ Links ]
43. Muzzio M. (2010) Isonimia y cromosoma Y: ¿concordancia o conflicto?. Tesis de Doctorado. Facultad de Ciencias Naturales y Museos. UNLP. [ Links ]
44. Muzzio M., Ramallo V., Bailliet G. (2010) Comparación de apellidos y linajes de cromosoma Y en 4 provincias del NOA. Manuscrito. [ Links ]
45. Ortega Valcarcel J. (2000) Los horizontes de la geografía. Teoría de la geografía. Editorial Ariel S.A. Barcelona, España. [ Links ]
46. Pontussi E.P. (1995) Geografía del Noroeste Argentino. Facultad de Ciencias Naturales. Universidad Nacional de Salta. [ Links ]
47. Pucci R. (1998) El tamaño de la población aborigen del Tucumán en la época de la conquista: balance de un problema y propuesta de una estimación. Población y Sociedad Nº 5: 239-270. [ Links ]
48. Ramallo V., Alfaro E.L., Dipierri J.E., Bianchi N.O., Bailliet G. (2005) Linajes holándricos en tres provincias del NOA. Rev Arg Antrop Biol 7(1):65. [ Links ]
49. Ramallo V., Mucci J.M., García A., Muzzio M., Motti J.M.B., Santos M.R., Pérez M.E., Alfaro E.L., Dipierri J.E., Demarchi D.A., Bravi C.M., Bailliet G. (2009) Comparison of Y chromosome haplogroup frequencies in 8 Provinces from Argentina. For Sci Inter: Genetics Supplement Series, 2(1):431-432. [ Links ]
50. Relethford J.H. (1988) Estimation of kinship and genetic distance from surnames. Hum Biol 60:475-492. [ Links ]
51. Rothhammer S., Silva S.M., Callegari Jacques S., Llop E., Salzano F.M. (1997) Gradients of HLA diversity in South America Indians. Ann Hum Biol 24:197-208. [ Links ]
52. Ruíz M.S. (1993) Prehistoria y arqueología regional. En: Campi, D (Editor) Jujuy en la Historia. Avances de Investigación I. Unidad de Investigación en Historia Regional, FHYCS-UNJU. [ Links ]
53. Salzano F.M., Callegari Jacques S. (1988) South American Indians. A case study in human for recovering genetic information. Hum. Biol. 55:219-255. [ Links ]
54. Tamm E., Kivisild T., Reidla M., Metspalu M., Glenn Smith D., Mulligan C.J., Bravi C.M., Rickards O., Martinez-Labarga C., Khusnutdinova E.K., Fedorova S.A., Golubenko M.V., Stepanov V.A., Gubina M.A., Zhadanov S.I., Ossipova I.P., Damba L., Voevoda M.I., Dipierri J.E., Villems R., Malhi R.S. (2007) Beringian Standstill and Spread of Native American Founders. PloS ONE 9(829): 1-6. [ Links ]
55. Vogel F., Motulsky A.G. (1982) Human Genetics. Problems and Approaches. Springer-Verlag, Berlin. [ Links ]
56. Wang S., Ray N., Rojas W., Parra M.V., Bedoya G., Gallo C., Poletti G., Mazzotti G., Hill K., Hurtado A.M., Camrena B., Nicolini H., Klitz W., Barrantes R., Molina J.A., Freimer N.B., Bortolini M.C., Salzano F.M., Petzl-Erler M.L., Tsuneto L.T., Dipierri J.E., Alfaro E.L., Bailliet G., Bianchi N.O., Llop E., Rothhammer F., Excoffier L., Ruiz-Linares A. (2008) Geographic patterns of genome admixture in latin american mestizos. Plos One 4(3):1-9. [ Links ]
57. Yasuda N., Morton E.N. (1967) Studies on human population structure. Proceedings of the 3rd International Congress of Human Genetics. Ed: Crow JF and Nell JV, pp. 249-265. Baltimore: Johns Hopkins. [ Links ]

