










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkBAG. Journal of basic and applied genetics
On-line version ISSN 1852-6233
BAG, J. basic appl. genet. vol.23 no.2 Ciudad Autónoma de Buenos Aires Dec. 2012
RESEARCH
La aptitud combinatoria específica en el maíz (Zea mays L. ssp. mays) para doble propósito en Río Cuarto, Córdoba, Argentina
Di Santo H. 1, Castillo E. 1, Ferreira A. 1, Grassi E. 1, Ferreira V. 1
1Genética, Facultad de Agronomía y Veterinaria. Universidad Nacional de Río Cuarto. egrassi@ayv.unrc.edu.ar
RESUMEN
Los maíces doble propósito son útiles en los establecimientos de producción mixta ya que se adaptan a los propósitos de cosecha de grano y alimento directo o diferido para el ganado. En la Universidad Nacional de Río Cuarto se ejecuta un proyecto de mejoramiento de maíz forrajero a partir de poblaciones locales. El objetivo fue determinar la aptitud combinatoria específica (ACE) de líneas endocriadas. Se realizó el cruzamiento dialélico y durante tres campañas se analizó la ACE de las líneas a través de sus híbridos, empleando 18 caracteres; el diseño fue en bloques completos al azar con tres repeticiones y los datos se analizaron mediante ANOVA. Los ensayos de ACE mostraron valores positivos y significativos para la mayoría de los caracteres evaluados. Esto permitió identificar y clasificar los híbridos simples según su potencial aptitud de uso. Del total de híbridos simples se eligieron siete, uno por su aptitud para doble propósito, tres por su alta producción de forraje diferido o grano y tres para ensilado en estado lechoso-pastoso. Se considera necesario confirmar el comportamiento y productividad de los híbridos simples promisorios variando ambientes regionales y condiciones agronómicas contrastándolos con testigos comerciales a fin de verificar el potencial de producción así como su estabilidad.
Palabras clave: Maíz; Líneas endocriadas; Híbridos simples; Usos agronómicos.
ABSTRACT
The dual purpose maize is useful in mixed farming establishments because they are suitable to the purposes of grain crop and direct or deferred feed for livestock. A breeding plan to improve land maize populations for forage use is carried out at the National University of Río Cuarto. The aim of this work was to determine the Specific Combining Ability of inbred lines through the productive performance of their single-cross hybrids. A diallel cross was performed, and eighteen traits of the single cross hybrids were studied during three years. A completely randomized block design with three replicates was used, and ANOVA for data analysis was carried out. Specific combining ability trials showed significant positive values for most traits. This allowed identifying and classifying single-cross hybrids according to their potential usability. Seven single-cross hybrids were chosen: one for dual purpose aptitude, three for grain yield or high deferred forage production and three for silage in milk-dough grain stage. It is considered necessary to confirm the performance and productivity of promising single-cross hybrids varying regional environments and agronomic conditions set against commercial simple cross hybrids to verify the production potential and stability.
Key words: Maize; Inbred lines; Single cross-hybrids; Agronomic uses.
INTRODUCCIÓN
El maíz, Zea mays L. ssp. mays (Iltis y Doebley, 1980), es uno de los cultivos más antiguos, ocupa el segundo lugar a nivel mundial en volumen de producción y tiene gran importancia socio-económica ya sea para alimento humano o animal o como fuente de un gran número de productos industriales (Watson, 1988; Fussell, 1999).
La planta de maíz es un excelente forraje para el ganado bovino de carne y de leche. Se utiliza como forraje en varias etapas del crecimiento, desde estado juvenil de pasto hasta la emisión de la panoja o más adelante. Para obtener la máxima producción de Materia Seca (MS) digestible por hectárea con la máxima energía (forraje energético), el estado de grano lechoso-pastoso es el más adecuado y supera a cualquier otro cultivo anual. En ese estado es el cultivo ideal para ensilar (Gallais et al., 1976; Paliwal et al., 2001).
El cultivo está ampliamente difundido en los establecimientos de producción mixta de la región subhúmeda-semiárida argentina. En la alimentación animal se emplea como grano, forraje verde, silaje en estado de grano lechoso-pastoso y pastoreo en pie cuando se prevé mala cosecha de grano o del rastrojo. La obtención de materiales con diferentes aptitudes de utilización para los sistemas mixtos regionales debería combinar la producción de grano con caracteres de gramíneas forrajeras, tales como alta cantidad de materia seca total, tolerancia a sequía, macollamiento, rebrote y plasticidad (Reynoso, 1996). La definición más simple para un maíz doble propósito es el que considera la producción de grano y como alternativa el pastoreo en estado vegetativo y reproductivo temprano, el picado y el pastoreo del cultivo diferido.
La superficie sembrada en la Argentina fue de aproximadamente 4,56 x 106 has en la campaña 2010/11, mientras que en la región subhúmeda-semiárida fue de 1,657 x 106 has, de las cuales, 68,9 % correspondieron a la provincia de Córdoba, 8,4 % a San Luis y 22,6 % a La Pampa. La región mencionada aportó el 29,7 % de la producción nacional (SIIA, 2012). Los registros históricos indican que el 90 % del maíz se cosecha como grano para consumo humano o animal y el resto se destina a silaje o consumo directo o diferido por el ganado; este último destino ocupa alrededor de 468.000 has en la región subhúmeda-semiárida (INDEC, 2005). Sin embargo, en las últimas dos campañas se estima una superficie de maíz picado para silaje de entre 750.000 y 850.000 has (CACF, 2012; Maizar, 2012).
En 1908, G.H. Shull fue el primero que sugirió la utilización de los híbridos F1 producto de la cruza entre líneas endocriadas (en Crow, 1998). Sprague y Tatum (1942) definieron los términos aptitud combinatoria general (ACG) y aptitud combinatoria específica (ACE) en relación al comportamiento relativo de las líneas al ser cruzadas, donde ACG designa el comportamiento promedio de una línea en combinaciones híbridas, y ACE designa la desviación de cada combinación con respecto a la ACG de los progenitores (Hallauer y Miranda Filho, 1988; Quemé de León et al., 1991).
El conocimiento de la aptitud combinatoria de los progenitores permite seleccionar aquellos de buen comportamiento en una serie de cruzamientos e identificar combinaciones específicas de comportamiento superior (Gutiérrez del Río et al., 2004; Castañón-Nájera et al., 2005; De la Cruz-Lázaro et al., 2010). Una vez que se han seleccionado las líneas endocriadas teóricamente mejores basándose en su buena aptitud combinatoria general, es necesario identificar los cruzamientos simples, de tres vías o dobles que producirán los mayores rendimientos (Sprague y Tatum, 1942; Allard, 1978). El desarrollo de cultivares híbridos simples requiere analizar los resultados de los cruzamientos dialélicos (Russel, 1973; Allard, 1978; Hallauer y Miranda Filho, 1988).
La Argentina posee como antecedentes la creación de una variedad de maíz doble propósito y macolladora en la EEA Bordenave, donde se obtuvo el cultivar "Don Faustino INTA" (Gorostegui, 1971) y se realizaron varios trabajos con distinto germoplasma de maíz y sus parientes silvestres en el Instituto Fitotécnico Santa Catalina de la UN de La Plata, EEA Pergamino-INTA, UN de La Pampa, UN de Lomas de Zamora (Magoja y Pischedda, 1988; Troiani et al., 1988; Paccapelo y Molas, 1996; Torrecillas y Bertoia, 2000).
A partir de un proyecto destinado al mejoramiento de maíz para uso forrajero iniciado con tres poblaciones locales macolladoras, en la UN Río Cuarto se efectuaron seis ciclos de selección recurrente de medios hermanos (Lonnquist, 1964), obteniendo el Grado de Determinación Genética y los coeficientes de variación genética de líneas S1 y S2 (Grassi et al., 2006). En base a una prueba top cross se evaluó la ACG y se eligieron cinco líneas para la formación de híbridos simples y estudiar su validación agronómica (Grassi et al., 2009; Grassi, 2009).
El objetivo propuesto en este trabajo fue determinar la aptitud combinatoria específica de líneas endocriadas de maíz, seleccionadas por su aptitud combinatoria general, a través de los híbridos simples obtenidos a partir del cruzamiento dialélico, identificando los más aptos para doble propósito en base a caracteres morfofisiológicos y de producción.
MATERIALES Y MÉTODOS
Los ensayos de aptitud combinatoria especifica (ACE) se realizaron en el Campo de Docencia y Experimentación de la UN de Río Cuarto (33° 06' 22'' S; 64° 17' 52'' O) ubicado a 443 msnm en la localidad de Las Higueras, Departamento Río Cuarto, Córdoba. El suelo es un Haplustol típico con 1,6 % de materia orgánica. El área cuenta con una temperatura media anual de 16,5° C. El período libre de heladas es de 240 días (entre septiembre y mayo). Las precipitaciones poseen un valor medio de 800 mm anuales con un régimen monzónico irregular con aproximadamente 80 % de las precipitaciones concentradas en el semestre más cálido (octubre a marzo).
Durante las campañas agrícolas 2004/2005, 2006/07 y 2007/08 se sembraron cinco líneas endocriadas, elegidas en base a una prueba top cross para la determinación de la ACG, y los 20 híbridos simples obtenidos a partir del cruzamiento dialélico completo de dichas líneas. Los ensayos se sembraron el día 19 de octubre de 2004, 10 de noviembre de 2006 y 7 de noviembre de 2007. Los materiales de dispusieron en un diseño de bloques completos al azar, con tres repeticiones. Cada parcela consistió en dos surcos a 0,70 metros de separación y una longitud de 3 metros (4,20 m2). Se sembraron a razón de cinco semillas por metro lineal, con doble golpe, para (con un raleo posterior) garantizar una densidad de cosecha de aproximadamente 71.000 plantas ha-1. La unidad experimental consistió en plantas individuales realizándose tres submuestreos por parcela.
Se evaluaron caracteres morfofisológicos, en estado R3-R4 y al final del ciclo del cultivo:
- Morfofisiológicos: altura total (m) de la planta, número de tallos por planta, número de espigas por planta, número de espigas por tallo.
- En estado R3-R4: peso verde (g) de planta entera, peso verde (g) de espigas sin chala por planta, porcentaje de espiga verde sin chala por planta, porcentaje de espiga seca sin chala por planta, porcentaje de materia seca de espiga sin chala por planta, peso seco (g) de planta entera, peso seco (g) de espigas sin chala por planta, porcentaje de materia seca total.
- Al final del ciclo del cultivo: peso seco (g) de planta entera, peso seco (g) de espigas sin chala por planta, número de granos por planta, peso (g) de granos por planta, peso (g) de 1000 granos, índice de cosecha.
Los valores obtenidos de los ensayos de ACE se analizaron mediante un análisis dialélico conjunto (Griffins, 1956) utilizando el siguiente modelo:
Yij = µ + gi + gj + sij + ak + gaik + gajk + saijk + rl + εij,
donde, µ= media general del ensayo; gi= efectos de la aptitud combinatoria general asociada al progenitor i; gj= efectos de la aptitud combinatoria general asociada al progenitor j; sij= efectos de la aptitud combinatoria especifica entre los progenitores i y j; ak= efecto del ambiente k; gaik y gajk = efectos de la interacción entre la ACG de los progenitores i y j y el ambiente k; saijk= interacción de la aptitud combinatoria especifica entre los progenitores i y j y el ambiente k ; rl= efecto de la repetición l; εij= error experimental. Se aclara que en este modelo Yij y εij son, respectivamente, la media experimental y el error aleatorio medio asociados a los tratamientos de orden ij (Cruz, 2006).
Este modelo, correspondiente al modelo experimental I propuesto por Griffins (1956), incluye los progenitores, híbridos F1 con recíprocos, aleatorizados en los bloques respectivos. El análisis se llevó a cabo utilizando el aplicativo computacional en genética y estadística GENES (Cruz, 2006). Los significados de los efectos genéticos de ACG y ACE de las variables medidas se evaluaron con una prueba "t", a través de su error estándar. El error estándar de una media de tratamiento se calculó a partir de la raíz cuadrada del cociente entre el cuadrado medio del error del residuo y el número de repeticiones (Steel y Torrie, 1988).
RESULTADOS Y DISCUSIÓN
Los ensayos se desarrollaron bajo condiciones hídricas adecuadas, donde las precipitaciones de los tres ciclos superaron al promedio histórico correspondiente. Las campañas 2004/05 y 2006/07 se iniciaron con menor recarga hídrica en el perfil del suelo debido a un atraso en el inicio de las lluvias primaverales. Durante el período crítico del cultivo, las precipitaciones fueron abundantes por lo que el cultivo no experimentó periodos de deficiencia de humedad edáfica.
Los valores medios y desvíos para los caracteres considerados en los tres años de ensayo se resumen en la Tabla 1. Algunos de los caracteres de producción medidos en R3-R4 y fin de ciclo se proyectaron a hectárea con el objetivo de dimensionar los valores obtenidos. La materia seca total en estado lechoso-pastoso (62.900 kg ha-1) fue similar y en algunos casos superior a los encontrados por Arias et al. (2003), Centeno et al. (2009; 2010), Di Nucci de Bedendo et al. (2009) y De León et al. (2011) empleando distintos híbridos comerciales.
Tabla 1. Valores medios y desvíos estándares para caracteres analizados en Ensayos de Aptitud Combinatoria Específica (EACE) de líneas endocriadas e híbridos simples de maíz en tres ciclos de producción (2004/05, 2006/07, 2007/08). Río Cuarto, Córdoba.

Ref. : AP= altura de planta (cm); TP= número de tallos por planta; EP= número de espigas por planta; ET= número de espigas por tallo; PVTLP= peso verde (g) de planta entera en R3-R4; PVELP= peso verde (g) de espigas sin chala por planta en R3-R4; %EVLP= % de espiga verde por planta en R3-R4; % ESLP= % de espiga seca por planta en R3-R4; % MSELP= % de materia seca de espigas sin chala en R3-R4; PSTLP= peso seco (g) de planta entera en R3-R4; PSELP= peso seco (g) de espigas sin chala/planta en R3-R4; % MSLP= % de materia seca total en R3-R4; PSTFC= peso seco (g) de planta entera en fin de ciclo; PSEFC= peso seco (g) de espigas sin chala por planta en fin de ciclo; NG= número de granos por planta; PG= peso (g) de granos por planta; P1000= peso (g) de 1000 granos; IC= índice de cosecha.
Los valores de MS total observada a fin de ciclo (24.345 kg ha-1) se corresponden con los valores observados en ensayos sobre maíz para uso como pastura diferida para la alimentación de ganado bovino (Maresca et al., 2006; Castaldo et al., 2009). El peso de grano promedio registrado (10.106 kg ha-1) fue superior o similar al obtenido en diferentes localidades de la provincia de Buenos Aires, en suelos con 2,5-3% de materia orgánica y 180-200 kg ha-1 de urea (Cirilo, 2004; Ferraris, 2004; Ventimiglia y Torrens Baudrix, 2012).
Los valores de F y el significado obtenido de los análisis de varianza conjuntos de los caracteres medidos en los híbridos simples se presentan en las Tablas 2, 3 y 4. La ACG fue altamente significativa para los caracteres altura de planta y espigas por planta y significativa para espigas por tallo y peso verde de espigas en estado R3-R4. La ACE fue muy altamente significativa en altura de planta, peso verde de planta entera y peso verde de espigas en estado R3-R4; los caracteres tallos por planta y espigas por tallo presentaron ACE significativa. La ACE recíproca fue no significativa en los cuatro caracteres (Tabla 2). No se observaron interacciones con el año, salvo para altura de planta donde la ACG tuvo interacción altamente significativa.
Tabla 2. Valores de F y significación del análisis de varianza conjunta para caracteres morfofisiológicos en los Ensayos de Aptitud Combinatoria Específica (EACE) de híbridos simples y líneas endocriadas de maíz (2004/05, 2006/07, 2007/08). Río Cuarto, Córdoba.
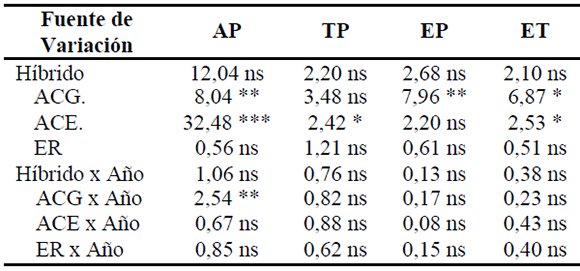
REF: AP= altura de planta (cm); TP= número de tallos por planta; EP= número de espigas por planta; ET= número de espigas por tallo; ACG= aptitud combinatoria general; ACE. = aptitud combinatoria específica directa; ER= ACE efecto recíproco. *, **, *** = significativo para p<0,05, p<0,01 y p<0,001, respectivamente; ns = no significativo.
Tabla 3. Valores de F y significación del análisis de varianza conjunta para caracteres en estado R3-R4 en los Ensayos de Aptitud Combinatoria Específica (EACE) de híbridos simples y líneas endocriadas de maíz (2004/05, 2006/07, 2007/08). Río Cuarto, Córdoba.
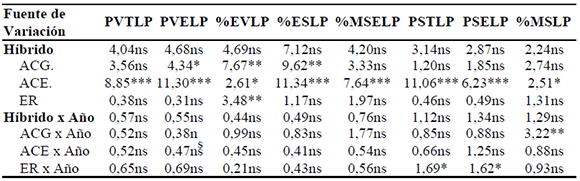
Ref: PVTL= peso verde (g) de planta entera en R3-R4; PVELP= peso verde (g) de espigas sin chala por planta en R3-R4; % EVLP= % de espiga verde por planta en R3-R4; % ESLP= % de espiga seca por planta en R3-R4; % MSELP= % de materia seca de espigas sin chala en R3-R4; PSTLP= peso seco (g) de planta entera en R3-R4; PSELP= peso seco (g) de espigas sin chala/planta en R3-R4; % MSLP= % de materia seca total en R3-R4; ACG= aptitud combinatoria general; ACE= aptitud combinatoria específica directa; ER= ACE. efecto recíproco. *, **, *** = significativo para p < 0,05, p < 0,01 y p < 0,001, respectivamente; ns = no significativo.
Tabla 4. Valores de F y significación del análisis de varianza conjunta para caracteres en fin de ciclo en los Ensayos de Aptitud Combinatoria Específica (EACE) de híbridos simples y líneas endocriadas de maíz (2004/05, 2006/07, 2007/08). Río Cuarto, Córdoba.
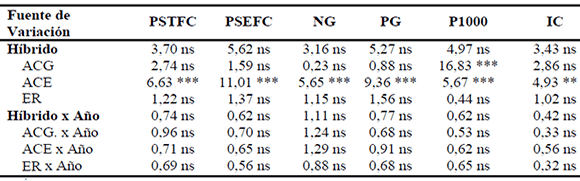
Ref. : PSTFC= peso seco (g) de planta entera en fin de ciclo; PSEFC= peso seco (g) de espigas sin chala por planta en fin de ciclo; NG= número de granos por planta; PG= peso (g) de granos por planta; P1000= peso (g) de 1000 granos; IC= índice de cosecha; ACG= aptitud combinatoria general; ACE= aptitud combinatoria específica directa; ER= ACE efecto recíproco. *, **, *** = significativo para p < 0,05, p < 0,01 y p < 0,001, respectivamente; ns= no significativo.
Los efectos de ACG fueron altamente significativos para porcentaje de espiga verde y porcentaje de espiga seca en estado R3-R4. Los efectos de ACE fueron significativos para los seis caracteres (muy altamente significativos en porcentaje de espiga seca, porcentaje de materia seca de espiga, peso seco de planta entera y peso seco de espigas en estado R3-R4). El ER presentó significación únicamente en porcentaje de espiga verde en estado R3-R4.
La interacción ACG x Año fue altamente significativa en % de materia seca total y la interacción de ER x Año fue significativa en los caracteres peso seco de planta entera y peso seco de espigas. Los caracteres de fin de ciclo (Tabla 4) mostraronm efectos de ACG muy altamente significativos en el peso de 1000 semillas, mientras que los efectos de ACE fueron muy altamente significativos en todos ellos, salvo para índice de cosecha que se mostraron altamente significativos. Los efectos de ACE recíproca y las interacciones de aptitud combinatoria y año fueron no significativos en todos los casos.
Caracteres morfofisiológicos
Los valores de ACE para los caracteres morfofisiológicos evaluados previo al corte en R3-R4 se presentan en la Tabla 5. Trece híbridos simples presentaron efectos significativos para altura de planta, donde se destacan 4x20, 20x4, 20x26 y 26x3 como los de mayor ACE para este carácter. Los híbridos 3x4, 4x3, 26x4 y 26x20 acompañan ese efecto con la relación espiga/tallo y 28x4 con número de espigas por planta, respectivamente. No existen valores significativos de efecto de ACE para tallos por planta.
Tabla 5. Valores de aptitud combinatoria especifica para caracteres morfofisiológicos en 20 híbridos simples y 5 líneas endocriadas progenitoras de maíz (2004/05, 2006/07, 2007/08). Río Cuarto, Córdoba.

Ref. : AP= altura de planta (cm); TP= número de tallos por planta; EP= número de espigas por planta; ET= número de espigas por tallo. Significación a través de una prueba t: asignificativo al 10 %; *significativo al 5 %; **significativo al 1 %. (1)híbrido simple analizado a través de su efecto recíproco.
Caracteres evaluados en estado R3-R4 (grano lechoso-pastoso)
En la Tabla 6 se resumen los valores de ACE de los híbridos analizados para los caracteres antes descriptos. En total, 10 HS muestran un efecto de ACE significativa en al menos un carácter. Se destaca el HS 3x26 por tener la mayor cantidad de caracteres con valores significativos de ACE positivos; éste y 28x4 son los híbridos de mayor efecto de ACE para peso verde de espiga. El HS 26x3 sobresalió en los caracteres peso seco de planta entera y peso seco de espigas, mientras que 20x4 se destaca en porcentaje de materia seca de espiga y ambos pesos secos.
Se observa que el híbrido 28x4, que involucra como progenitor femenino una línea con ACG negativa (línea 28), presentó un alto valor de ACE para peso verde de planta entera. Este efecto fue descripto en maíces tropicales y subtropicales donde progenitores con efectos bajos o negativos de ACG produjeron cruzas con alto valores en los caracteres respectivos (Guillén-de la Cruz et al., 2009; De la Cruz-Lázaro et al., 2010).
Tabla 6. Valores de aptitud combinatoria específica para caracteres en estado R3-R4 en 20 híbridos simples y 5 líneas progenitoras de maíz (2004/05, 2006/07, 2007/08). Río Cuarto, Córdoba.
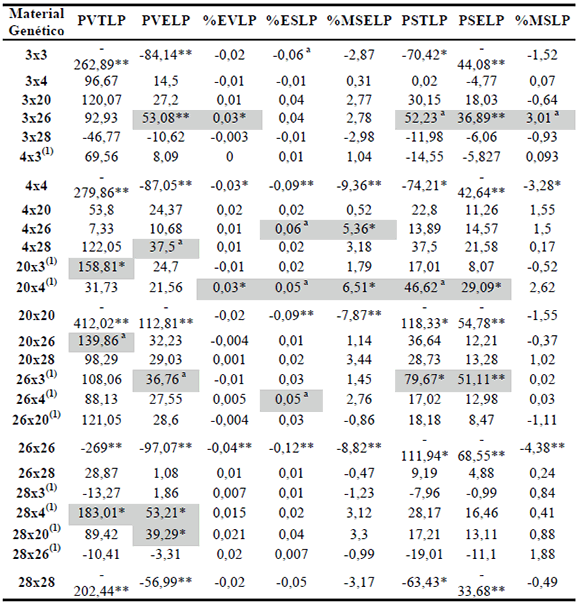
Ref: PVTLP= peso verde (g) de planta entera en R3-R4; PVELP= peso verde (g) de espigas sin chala por planta en R3-R4; % EVLP= % de espiga verde por planta en R3-R4; % ESLP= % de espiga seca por planta en R3-R4; % MSELP =% de materia seca de espigas sin chala en R3-R4; PSTLP= peso seco (g) de planta entera en R3-R4; PSELP= peso seco (g) de espigas sin chala/planta en R3-R4; % MSLP= % de materia seca total en R3-R4. Significación a través de una prueba t: asignificativo al 10 %; *significativo al 5 %; **significativo al 1 %. (1) híbrido simple analizado a través de su efecto recíproco.
Tabla 7. Valores de aptitud combinatoria específica para los caracteres analizados en fin de ciclo en 20 híbridos simples y 5 líneas endocriadas progenitoras de maíz (2004/05, 2006/07, 2007/08). Río Cuarto, Córdoba.

Ref: PSTFC= peso seco (g) de planta entera en fin de ciclo; PSEFC= peso seco (g) de espigas sin chala por planta en fin de ciclo; NG= número de granos por planta; PG= peso (g) de granos por planta; P1000= peso (g) de 1000 granos; IC= índice de cosecha. Significación a través de una prueba t: asignificativo al 10 %; *significativo al 5 %; ** significativo al 1 %. (1) híbrido simple analizado a través de su efecto recíproco.
Tabla 8. Número de caracteres con valor positivo y significativo de aptitud combinatoria específica para los híbridos simples de maíz seleccionados (2004/05, 2006/07, 2007/08). Río Cuarto, Córdoba.
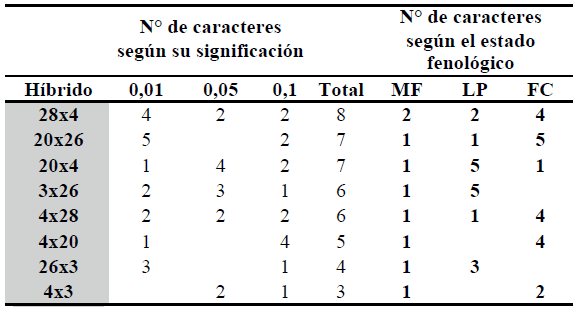
Ref. : MF= caracteres morfofisiológicos; LP= caracteres medidos en estado R3-R4; FC= caracteres medidos a fin de ciclo.
CONCLUSIONES
Los ensayos permitieron determinar la aptitud combinatoria específica de cinco líneas endocriadas de maíz a través de los 20 híbridos simples originados en el cruzamiento dialélico. Se identificaron materiales con diferentes aptitudes de uso.
El híbrido simple 28x4 fue seleccionado por su ACE presentando valores promisorios como doble propósito. Los híbridos simples 20x26, 4x28, 4x20 y 4x3 mostraron un comportamiento similar, mejor adaptados a la producción de forraje diferido y grano. Por último, el análisis dialélico detectó a los híbridos simples 20x4 y 3x26 como materiales mejor adaptados a la producción de forraje en estado R3-R4.
BIBLIOGRAFÍA
1. Allard R.W. (1978) Principios de la mejora genética de las plantas. Editorial Omega. Barcelona, España. [ Links ]
2. Arias S., Freddi A.J., Sánchez O., Arzadún M. (2003) Rendimiento de materia seca y composición morfológica de híbridos de sorgo y maíz para silaje de planta entera. XXVI Congr. Arg. Prod. Animal, RAPA 23 Supl. 1: 221-222. Mendoza. [ Links ]
3. CACF (2012) Cámara Argentina de Contratistas Forrajeros. [ Links ]
4. Castaldo A., Pariani A., Bulnes N., Illuminati H. (2009) Evaluación de un cultivo de maizal en estado diferido. Ciencia Veterinaria 11 (1): 37-41. [ Links ]
5. Castañon-Nájera G., Latournerie-Moreno L., Mendoza-Elos M. (2005) Macro de SAS-IML para analizar los diseños II y IV de Griffins. Universidad y Ciencia 21: 27-35. [ Links ]
6. Centeno A., Cortés E., Gallo E. (2009) Evaluación de híbridos de maíz para silo. Campaña 2008-09. Hoja de Información Técnica. UEE-INTA San Francisco-IPEM N° 222. [ Links ]
7. Centeno A., Cortés E., Ciacci M.B. (2010) Evaluación de híbridos de maíz para silo. Campaña 2009-10. Hoja de Información Técnica. UEE San Francisco. INTA. http://www.produccion-Animal.com.ar/produccion_y_manejo_reservas/reservas_ silos/148informe_maices_sileros_09-10.pdf (acceso julio 2012). [ Links ]
8. Cirilo A.G. (2004) Fecha de siembra y rendimiento en maíz. IDIA XXI 6: 122-127. [ Links ]
9. Crow J.F. (1998) 90 Years Ago: The Beginning of Hybrid Maize. Genetics 148: 923-928. [ Links ]
10. Cruz C.D. (2006) Programa Genes: Biometría. Editora UFV. Viçosa (MG), Brasil. [ Links ]
11. De la Cruz-Lázaro E., Castañón-Najera G., Brito-Manzano N., Gómez-Vásquez A., Robledo-Torres V., Lozano del Río A. (2010) Heterosis y aptitud combinatoria de poblaciones de maíz tropical FYTON (Buenos Aires) 79 (1): 11-17. [ Links ]
12. De León M., Giménez R.A., Ruolo M.S. (2011) Caracterización y variabilidad de distintos genotipos de maíz para ensilaje. EEA INTA Manfredi. http://www.inta.gob.ar/documentos/caracterizacion-y-variabilidad-de-distintos-genotipos-de-maiz-para-ensilaje/ (acceso julio 2012). [ Links ]
13. Di Nucci de Bedendo E., Firpo M.V., Martínez M. Del H. (2009) Aptitud silera de híbridos de maíz (Campaña 2008/09). INTA EEA Paraná. http://inta.gob.ar/parana/info/documentos/aptitud-silera-de-hibridos-de-maiz- campaña-2008-09/ (acceso julio 2012). [ Links ]
14. Ferraris G. (2004) Ensayo comparativo de híbridos comerciales de maíz en el área de Colón-Wheelwright. Campaña 2003/04. Proyecto Regional Agrícola. INTA Pergamino. http://www.elsitioagricola.com/articulos/ferraris/EnsayocomparativodehibridoscomercialesdeMaizenelareadeColon-Wheelwright.asp (acceso junio 2012). [ Links ]
15. Fussell B. (1999) The story of corn. North Point Press, NY, USA. [ Links ]
16. Gallais A., Pollacsek M., Huguet L. (1976) Possibilites de selection du mais en tant que plant fourragere. Ann. Amelior. Plantes 26: 591-605. [ Links ]
17. Gorostegui J. (1971) Variedades vegetales del INTA, cultivares creados e introducidos desde 1958. Serie: Colección agropecuaria del INTA 19. Buenos Aires. [ Links ]
18. Grassi E. (2009) Caracterización genética de maíz local con fines forrajeros. Tesis Maestría en Ciencias Agropecuarias. Facultad de Agronomía y Veterinaria. Universidad Nacional de Río Cuarto. Río Cuarto, Córdoba. [ Links ]
19. Grassi E., Szpiniak B., Ferreira V. (2006) Estimación de la productividad y variación de líneas endocriadas de maíz para doble propósito. Rev. Univ. Nac. Río Cuarto 26 (1-2): 3-13. [ Links ]
20. Grassi E., Reynoso L., Brun A., Ferreira A., Castillo E., Ferreira V. (2009) Líneas endocriadas de maíz: materia seca, aptitud combinatoria y agrupamiento según posibilidades de uso. Rev. Univ. Nac. de Río Cuarto 29 (1-2): 19-36. [ Links ]
21. Griffins B. (1956) Concept of general and specific combining ability in relation to diallel crossing sistems. Aust. Jour. Biol. Sci. 9: 463-493. [ Links ]
22. Guillén-De la Cruz E., De la Cruz-Lázaro E., Castañon-Nájera G., Osorio-Osorio R., Brito-Manzano N.P., Lozano-Del Río A., López-Noverola U. (2009) Aptitud combinatoria general y específica de germoplasma tropical de maíz. Tropical and Subtropical Agroecosystems 10: 101-107. [ Links ]
23. Gutiérrez del Río E., Espinoza Banda A., Palomo Gil A., Lozano García J., Antuna O. (2004) Aptitud combinatoria de híbridos de maíz para la comarca Lagunera. Rev. Fitotecnia Mexicana 27 (1): 7-11. [ Links ]
24. Hallauer A.R., Miranda Filho J.B. (1988) Quantitative genetics in maize breeding. Iowa State University Press, Ames, Iowa. U.S.A. [ Links ]
25. Iltis H., Doebley J. (1980) Taxonomy of Zea (Gramineae). II. Subespecific categories in the Zea mays complex and a generic synopsis. Amer. J. Bot. 67 (6): 994-1004. [ Links ]
26. INDEC (2005) Instituto Nacional de Estadística y Censos. Encuesta Nacional Agropecuaria. Buenos Aires. Argentina. [ Links ]
27. Lonnquist J. (1964) A modification of the ear-to-row procedure for the improvement of maize populations. Crop Sci. 4: 227-228. [ Links ]
28. Magoja J., Pischedda G. (1988) Aptitud combinatória de progênies S1 derivadas de uma población com introgresión de germoplasma silvestre. XIX Congreso Argentino de Genética. UNJujuy. 28 de agosto-1 de septiembre. p.78. [ Links ]
29. Maizar (2012) Asociación Maíz Argentino. [ Links ]
30. Maresca S., Echeverri D., Quiroz J.L. (2006) Ensayo comparativo de sorgo y maíz diferido para vacas de cría. EEA INTA Cuenca del Salado; EEA INTA Balcarce. http://www.inta.gob.ar/documentos/ensayo-comparativo-de-sorgo-y-maiz-diferido-para-vacas-de-cria.pdf (acceso abril 2012). [ Links ]
31. Paccapelo H., Molas M. (1996) Caracterización de una población de maíz forrajero con introgresión de Zea diploperennis. Revista de investigaciones Agropecuarias Vol. 27 (1): 33-38. [ Links ]
32. Paliwal R.L., Granados G., Lafitte H.R., Violic A.D., Marathee J.P. (2001) El maíz en los trópicos: mejoramiento y producción. Usos del maíz. En: Colección FAO: Producción y Protección Vegetal, N° 28. FAO, Rome (Italy). [ Links ]
33. Quemé de León J., Larios Bobadilla L., Pérez Rodas C., Soto León N. (1991) Aptitud combinatoria y predicción de híbridos de maíz (Zea mays L.) de grano amarillo a partir de cruzas dialélicas, evaluadas en dos localidades de la zona baja de Guatemala, 1989. Agronomía Mesoamericana 2: 24-30. [ Links ]
34. Reynoso L. (1996) Variabilidad genética para macollamiento y rebrote en el género Zea. MSc. Tesis Posgrado en Mejoramiento Genético Vegetal. Fac. Cs. Agrarias, UN Rosario, Santa Fe, Argentina. [ Links ]
35. Russell A.W. (1973) Desarrollo y evaluación de líneas endocriadas de maíz. Fundación Cargill, Publicación Nº 2, Buenos Aires, Argentina. [ Links ]
36. SIIA (2012) Sistema Integrado de Información Agropecuaria. Ministerio de Agricultura, Ganadería y Pesca. Argentina. [ Links ]
37. Sprague G.F., Tatum L.A. (1942) General vs. specific combining ability in single crosses of corn. J. Amer. Soc. Agron. 34: 923-932. [ Links ]
38. Steel D., Torrie J. (1988) Bioestadística: Principios y procedimientos. Editorial McGraw-Hill/Interamericana, México. [ Links ]
39. Torrecillas M., Bertoia L. (2000) Aptitud combinatoria para caracteres forrajeros en poblaciones nativas y compuestos raciales de maíz en Argentina. Invest. Agr. Prod. Prot. Veg. 15: 80-90. [ Links ]
40. Troiani H., Paccapelo H., Golberg A. (1988) Descripción botánica del híbrido interespecífico entre Zea mays x Zea diploperennis. Rev. Fac. Agronomía. UNLPam. Vol 3 Nº 2: 53-58. [ Links ]
41. Ventimiglia L., Torrens Baudrix L. (2012) Ensayo híbridos de maíz 2011/12 (9 de Julio B. Aires). EEA Pergamino. AER 9 de Julio. INTA Pergamino. http://inta.gob.ar/documentos/ensayo-hibridos-de-maiz-2011-12-9-de-julio-bs.as1/atmultidownload/file?name=Ensayo_Híbridos_maíz_2011_12_9 Julio.pdf (acceso julio 2012). [ Links ]
42. Watson S.A. (1988) Corn marketing, processing, and utilization. In: Sprague G.F., Dudley J.W. (Eds.) Corn and corn improvement. American Society of Agronomy. Madison, WI, USA, pp. 882-940. [ Links ]
