










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkBAG. Journal of basic and applied genetics
versión On-line ISSN 1852-6233
BAG, J. basic appl. genet. vol.30 no.1 Ciudad Autónoma de Buenos Aires jun. 2019
RESEARCH
Diversidad genotípica de 291 líneas de maíz de CIMMYT y caracterización fenotípica en el sur de Córdoba, Argentina
Genotypic diversity in 291 maize lines from CIMMYT and phenotypic characterization in southern Córdoba, Argentina
Rossi E.A.1,2, Ruiz M.1,2, Di Renzo M.2, Bonamico N.C.1,2
1 Instituto de Investigaciones Agrobiotecnológicas (UNRC-CONICET)
2 Facultad de Agronomía y Veterinaria, Universidad Nacional de Río Cuarto
Corresponding author: Ezequiel Rossi ezequiel_455@hotmail.com
DOI: 10.35407/bag.2019.XXX.01.04
Received: 04/26/2018
Revised version received: 03/15/2019
Accepted: 06/25/2019
RESUMEN
El maíz (Zea mays L.) posee un genoma complejo y una amplia diversidad genética. La información de caracteres fenotípicos y marcadores moleculares en su conjunto provee una mejor descripción e interpretación de la variabilidad genética. El objetivo del trabajo fue estimar la diversidad genética y caracterizar fenotípicamente un panel de 291 líneas de maíz de CIMMYT. Las líneas corresponden a ocho grupos establecidos de acuerdo a su adaptación ambiental y origen geográfico. Éstas se evaluaron fenotípicamente por medio de nueve caracteres agro-morfológicos en tres ambientes del sur de la provincia de Córdoba, Argentina. El intervalo antesis-estigma (IAE) presentó la mayor variabilidad fenotípica. El 40% de las líneas de maíz registraron un IAE menor a cinco días, indicando buena adaptación a los ambientes de evaluación. La estructura poblacional dada por los subgrupos de adaptación ambiental es sólo un factor menor que contribuye a la variabilidad fenotípica del panel estudiado. El análisis de componentes principales (ACP) permitió obtener el ordenamiento fenotípico y el genotípico, mientras que el análisis de procrustes generalizado indicó un consenso del 60% entre ambos ordenamientos para el total de líneas. El consenso entre el ordenamiento obtenido con caracteres agro-morfológicos y con marcadores moleculares indica desequilibrio de ligamiento entre los SNPs y los genes que controlan los caracteres agro-morfológicos. Los resultados muestran una amplia diversidad genética en el germoplasma evaluado, lo que sugiere que esta colección de líneas es un recurso importante para impulsar ganancias genéticas futuras en los programas de mejoramiento de maíz de Argentina.
Palabras clave: Heredabilidad en sentido amplio; Análisis multivariado; SNPs; Caracteres agro-morfológicos.
ABSTRACT
CIMMYT maize inbred lines (CMLs) are freely distributed to breeding programs around the world. Better information on phenotypic and genotypic diversity may provide guidance to breeders on how to use more efficiently the CMLs in their breeding programs. In this study a group of 291 CIMMYT maize inbred lines, was phenotyped by nine agro-morphological traits in south Córdoba, Argentina and genotyped using 18,082 SNPs. Based on the geographic information and the environmental adaptation, 291 CMLs were classified into eight subgroups. Anthesis-silking interval (IAE) was the trait with higher phenotypic diversity. A 40% of maize inbred lines, with IAE less than five days, show a good adaptation to growing conditions in south Córdoba, Argentina. The low phenotypic variation explained by environmental adaptation subgroups indicates that population structure is only a minor factor contributing to phenotypic diversity in this panel. Principal component analysis (ACP) allowed us to obtain phenotypic and genotypic orderings. Generalized procrustes analysis (APG) indicated a 60% consensus between both data type from the total panel of maize lines. In each environmental adaptation subgroup, the APG consensus was higher. This result, which might indicate linkage disequilibrium between SNPs markers and the genes controlling these agro-morphological traits, is promising and could be used as an initial tool in the identification of Quantitative Trait Loci (QTL). Information on genetic diversity, population structure and phenotypic diversity in local environments will help maize breeders to better understand how to use the current CIMMYT maize inbred lines group.
Key words: Broad-sense heritability; Multivariate analysis; SNPs; Agro-morphological traits.
INTRODUCCIÓN
El Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT) es uno de los centros más importantes de recolección, conservación y utilización de germoplasma de maíz (Yan et al., 2009). Las líneas desarrolladas en el CIMMYT constituyen una fuente pública mundial de germoplasma de maíz, las cuales se distribuyen por medio del Tratado Internacional sobre los Recursos Fitogenéticos para la Alimentación y la Agricultura (ITPGRFA) (Chen et al., 2016). Las líneas de maíz del CIMMYT (CML) se han desarrollado para un amplio número de ambientes en todo el mundo. Cada ambiente presenta diferentes temperaturas durante la estación de crecimiento, diferentes altitudes, así como otros aspectos que definen la adaptación del cultivo. El CIMMYT considera cuatro mega-ambientes principales, y cuatro subprogramas de investigación de maíz que abordan las necesidades de los productores en estas áreas. Los mega-ambientes son (i) tropicales de altitud baja, (ii) subtropical, (iii) altitud media, y (iv) ambientes de altitud alta (Xia et al., 2004., 2005).
Según el programa de mejoramiento donde se desarrollaron, estos mega-ambientes contienen líneas de distintos orígenes: México, Tailandia, Kenia, Zimbabue y Colombia (Wu et al., 2016). Comprender las relaciones entre las líneas de diferentes mega-ambientes es de utilidad para incorporar germoplasma exótico en los diferentes programas de mejoramiento. Las líneas de CIMMYT se distribuyen libremente a los programas de mejoramiento de todo el mundo, donde se cruzan con líneas adaptadas localmente. Información sobre el comportamiento de las CML en distintos ambientes permite orientar a los mejoradores sobre cómo utilizar de manera más eficiente las líneas del CIMMYT en los programas de mejoramiento (Xia et al., 2005). La variabilidad presente en el germoplasma disponible es requisito fundamental para identificar genotipos con caracteres específicos (Dinesh et al., 2016). Tradicionalmente, la diversidad genética se estimó a partir de caracteres agro-morfológicos (Yang et al., 2010), con la limitación dada por la interacción en la expresión de éstos con el ambiente (Mienie y Fourie, 2013; Tiwari et al., 2017). El análisis fenotípico sólo considera parte de la variación del germoplasma. Por ello, el análisis genotípico con marcadores moleculares es el complemento adecuado (Warburton et al., 2002). Actualmente, los marcadores SNPs (polimorfismos de un nucleótido) son ampliamente utilizados en estudios genéticos (Wu et al., 2016). Los caracteres agro-morfológicos y marcadores moleculares proveen conjuntamente una mejor descripción e interpretación de la variabilidad genética. Esto puede deberse a que la región del genoma que se explora es mayor, así como a la incorporación de caracteres de alta heredabilidad que no interaccionan con el ambiente (Demey, 2008). La heredabilidad en sentido estricto sólo cuantifica los efectos genéticos aditivos, mientras que la heredabilidad en sentido amplio comprende la suma de los efectos aditivos, de dominancia y epistáticos (Nyquist, 1991; Falconer y Mackay, 1996).
Distintas configuraciones se obtienen a partir de caracteres agro-morfológicos y marcadores moleculares para un mismo grupo de genotipos (Balzarini et al., 2008). En esta situación el análisis procrustes generalizado (APG) resulta adecuado como método para analizar la congruencia entre las configuraciones individuales (Bruno y Balzarini, 2010). El APG fue aplicado por Bramardi et al. (2005) para investigar relaciones entre genotipos de Cucumis sativus L. evaluados con caracteres agronómicos y caracterizados con marcadores RAPD, así como también por Hernández et al. (2010) para consensuar el ordenamiento obtenido mediante marcadores moleculares y caracteres agromorfológicos en maíz. El presente estudio se realizó con el objetivo de: i) estimar la diversidad genética de un panel de líneas de maíz de CIMMYT mediante marcadores moleculares SNPs; ii) caracterizar fenotípicamente el panel de líneas de maíz del CIMMYT mediante caracteres agromorfológicos en el sur de Córdoba, Argentina; iii) evaluar los efectos de la estructura de la población sobre el fenotipo; iv) evaluar el consenso entre el ordenamiento fenotípico obtenido mediante caracteres agromorfológicos y el ordenamiento genotípico obtenido a partir de marcadores moleculares.
MATERIALES Y MÉTODOS
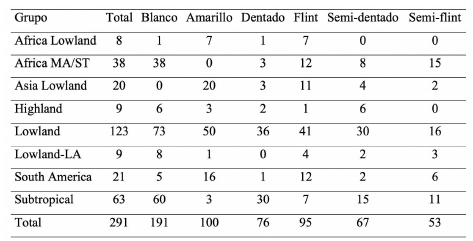
Material vegetal y ensayo de campo
El material vegetal estuvo constituido por 291 líneas de maíz desarrolladas y provistas por el CIMMYT (http:// www.cimmyt.org/seed-request). La zona de adaptación o el programa de mejoramiento de origen permitieron diferenciar ocho grupos: tropical de altitud baja (Lowland), subtropical (Subtropical), subtropical de África (Africa MA/ST), América del Sur (South America), altitud alta (Highland), altitud baja de Asia (Asia Lowland), de África (Africa Lowland) y de América Latina (Lowland-LA)(http://hdl.handle.net/11529/10246). En la Tabla 1 se presenta un resumen de la diversidad fenotípica para color y textura de grano en cada uno de estos grupos. En los ciclos agrícolas 2015/2016 y 2016/2017 se realizaron tres ensayos a campo con la finalidad de evaluar fenotípicamente las líneas de maíz. En el primer ciclo de evaluación se estableció un ensayo en el campo experimental de la Facultad de Agronomía y Veterinaria de la Universidad Nacional de Río Cuarto (CAMDOCEX, FAV-UNRC, 33º 06′ S; 64º 17′ O). En el segundo ciclo se realizaron dos ensayos, uno en la localidad de Río Cuarto (CAMDOCEX, FAV-UNRC) y otro en la localidad de Chaján (33° 33′ S 65° 00′ O). El diseño utilizado fue en bloques aumentados parcialmente repetidos (Williams et al., 2011), con un 25% de los genotipos, elegidos al azar, con tres repeticiones y el resto de los genotipos con sólo una repetición. Cada genotipo se estableció en parcelas de un surco de 3,0 m de largo espaciados a 0,52 m. Las diferentes combinaciones año-localidad constituyeron los tres ambientes de evaluación: Río Cuarto, ciclo agrícola 2015/2016 (E1); Río Cuarto, ciclo agrícola 2016/2017 (E2) y Chaján, ciclo agrícola 2016/2017 (E3).
Tabla 1. Número de genotipos por grupo de adaptación, color de grano (blanco y amarillo) y textura de grano (dentado, flint, semi-dentado y semi-flint) de las 291 líneas de maíz de CIMMYT.
Evaluación fenotípica
Las líneas de maíz se evaluaron mediante nueve caracteres agro-morfológicos. Los caracteres fueron altura de planta (AP), altura de espiga (AE), largo de la panoja (LP), ángulo de inserción de la primera hoja por encima de la espiga principal (AH1), ángulo de inserción de la segunda hoja por encima de la espiga principal (AH2), número de raquis por panoja (RP), días a floración masculina (DFM), días a floración femenina (DFF). Estos se definieron cuando el 50% de las plantas de cada surco alcanzó cada uno de los estadios. A partir de los DFM y los DFF se estimó el carácter intervalo antesis-estigma (IAE). Los caracteres AP, AE, LP, AH1, AH2, y RP se midieron en cinco plantas para obtener el valor medio de cada surco. Los caracteres DFM, DFF e IAE se midieron sólo en los ambientes E1 y E2.
Caracterización genotípica
La caracterización de las líneas de maíz con marcadores moleculares realizada por Chen et al. (2016) es de disponibilidad pública (http://data.cimmyt.org/dvn/dv). La información disponible consta de 18.082 marcadores moleculares SNPs, los cuales fueron utilizados en el presente estudio para el panel total de líneas de maíz y cada uno de los grupos de adaptación.
Análisis estadístico
Fenotípico
La normalidad de los caracteres AP, AE, LP, AH1, AH2, RP, DFM, DFF y del IAE se comprobó con el test de Shapiro-Wilks modificado y luego se realizó un Análisis de la Varianza (ANOVA) a través de ambientes según el siguiente modelo estadístico:
![]()
donde es la observación de cada uno de los caracteres medidos, μ es la media general, es el efecto del i-ésimo genotipo, es el efecto del j-ésimo ambiente, es el efecto del k-ésimo bloque dentro de cada ambiente, es el efecto de la interacción genotipo-ambiente y es un término de error aleatorio asociado a la observación. La aplicación de este modelo permitió obtener las medias de cada carácter a través de ambientes y los efectos de la interacción genotipo-ambiente (GE). Los componentes de varianza de genotipo, ambiente, interacción GE y del error residual se estimaron a partir de los cuadrados medios obtenidos del ANOVA. Los genotipos que presentaron repeticiones permiten obtener una estimación del error aleatorio, mientras que la estimación de la varianza de la interacción genotipoambiente (GE) y la varianza genotípica se realizó con la totalidad de los genotipos (Bernardo, 2002). Los componentes de varianza fueron utilizados para estimar la heredabilidad en sentido amplio sobre la media de cada parcela según Holland et al. (2003). La estimación de la proporción de la variación fenotípica explicada por la estructura de la población se realizó mediante una regresión lineal múltiple. Para cada carácter, se utilizaron las medias a través de ambientes como variable respuesta y la matriz de estructura poblacional de acuerdo a la zona de adaptación, como variable predictora.
Además, para cada carácter se evaluó la diversidad fenotípica del panel de líneas de maíz mediante la estimación del índice de Shannon-Weaver (S-W). Para esto, cada uno de los caracteres se dividió en 10 categorías utilizando el valor medio (M) y el desvío estándar (DE). En la categoría 1 se incluyeron los valores fenotípicos menores a M - 2DE, mientras que la categoría 10 incluyó a los valores mayores a M + 2DE. El intervalo entre cada una de las categorías fue de 0,5DE. Los valores medios a través de ambientes de los nueve caracteres agro-morfológicos se analizaron con un enfoque multivariado, el Análisis de Componentes Principales (ACP), para explorar las relaciones entre caracteres, entre genotipos y entre caracteres y genotipos. El ACP se realizó para el panel de 291 líneas de maíz, así como también para cada uno de los grupos de adaptación. Los análisis se realizaron mediante el programa Infostat (Di Rienzo et al., 2018).
Genotípico
El ordenamiento de las líneas basado en la caracterización molecular se realizó a partir de un Análisis de Componentes Principales (ACP) con los 18.082 SNPs. Las dos primeras componentes principales permitieron realizar el ordenamiento o visualización de los genotipos en un plano. El ACP con los 18.082 SNPs también se realizó para el panel total de líneas y para cada uno de los grupos. Los análisis se realizaron mediante el programa TASSEL (Bradbury et al., 2007).
Análisis procrustes generalizado
El consenso entre el ordenamiento fenotípico y el ordenamiento genotípico se estimó mediante un Análisis Procrustes Generalizado (APG) (Gower, 1975). Este se realizó a partir de las componentes principales obtenidas del ordenamiento fenotípico y genotípico, respectivamente. Las componentes principales fenotípicas seleccionadas fueron las que tenían un auto valor igual o mayor a uno. Mientras que, para las componentes principales genotípicas se seleccionó el número de componentes donde se produce la mayor caída en la variación explicada. El APG se realizó con la totalidad de las líneas y para cada uno de los grupos de adaptación.
RESULTADOS
La evaluación fenotípica del panel de líneas de maíz de CIMMYT se realizó mediante la medición de nueve caracteres agro-morfológicos. Los caracteres se midieron en ambientes del sur de Córdoba, Argentina y se observó un amplio rango de variación para cada uno de ellos (Cuadro 2). De acuerdo al cociente entre los valores máximos y mínimos, el IAE fue el carácter que presentó mayor variabilidad fenotípica, el cual varió entre 1 y 22 días. El número de RP fue el otro carácter que presentó amplia variación con una media de 17,6 (±6,6) con un mínimo de 5 raquis y un máximo de 47 raquis. Mientras que los caracteres AP, DFM y DFF fueron los que mostraron menor variabilidad fenotípica con medias de 172,4 (±19), 90,8 (±9,1) y 96,3 (±10,4), respectivamente. El índice de Shannon-Weaver presentó valores entre 1,92, para el IAE, y 2,09 para AP, en tanto que el resto de los caracteres presentó valores intermedios. Al estimar el índice de Shannon-Weaver se pudo observar en base a las frecuencias de cada categoría que el 40% de las líneas de maíz registraron un IAE menor a 5 días (datos no mostrados).
Tabla 2. Estadística descriptiva, índice de Shannon-Weaver, heredabilidad y porcentaje de variación fenotípica explicada por los grupos de adaptación para los nueve caracteres agro-morfológicos medidos en las 291 líneas de maíz de CIMMYT en los ambientes Río Cuarto, ciclo agrícola 2015/2016 (E1); Río Cuarto, ciclo agrícola 2016/2017 (E2) y Chaján, ciclo agrícola 2016/2017 (E3).
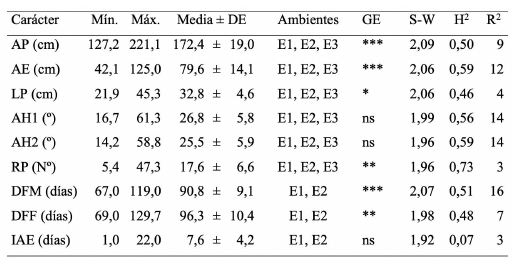
El ANOVA a través de ambientes permitió observar que la interacción GE fue significativa en todos los caracteres excepto para AH1, AH2 e IAE. La heredabilidad en sentido amplio presentó valores intermedios para la mayoría de los caracteres. El carácter RP presentó el mayor valor de heredabilidad (0,73), y el carácter IAE el menor valor (0,07). El porcentaje de variabilidad fenotípica explicada por los distintos grupos de adaptación ambiental fue bajo para los nueve caracteres. Sin embargo, en los caracteres DFM y AH la presencia de grupos explicó el 16% y el 14% de la variación fenotípica, respectivamente. El análisis de componentes principales (ACP) redujo la dimensión de la matriz de información fenotípica de los nueve caracteres agro-morfológicos medidos. Este análisis permitió obtener cinco componentes principales (CP) significativas que explicaron el 88% de la variabilidad total. La Figura 1 presenta el gráfico generado a partir de las dos primeras CP del ACP que explican el 47% de la variabilidad. En el gráfico se observa que los distintos grupos de adaptación tienen poca influencia en la variación fenotípica de los nueve caracteres medidos ya que no existe separación entre los genotipos pertenecientes a los distintos grupos.
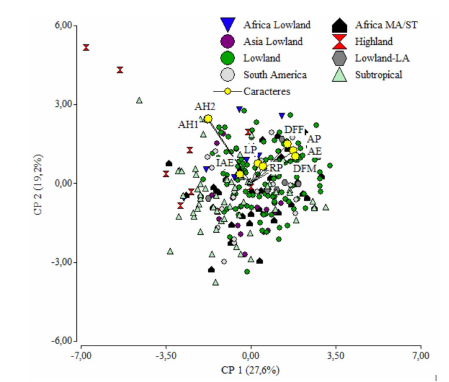
Figura 1. Gráfico biplot a partir de las dos primeras componentes principales obtenidas del análisis de componentes principales con los nueve caracteres agro-morfológicos medidos en las 291 líneas de maíz del CIMMYT evaluadas en tres ambientes del sur de la provincia de Córdoba, Argentina, durante los ciclos agrícolas 2015/2016 y 2016/2017.
AP: altura de planta; AE: altura de espiga; LP: largo de panoja; AH1: ángulo de inserción de la primera hoja por encima de la espiga principal; AH2: ángulo de inserción de la segunda hoja por encima de la espiga principal; RP: número de raquis por panoja; DFM: días a floración masculina; DFF: días a floración femenina; IAE: intervalo antesis-estigma.
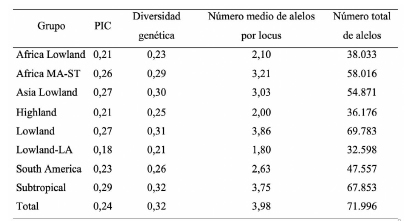
Los 18.082 SNPs permitieron estimar el contenido de información polimórfica (PIC) y la diversidad genética presente en el panel de líneas de maíz de CIMMYT. El PIC y la diversidad genética oscilaron entre los distintos grupos. El grupo de altitudes bajas de Latinoamérica (Lowland-LA) registró el menor valor y el grupo Subtropical el mayor valor. El número total de alelos para el panel de líneas fue de 71.996 con una media de 3,98 alelos por locus (Cuadro 3).
Tabla 3. Medidas de resumen relacionadas con la diversidad genética para las 291 líneas de maíz de CIMMYT y para cada uno de los grupos de adaptación, estimadas con 18.082 SNPs.
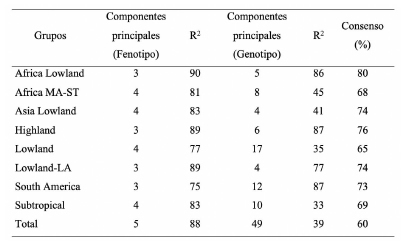
El análisis de componentes principales del panel de líneas, a partir de los 18.082 marcadores SNPs, permitió obtener 49 componentes principales que explican el 39% de la variabilidad. El gráfico obtenido con las dos primeras CP se muestra en la Figura 2. En general líneas pertenecientes a un mismo grupo de adaptación ambiental presentan menores distancias. El grupo de líneas tropicales de altitud baja (Lowland) que es el de mayor número de líneas presenta una amplia variación entre sus genotipos, superponiéndose con el resto de los grupos. Al analizar cada uno de los grupos de adaptación, el ACP realizado con los caracteres agro-morfológicos permitió obtener entre tres y cuatro CP significativas, que explicaron entre 75% y 90% de la variabilidad fenotípica. Mientras que en el ACP realizado a partir de los 18.082 SNPs, entre 4 y 17 CP fueron significativas. Estas explicaron entre 33% y 87% de la variabilidad genotípica presente (Tabla 4). El porcentaje de consenso para el panel de líneas fue de 60%, mientras que para cada uno de los grupos este valor de consenso fue mayor. Así, en el grupo de líneas correspondientes a altitudes bajas de África (Africa Lowland) se observó el mayor valor de consenso (80%) entre ambos ordenamientos.
Figura 2. Gráfico de las dos primeras componentes principales obtenidas del análisis de componentes principales a partir de 18.082 SNPs en 291 líneas de maíz del CIMMYT.
Tabla 4. Número de componentes principales significativos y porcentaje de variación, fenotípica y genotípica, explicada por esas componentes. Consenso obtenido mediante el Análisis de Procrustes Generalizado (APG) a partir de información proveniente de caracteres agro-morfológicos y de marcadores moleculares SNPs de las 291 líneas de maíz de CIMMYT.
DISCUSIÓN
En los programas de mejoramiento genético de maíz a nivel mundial se aplicó selección directa para rendimiento en grano, lo cual llevó a una selección indirecta para caracteres relacionados con el rendimiento como ángulo de hoja e intervalo antesis-estigma. Las hojas erectas permiten una mejor intercepción de la luz ante el incremento de la densidad de plantas, propio del cultivo moderno. Mientras que, intervalos antesis-estigma breves indican que la espiga presentó un ritmo de crecimiento normal sin obstáculos (Duvick, 2004). El grupo de líneas de maíz utilizadas en este estudio mostró estar sometidas a varias generaciones de mejoramiento ya que en general presentan hojas erectas e intervalos antesis-estigma menores a cinco días, lo que indicaría adecuada adaptación a los ambientes de evaluación de este estudio. La estructura de la población, los ocho grupos en este panel de líneas, explicó en promedio un 9% de la variación fenotípica para los nueve caracteres agromorfológicos medidos. Esto sugiere que los grupos de adaptación son un factor con una contribución menor a la variación fenotípica en este panel de líneas. Sin embargo, los efectos variaron según el carácter y se observó mayor influencia de la estructura poblacional para los caracteres días a floración masculina y ángulo de inserción de la primera y segunda hoja por encima de la espiga principal. El conocimiento de la variación fenotípica y genotípica existente entre diversos caracteres agro-morfológicos, así como de la heredabilidad de éstos es importante en los programas de mejoramiento (Beyene et al., 2005). Las líneas de maíz del CIMMYT evaluadas en este estudio presentan amplia variabilidad para los caracteres agro-morfológicos medidos en el sur de la provincia de Córdoba, Argentina. Los valores medios a altos estimados de heredabilidad en sentido amplio en este estudio indican la posibilidad de utilizar las líneas de maíz mejor adaptadas en programas de mejoramiento locales. Éstas pueden incorporarse para el mejoramiento poblacional, para el desarrollo de genotipos híbridos, variedades de polinización abierta o poblaciones de mapeo. La diversidad genética y el contenido de información polimórfica fueron similares a los informados por Wu et al. (2016), quienes caracterizaron la totalidad de líneas de maíz de CIMMYT mediante un amplio set de marcadores moleculares SNPs obtenidos a través de genotipado por secuenciación. Los resultados del presente estudio indican que el panel de líneas caracterizadas fenotípicamente en el sur de Córdoba es una muestra representativa del conjunto de líneas de maíz público de CIMMYT.
La estadística multivariada es ampliamente usada para describir y analizar observaciones multidimensionales obtenidas al compilar información sobre diferentes caracteres en distintos genotipos (Balzarini et al., 2008). Estudios previos informan el uso de análisis de procrustes generalizado para consensuar el ordenamiento obtenido mediante marcadores moleculares y el ordenamiento obtenido mediante caracteres agro-morfológicos. Bramardi et al. (2005) aplicaron APG para investigar relaciones entre genotipos de Cucumis sativus L. evaluados mediante caracteres agronómicos y caracterizados con marcadores RAPD. Hernández et al. (2010) informan un 67% de consenso entre los ordenamientos individuales obtenidos con 14 caracteres fenotípicos y 40 marcadores moleculares de 14 líneas de maíz evaluadas fenotípicamente en Venezuela. En el presente estudio, la matriz de caracteres agro-morfológicos se redujo a cinco componentes principales que explicaron el 88% de la variabilidad total y la matriz de información genotípica se redujo a 49 componentes principales que explicaron el 39% de la variabilidad. El análisis de procrustes generalizado permitió obtener un valor moderado de consenso (60%) entre el ordenamiento de los genotipos evaluados mediante caracteres agromorfológicos y marcadores moleculares. Este valor de consenso fue mayor al analizar cada grupo por separado, así en el grupo de líneas de baja altitud de África (Africa Lowland) se observó un 80% de consenso. El consenso entre ordenamientos fenotípicos y genotípicos podría ser usado como una herramienta inicial promisoria en la identificación de loci de caracteres cuantitativos (QTL), con la ventaja de poder realizarse para más de un carácter simultáneamente (Demey, 2008). La correlación entre la diversidad agro-morfológica y molecular indicaría la existencia de desequilibrio de ligamiento entre genes que controlan los caracteres agro-morfológicos y los marcadores moleculares (Baranger et al., 2004). En este estudio, el valor de consenso observado entre los ordenamientos individuales, fenotípico y genotípico, indica la presencia de desequilibrio de ligamiento entre loci que controlan los caracteres agro-morfológicos medidos y los marcadores SNPs empleados. Esto pone de manifiesto el potencial de este grupo de líneas de maíz para identificar QTL de caracteres de interés agronómico mediante la implementación de estudios de mapeo por asociación. Además, brinda información sobre la estructura de la población basada en los grupos de adaptación ambiental, la cual debería ser considerada en futuros estudios de mapeo por asociación. Un panel de mapeo por asociación debe abarcar la máxima diversidad fenotípica y molecular que pueda ser estimada de manera confiable en un ambiente común (Flint-Garcia et al., 2005). El uso de germoplasma con caracteres favorables es el procedimiento más económico y ambientalmente sustentable para lograr incrementos y estabilidad en la producción de maíz (Di Renzo et al., 2002). En la práctica, los programas de mejoramiento de clima templado intercambian germoplasma con programas de mejoramiento subtropical o de mediana altitud (Wu et al., 2016). Los resultados del presente estudio mostraron una amplia diversidad genética en el germoplasma evaluado, el cual en su mayoría corresponde a programas de origen tropical. Esta colección de 291 líneas de maíz de CIMMYT constituye un valioso recurso para impulsar ganancias genéticas futuras en los programas de mejoramiento de maíz en Argentina.
BIBLIOGRAFÍA
1. Balzarini M., Gonzalez L., Tablada M., Casanoves F., Di Rienzo J., Robledo C. (2008) Infostat. Manual del Usuario, Editorial Brujas, Córdoba, Argentina. [ Links ]
2. Baranger A., Aubert G., Arnau G., Lainé A.L., Deniot G., Potier J., Weinachter C., Lejeune-Hénaut I., Lallemand J., Burstin J. (2004) Genetic diversity within Pisum sativum using protein- and PCR-based markers. Theor. Appl. Genet. 108: 1309-1321. https://doi.org/10.1007/s00122-003-1540-5 [ Links ]
3. Bernardo R. (2002) Breeding for Quantitative Traits in Plants. Stemma Press,
4. Beyene T., Botha A.M., Myburg A.A. (2005) Phenotypic diversity for morphological and agronomic traits in traditional ethiopian highland maize accessions. South African J. Plant Soil. 22: 100-105. https://doi.org/10.1080/02571862.2005.10634689 [ Links ]
5. Bradbury P., Zhang Z., Kroon D., Casstevens T., Ramdoss Y., Buckler E. (2007) TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 23: 2633-2635. [ Links ]
6. Bramardi S.J., Bernet G.P., Asíns M.J., Carbonell E.A. (2005) Simultaneous agronomic and molecular characterization of genotypes via the generalised procrustes analysis: An application to cucumber. Crop Sci. 45: 1603-1609. https://doi.org/10.2135/cropsci2004.0633 [ Links ]
7. Bruno C., Balzarini M. (2010) Ordenaciones de material genético a partir de información multidimensional. Rev. FCA UNCuyo. 42: 183-200. [ Links ]
8. Chen J., Zavala C., Ortega N., Petroli C., Franco J., Burgueño J., Costich D., Hearne, S.J. (2016) The development of quality control genotyping approaches: A case study using elite maize lines. PLoS ONE. 11: 1-17 https://doi.org/10.1371/journal.pone.0157236 [ Links ]
9. Demey J. (2008) Diversidad genética en bancos de germoplasma: un enfoque biplot. Tesis de doctorado. Departamento de estadística. Universidad de Salamanca, España. [ Links ]
10. Dinesh A., Patil A., Zaidi P.H., Kuchanur P.H., Vinayan M.T., Seetharam K. (2016) Genetic diversity, linkage disequilibrium and population structure among CIMMYT maize inbred lines, selected for heat tolerance study. Maydica 61. [ Links ]
11. Di Renzo M.A., Bonamico N.C., Díaz D.D., Salerno J.C., Ibañez M.A., Gesumaría J.J. (2002) Inheritance of resistance to Mal de Río Cuarto (MRC) disease in Zea mays (L.). J. Agr. Sci. 139: 47-53. [ Links ]
12. Di Rienzo J., Casanoves F., Balzarini M., Gonzalez L., Tablada M., Robledo C. (2018) InfoStat. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. URL http://www.infostat.com.ar [ Links ]
13. Duvick D. (2004) Long-term selection in a commercial hybrid maize breeding program. Plant Breed. In: J. Janick (eds.) John Wiley & Sons, Inc., Hoboken, NJ, pp. 109-151. [ Links ]
14. Falconer D.S., Mackay T.F.C. (1996) An Introduction to Quantitative Genetics, Ed. 4. Prentice Hall, London. [ Links ]
15. Flint-Garcia S.A., Thuillet A.C., Yu J., Pressoir G., Romero S.M., Mitchel S.E., Doebley J., Kresovich S., Goodman M.M., Buckler E.S. (2005) Maize association population: a high-resolution platform for quantitative trait locus dissection. Plant J. 44: 1054-1064. [ Links ]
16. Gower J. (1975) Generalized procrustes analysis. Psychometrika 40: 33-51. [ Links ]
17. Hernández A., San Vicente F., Figueroa-Ruiz R. (2010) Evaluación y caracterización de líneas parentales de híbridos de maíz (Zea mays L.) en tres ambientes de Venezuela. Interciencia 35: 290-298. [ Links ]
18. Holland J.B., Nyquist W.E., Cervantes-Martínez C.T. (2003) Estimating and Interpreting Heritability for Plant Breeding: An Update. Plant Breed. Reviews. https://doi.org/10.1002/9780470650202.ch2 [ Links ]
19. Mienie C.M.S., Fourie A.P. (2013) Genetic diversity in South African maize (Zea mays L.) genotypes as determined with microsatellite markers. African J. Biotechnol. 12: 123-133. https://doi.org/10.5897/AJB12.2282 [ Links ]
20. Nyquist W.E. (1991) Estimation of heritability and prediction of selection response in plant populations. Crit. Rev. Plant Sci. 10: 235-322. [ Links ]
21. Tiwari A., Vajire D., Thakare K., Solunke R., Panche A. (2017) The Possibilities of the Prediction of Heterosis in Elite Lines of Onion Based on the Assessment of Genetic Diversity. EC Microbiology 10 (5): 211-219. [ Links ]
22. Warburton M.L., Xianchun X., Crossa J., Franco J., Melchinger A.E., Frisch M., Bohn M., Hoisington D. (2002) Genetic characterization of CIMMYT inbred maize lines and open pollinated populations using large scale fingerprinting methods. Crop Sci. 42: 1832-1840. https://doi.org/10.2135/cropsci2002.1832 [ Links ]
23. Williams E., Piepho H.P., Whitaker D. (2011) Augmented p-rep designs. Biometrical J. 53: 19-27. https://doi.org/10.1002/bimj.201000102 [ Links ]
24. Wu Y., San Vicente F., Huang K., Dhliwayo T., Costich D. E., Semagn K., Sudha N., Olsen M., Prasanna B.M., Zhang X., Babu R. (2016) Molecular characterization of CIMMYT maize inbred lines with genotyping-bysequencing SNPs. Theor. Appl. Genet. 129: 753-765. https://doi.org/10.1007/s00122-016-2664-8 [ Links ]
25. Yan J., Shah T., Warburton M.L., Buckler E.S., McMullen M.D., Crouch J. (2009) Genetic characterization and linkage disequilibrium estimation of a global maize collection using SNP markers. PLoS One 4. https://doi.org/10.1371/journal.pone.0008451 [ Links ]
26. Yang X., Yan J., Shah T., Warburton M.L., Li Q., Li L., Gao Y., Chai Y., Fu Z., Zhou Y., Xu S., Bai G., Meng Y., Zheng Y., Li J. (2010) Genetic analysis and characterization of a new maize association mapping panel for quantitative trait loci dissection. Theor. Appl. Genet. 121: 417-431. https://doi.org/10.1007/s00122-010-1320-y [ Links ]
27. Xia X.C., Reif J.C., Hoisington D.A., Melchinger A.E., Frisch M., Warburton M.L. (2004) Genetic diversity among CIMMYT maize inbred lines investigated with SSR markers: I. Lowland tropical maize. Crop Sci. 44: 2230-2237. [ Links ] [ Links ]

