











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCTION
Polyploidy is a common phenomenon across numerous eukaryotic taxa (Soltis et al., 2014; Marchant et al., 2016) and is believed to play a significant role in the evolution of vascular plants (Otto and Whitton, 2000; Levin, 2002). As many as 70-80% of Angiosperms, including many crop species, and up to 95% of Pteridophytes have a polyploid origin (Bennet, 2004; Gregory and Mable, 2005; Otto, 2007). In addition, owing to the role in the origin of evolutionary novelties and the maintenance of diversity in plant populations, polyploidy was recognized as an integral component of the ecological and evolutionary dynamics of plant species populations. In this regard, different theoretical and experimental efforts to examine the evolutionary significance of polyploidy have focused on the processes responsible for the origin of polyploid plants and the conditions favouring their establishment and persistence (Thompson and Lumaret, 1992).
Spontaneous doubling of somatic chromosomes (zygotic or meristematic) was long considered the predominant mode of polyploid origin in flowering plants (Winge, 1917). However, it became soon clear that 2n gametes detected in many plant’ species (Karpechenko, 1927; Darlington, 1937) are involved in polyploid origin and, that sexual polyploidization is the driving force giving rise to polyploid plant species (Harlan and de Wet, 1975; Camadro, 1986; Thompson and Lumaret, 1992; Bretagnolle and Thompson, 1995; Ramsey and Schemske, 1998; Brownfield and Köhler, 2011). Particularly, autotetraploids may be formed in single steps through the union of two 2n gametes produced by diploids (bilateral polyploidization) or, alternatively, may arise in two steps from matings involving viable triploids (triploid bridge hypothesis), themselves the result of the union of n and 2n gametes produced by diploids (unilateral polyploidization) (Harlan and de Wet, 1975).
On the other hand, the process of neopolyploids establishment in diploid populations may be limited by the difficulty of mating with other plants of the same ploidy level, the unviability of triploid hybrids, the viability and fertility of polyploids relative to diploids, and the potential for genetic swamping of the more frequent cytotype (minority cytotype exclusion) (Levin, 1975; Fowler and Levin, 1984; Bever and Felber, 1998; Erilova et al., 2009; Chrtek et al., 2017). Self-pollinating plants and individuals with multiple breeding opportunities throughout life can more easily overcome the difficulties associated with intercytotypic mating (Rodriguez, 1996; Bever and Felber, 1998; Ramsey and Shemske, 1998; Husband, 2000). However, in outbreeding plants and if pollination is random, neopolyploids will be at a disadvantage, as most of their megaspores will be fertilised by n microspores of diploids forming triploids, whereas most of the relatively abundant diploid megaspores will be conveniently fertilised. Consequently, a neopolyploid is likely to be excluded from a diploid population (Levin, 1975).
Besides, polyploidy can potentially contribute to the acquisition of new morphological, genetic and/or physiological traits, which may improve the competitive ability, fitness or ecological tolerance of polyploids compared to the diploid parents. These events, which have occurred on time scales ranging from ancient to contemporary, are assumed to be of fundamental importance for plant adaptation and range expansion (Levin, 1983; Udall and Wendel, 2006; Van de Peer et al., 2021). Thus, polyploids may have a distinct or peripheral distribution compared to the parental diploids along climatic or environmental gradients (Levin, 2002). Such differences in geographical ranges between cytotypes may reflect historical patterns of colonisation or genetic differentiation that have occurred in association with or subsequent to polyploid formation (Levin, 1983). Alternatively, the spatial patterns of cytotypes may be explained through frequency-dependent production of hybrids with low frequency (Levin, 1975). Despite the evolutionary significance of polyploidy, many important questions about the mechanisms by which polyploids are formed and become established in natural populations remain unanswered (Soltis et al., 2010; Castro et al., 2018).
Turnera sidoides L. (Passifloraceae, Turneroideae) is one of the few well-studied autopolyploid complexes in South America (Fernández, 1987; Solís Neffa, 2000; Solís Neffa and Fernández, 2001, 2002; Elías et al., 2011, Kovalsky and Solís Neffa, 2012, 2015, 2016; Kovalsky et al., 2014, 2018; Roggero Luque et al., 2015, 2017; Solís Neffa et al., 2022). Because of the prominent role played by polyploidy within T. sidoides, in order to understand the evolutionary significance of autopolyploidy, an investigation is in progress in this complex using integrative approaches combining cytogenetics, together with genetic, morphological, biogeographic and environmental data. As a part of this study, we selected this autopolyploid complex as an ideal biological model system to investigate the evolutionary dynamics of polyploids in natural populations. In this paper we synthesise the results of more than 20 years of research on this topic.
TURNERA SIDOIDES
Turnera sidoides (Figure 1) is a complex of perennial, rhizomatous herbs that is naturally distributed in southern Bolivia and Brazil, southwestern Paraguay, Uruguay and Argentina, reaching up to 39º S (Arbo, 1985; Solís Neffa, 2000). It grows in a wide variety of habitats ranging from mountain regions (up to 2700 m a.s.l) to the sea level and encompassing different climatic regimes (Arbo, 1985; Solís Neffa, 2000, 2010).

Figure 1 Turnera sidoides complex. A) T. sidoides subsp. carnea. B) T. sidoides subsp. holosericea. C) T. sidoides subsp. integrifolia. D)-E) T. sidoides subsp. pinnatifida D) Andino morphotype. E) Serrano morphotype. F) T. sidoides subsp. sidoides.
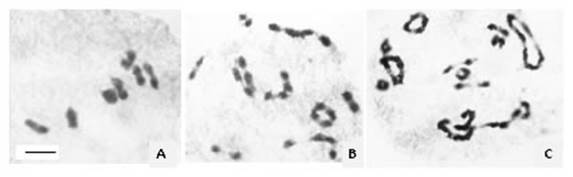
Figure 2 Meiotic chromosomes of Turnera sidoides. A) Diploid cytotype, 7 II. B) Tetraploid cytotype, 4II + 5IV. C) Hexaploid cytotype, 8 II + 5 IV + 1 VI. Bar= 5μm.
Turnera sidoides is outbreeder because of distyly and genetic self-incompatibility, being legitimate crossings those between short-styled (S) and long-styled (L) morphs, S x L or L x S (Arbo, 1985; Solís Neffa, 2000). Pollination is carried out by butterflies, small wasps and bees, and seeds are dispersed by gravity and ants. Discrete populations, ranging from less than ten to hundreds of plants, are separated from each other by a few to several kilometres (Solís Neffa, 2000).
Five subspecies were recognized on the basis of geographical distribution and the variability of some morphological features (Arbo, 1985): T. sidoides subsp. carnea (Cambess.) Arbo, T. sidoides subsp. holosericea (Urb.) Arbo, T. sidoides subsp. integrifolia (Griseb.) Arbo, T. sidoides subsp. pinnatifida (Juss. ex Poir.) Arbo, and T. sidoides subsp. sidoides. Besides, populations of T. sidoides subsp. carnea were grouped into two morphotypes (grandense and mercedeño) according to the leaf consistency, the colour of the flowers and geographical distribution (Moreno et al., 2021); while in T. sidoides subsp. pinnatifida five morphotypes (andino, chaqueño, mesopotamico, pampeano and serrano) have been distinguished based on the degree of incision of the leaf blade, the colour of the flowers and geographical distribution (Solís Neffa, 2010).
In addition to the morphological variability, polyploidy is the most outstanding feature of T. sidoides complex, with ploidy levels ranging from diploid (2n=2x=14) to octoploid (2n=8x=56), all based on x=7. Polyploid series within each subspecies/morphotype were reported (Fernández, 1987; Solís Neffa, 2000; Solís Neffa and Fernández, 2001; Solís Neffa et al., 2004; Speranza et al., 2007; Elías et al., 2011; Moreno et al., 2015, 2021; Solís Neffa et al., 2022). From the study of meiosis (Figure 2) and pollen viability it was suggested that polyploids of T. sidoides would have an autopolyploid origin (Fernández, 1987; Solís Neffa, 2000). These observations were supported by the results obtained from the analysis of the effect of polyploidy in some morphological, cytological, and biochemical traits (Solís Neffa, 2000; Solís Neffa and Fernández, 2002; Roggero Luque et al., 2022; Solís Neffa et al., 2003).
MECHANISMS OF POLYPLOID ORIGIN IN TURNERA SIDOIDES
Evidence of 2n microspore production in natural diploid populations of T. sidoides
Aiming to contribute to the understanding of the mechanisms involved in the origin and the establishment of polyploids, we firstly investigate the occurrence and frequency of unreduced microspores in diploids of T. sidoides. Early experimental studies provided the first evidences for the production of 2n and 4n microspores by analysing the size range of pollen and the constitution of the sporads in diploid plants (Panseri et al., 2008). Different facts supported that the giant pollen grains observed in the complex correspond to unreduced gametes. First, the volume of the giant grains relative to the mean pollen size of the population studied was comprised in the range proposed by different authors as typical of unreduced gametes (Darlington, 1965; Orjeda et al., 1990; Ramsey and Schemske, 1998). The existence of unreduced microspores in some plants was further confirmed by a particular bimodal distribution of pollen volume. Additionally, the presence of triads and dyads in plants with giant pollen indicates that 2n gametes certainly occurred and that the formation of 2n gametes would be expected. Moreover, the finding of monads suggested that “jumbo” (4n) gametes may also be formed. The mean frequency of unreduced gametes observed in such populations was in agreement with the estimations of the production of unreduced gametes for non-hybrid plants (Ramsey and Schemske, 1998; Ramsey, 2007).
Then, to further contribute to the understanding of the mode of polyploid formation in T. sidoides, we selected natural diploid populations that had been experimentally identified for its high production of 2n gametes (Panseri et al., 2008). In these natural populations we studied the occurrence of unreduced microspores and estimated the frequency of their production by analysing the pollen size distribution and the constitution of the sporads (Figure 3) from samples obtained in the field (Kovalsky, 2012; Kovalsky and Solís Neffa, 2012). Some of these plants were then grown under the same soil and irrigation conditions at the greenhouse of the Instituto de Botánica del Nordeste for, at least, one year. The exhaustive study on such natural populations of T. sidoides (Kovalsky and Solís Neffa, 2012) revealed that some plants were more capable of inducing unreduced microspores production than others. The percentage of 2n gamete producer plants detected (~26%) evidenced that the production of unreduced microspores was not uncommon in diploid populations of T. sidoides. These studies also showed that, within populations, S and L plants differed in their ability to produce 2n microspores and, that the frequency of 2n gametes production was highly variable. Such variation in the frequencies of unreduced microspores production observed in T. sidoides was attributed to different factors. Considering that only some individuals produced unreduced microspores, the frequency of 2n microspores production varied among producer plants and that, producer plants identified in the field were consistently classified as producers in the greenhouse we concluded that production of 2n microspores in this species is under genetic control. However, the variation in the frequency of 2n microspores production among producer plants and also among anthers of the same flower suggested differences in gene expression. On the other hand, although the flowering period of T. sidoides is extended throughout spring and summer (Solís Neffa, 2000), the frequency of 2n microspores yield by each producer plant cultivated in the greenhouse increased up to 22% in summer. Therefore, it was suggested that the higher production of unreduced microspores might be caused by the environmental stress due to the high temperatures to which plants of T. sidoides were exposed (Kovalsky and Solís Neffa, 2012).

Figure 3 Sporads and pollen in Turnera sidoides. A-D) Sporads: A) Monad. B) Dyad. C) Tryad. D) Tetrad. E-F) Pollen grains. E) Unreduced pollen grain (2n). F) Reduced (n) pollen grain. Bar= 5 μm.
Finally, in order to contribute to the understanding of the cytological mechanisms involved in the production of unreduced male gametes, we analysed the meiotic behaviour and pollen viability of 2n and 4n gamete producers from diploid populations of T. sidoides. The results obtained showed that meiotic behaviour was mostly regular; however, some irregularities such as parallel spindles, fused spindles and nuclear restitution were also detected. Pollen viability varied from 44.75% to 95.82%. Overall, our results suggested that nuclear restitution at both the first and the second meiotic division were involved in the production of 2n male gametes in T. sidoides (Kovalsky et al., 2014).
Evidence of the production of 2n eggs in diploid plants of T. sidoides complex
As a second step to understand how unreduced gametes may have contributed to the origin and establishment of polyploids in natural diploid populations of T. sidoides, we provide the first evidence of the production of 2n megaspores by progeny test and by flow cytometric analysis of seeds (Figure 4) collected in natural populations (Kovalsky and Solís Neffa, 2016). In spite of the low number of 2n megaspores detected, our results suggested that, in natural diploid populations of T. sidoides, some plants would be more likely to produce 2n megaspores than others. This data agrees with our previous findings in this species, which demonstrated that only some S and L plants (26%) were capable of producing 2n and 4n microspores and, that the capability of producing such unreduced gametes is under genetic control (Kovalsky and Solís Neffa, 2012, 2015). The finding of only some plants of T. sidoides that produced 2n megaspores suggests that their production could be under genetic control as well. Furthermore, although the frequency of plants producing 2n megaspores in T. sidoides was lower than that previously reported for plants producing 2n microspores (Kovalsky and Solís Neffa, 2012), all plants that produced 2n megaspores also produced 2n microspores. However, the fact that plants producing 2n microspores do not always produce 2n megaspores it was proposed that their simultaneous production may be independent from each other. Our results also suggested differences in the relative contributions of 2n microspores and 2n megaspores to polyploid formation. In diploid-tetraploid crosses, 2n megaspores are supposed to be more likely to generate viable seeds than 2n microspores (Thompson and Lumaret, 1992; Ramsey and Schemske, 1998) and it was suggested that formation of neopolyploids in natural populations would proceed in a similar way to these interploidy crosses (Ramsey, 2007). However, this would not be the case of T. sidoides, since the major frequency of triploid embryos from 2n microspores in seeds collected in natural populations suggested that 2n microspores would contribute more than 2n megaspores to the origin of neopolyploids. Although, this would not imply that triploid embryos generated by fusion of 2n microspores and n megaspores result in plants which will effectively establish in populations of T. sidoides. In this species, plants which produced 2n megaspores develop more viable seeds per fruit, and seeds have a higher germination rate than those plants that exclusively produce 2n microspores (Kovalsky, 2012). This suggested that, although in T. sidoides 2n microspores were involved in the origin of most triploid embryos found in natural populations, the ploidy ratios among embryo and endosperm and/or epigenetic processes might confer triploids originated from the fusion of 2n megaspores with n microspores an advantage during more advanced stages of their development and establishment, being effectively established in diploid populations (Kovalsky and Solís Neffa, 2016).

Figure 4 Flow cytometry histograms of triploid seeds of Turnera sidoides. A) Triploid seed originated from an 2n microspore showing and embryo with a peak at 3C and a peak at 4C corresponding to the endosperm. B) Flow cytometry histogram of a triploid seed originated from an 2n megaspore showing and embryo with a peak at 3C and a peak at 5C corresponding to the endosperm.
Unilateral vs bilateral sexual polyploidization
Both unilateral and bilateral sexual polyploidization may be involved in the origin of the polyploids of T. sidoides in natural populations. Because all polyploids of the complex so far analysed have an even ploidy level (Fernández, 1987; Solís Neffa and Fernández, 2001; Solís Neffa et al., 2004), bilateral polyploidization was expected to be the most important mechanism of polyploid origin in the complex (Panseri et al., 2008). However, taking into account that this species is dystilous and outbreeder (Solís Neffa, 2000), for bilateral sexual polyploidization to occur both, S and L individuals should produce 2n microspores and 2n megaspores. In this sense, our finding of 2n microspores (Kovalsky and Solís Neffa, 2012), together with the detection of 2n megaspores (Kovalsky and Solís Neffa, 2016) in non-hybrid diploid populations of T. sidoides, and the fact that both the L and S plants can produce both 2n microspores and 2n megaspores, suggested that bilateral sexual polyploidization can occur in natural populations of this species. However, owing to the limited chances of fertilization between simultaneously formed 2n microspores and 2n megaspores, bilateral polyploidization would occur less frequently than the unilateral sexual polyploidization. The finding of triploids in natural diploid populations of T. sidoides (Elías et al., 2011; Kovalsky and Solís Neffa, 2012; Kovalsky et al., 2018) may reflect triploid formation through the union of n and 2n gametes, suggesting that unilateral polyploidization by a triploid bridge may be an alternative mechanism of polyploid formation in this species complex.
Multiple origin of polyploids
The formation of autopolyploids was considered a rare event, but it was later recognised that multiple origins typify polyploid plant species (Segraves et al., 1999). In T. sidoides, the multiple diploid-polyploid transitions revealed in a phylogenetic analysis based on the sequences of the plastid DNA regions supported a multiple origin of autopolyploids in each subspecies and morphotypes (Solís Neffa et al., 2022), as previously suggested for the complex (Solís Neffa and Fernández, 2001). Also, the finding of lineage-exclusive haplotypes in diploid and polyploid populations were interpreted as independent polyploidization events. The occurrence of polyploids with unique haplotypes, not directly related to any of the haplotypes detected within the diploids analysed, suggested an additional independent origin of polyploids (Solís Neffa et al., 2022). Likewise, analyses of the genetic variability and structure of the diploid and polyploid populations of T. sidoides using nuclear molecular markers, showed that the greater genetic similarity of tetraploids to diploids from the same geographic region than to diploids from other regions was consistent with the origin of polyploids in multiple polyploidization events from genetically differentiated diploid populations (Panseri, 2012; Dabrio et al., 2020). Such multiple polyploidization events detected in T. sidoides complex were related to the capacity of many diploid populations to produce unreduced gametes (Panseri et al., 2008; Kovalsky and Solís Neffa, 2012, 2016), but also to the ability of polyploids to survive and establish in nature.
POLYPLOID ESTABLISHMENT IN T. SIDOIDES
Owing to the lower rate of neopolyploid formation in T. sidoides complex, their establishment constitute a critical step. Thus, for a better understanding of the evolution of polyploidy in T. sidoides our next objective was to assess the possible factors influencing neotetraploids establishment in natural populations.
Patterns of cytotype variation of T. sidoides in a diploid- tetraploid contact zone
As a first step, we examined the cytotypes structure in a diploid-tetraploid contact zone of T. sidoides in the mountain ranges of central Argentina, aiming to analyse whether the frequency and distribution of cytotypes at microgeographical scale was explained by ecological sorting in heterogeneous environments or, alternatively, due to competitive cytotype exclusion (Elías, 2010; Elías et al., 2011). The results evidenced that diploids and tetraploids were spatially segregated, although both cytotypes can occur in close proximity. Diploids grew in the piedmont of the hills, tetraploids in the adjacent valley, while patches of diploids and triploids plants were found, in the transition zone of both cytotypes. Since diploids and tetraploids occur under similar climatic conditions, it is unlikely that climate influences cytotype distribution in the contact zone. Also, the edaphic conditions would only partially contribute to the spatial segregation of cytotypes at local scale. The similar ecological preferences of both cytotypes and the lack of mixed diploid-tetraploid patches suggested that the separate distribution of cytotypes in the contact zone may be independent of the selective environment; diploids and tetraploids being unable to coexist as a result of reproductive exclusion (Elías, 2010; Elías et al., 2011).
Additionally, to assess whether differences in the biological fitness between cytotypes would be influencing the cytotype distribution and frequency in the contact zone, we analysed the variation in reproductive and phenological traits of diploids and tetraploids (Elías, 2010; Panseri, 2012). From this study it was evident that although the average number of seeds per fruit was similar in both cytotypes, the continuity of flowering and fruiting in tetraploids in relation to the discontinuity of diploids, the highest number of bloomed individuals per site as well as the greater number of fruits per individual in tetraploids result in an increase number of propagules that tetraploids contribute to the seed bank as well as to the next generation, increasing the frequency of tetraploids in each generation. This fact, together with the perennial condition of T. sidoides and its ability to multiply asexually by rhizomes (Solís Neffa, 2000) would favour the success and establishment of neotetraploids in natural populations. Also, the occurrence of occasional cases of self-compatibility and successful illegitimate crosses (S × S and L × L) in polyploids of T. sidoides suggests that such polyploids may produce seeds by selfing or from crosses between plants of the same floral type, further increasing their chances of establishment and expansion (Solís Neffa, 2000; Solís et al., 2020).
The role of 2n gametes in polypoid establishment of the T. sidoides
Theoretical models of polyploid evolution suggest that 2n gamete production by diploids is an essential factor in the dynamics of mixed diploid-tetraploid populations since tetraploids are more likely to establish or to be maintained at a low frequency within diploid populations when they are formed recurrently through the union of 2n gametes (Levin, 1975; Felber and Bever, 1997). Turnera sidoides is an outbreeder, therefore, the occurrence of plants that continuously form many 2n microspores and 2n megaspores would play a key role in the establishment of neopolyploids in natural diploid populations. Taking into account that T. sidoides grow in discrete populations (mostly with fewer than 100 individuals, Solís Neffa, 2000), the occurrence of a low number of plants producing 2n gametes may be significant to polyploid dynamics in diploid populations. Moreover, since the capability to produce 2n microspores is a heritable trait in this species complex and, the frequency of production of 2n microspores was higher in the progeny of 2n microspores producers (Kovalsky and Solís Neffa, 2015), the frequency of 2n microspores and 2n megaspores and, consequently, the likelihood of origin of neopolyploids by sexual polyploidization would increase after successive generations. Additionally, considering that T. sidoides is a perennial species and, that its seeds are mostly dispersed by gravity in such a way that individuals concentrate in localized areas (Solís Neffa, 2000), the progeny of plants that produce 2n gametes would concentrate near the mother plant, increasing the likelihood of crosses between 2n microspores and 2n megaspores producers. The continuous formation of neopolyploids as a consequence of successive backcrosses between 2n gametes producers and their progeny would favour the establishment and persistence of such neopolyploids in diploid populations of T. sidoides. Furthermore, although diploid individuals produced low levels of 2n gametes but, under certain environmental conditions, such production may be increased, enhancing the likelihood that a low frequency of polyploids can be originated and maintained in this population (Kovalsky and Solís Neffa, 2015).
The role of triploids in the origin and evolution of polyploids of T. sidoides complex
Simulation models have demonstrated that the evolution of tetraploids in a diploid population depends on the reproductive efficiency of triploids and the ploidy level of functional gametes (n=x, n=2x and, n=3x) they produce, contributing to the formation of neopolyploids in each generation (Husband, 2004). Thus, aiming to account for the role of triploids in the origin and demographic establishment of tetraploids of T. sidoides in natural populations, we analysed the microsporogenesis of triploids as well as the crossabilty among diploid, triploid and tetraploid plants of the complex (Kovalsky et al., 2018). Triploids exhibit irregular meiotic behaviour that results in unbalanced and/or non-viable gametes and, low pollen viability. Although, in spite of abnormalities in chromosome pairing and unequal chromosome segregation, triploids are not completely sterile and form viable gametes (pollen viability up to 67%). Triploids originated by the fusion of 2n × n gametes showed more regular meiotic behaviour and higher fertility than triploids from the contact zone among diploids and tetraploids (Elías, 2010) and, triploids with hybrid origin (Moreno et al., 2015). Also, our results evidenced that reproductive isolation among cytotypes of T. sidoides was not so strong. In this species complex most populations are constituted by a single cytotype, even though some mixed diploid - triploid - tetraploid populations were also detected (Elías et al., 2011; Kovalsky and Solís Neffa, 2012; Moreno et al., 2015). This fact together with our finding of triploids in the progeny of 2x × 4x experimental crosses, evidence that triploid block may be frequently overcome, being not enough to prevent the regular occurrence of triploids in natural populations of T. sidoides (Kovalsky et al., 2018). Moreover, although in this species the success of crosses involving triploids is generally low, their gametes can participate in fertilization, producing progeny of variable ploidy level. The diploid plants used for experimental crossings produced reduced gametes (n=x), therefore, the frequency of diploid and triploid progeny obtained from crosses 2x × 3x constituted an indirect measure of the frequency of n and 2n gamete production by triploids. The results of our studies evidenced that triploids of T. sidoides produce n and 2n gametes, although n gametes were the most frequently produced. This finding was supported by the fact that the progeny of 3x × 4x crosses was 100% triploid, resulted of the fusion of n gametes of triploid and tetraploid parents. Besides, the experimental crosses performed evidenced that triploids produce diploid and, triploid progeny, suggesting that new generations of triploids and tetraploids would originate by crossings between triploids or by backcrosses with diploid progenitors that produce 2n microspores and 2n megaspores. Therefore, triploids of T. sidoides would contribute both to the origin of new tetraploids (Kovalsky et al., 2018) and to gene flow among diploids and tetraploids in the contact zones (Moreno et al., 2015).
POLYPLOIDY AND RANGE EXPANSION
Since the analysis of geographical differentiation of cytotypes provides useful insights into the evolutionary dynamics and ecological differentiation of polyploids, we employed a cytogeographical approach for T. sidoides. The results so far have revealed that, at a geographical scale and considering the complex as a whole, diploids have restricted and disjunct distributions, while tetraploids are the most widespread, being the cytotype that gives continuity to the complex throughout the distribution area. Likewise, populations with higher ploidy levels are rare and were found in disjunct, and usually marginal, areas of the complex. These patterns of cytotypes frequencies and distribution support the classification of T. sidoides as a mature polyploid complex as was initially proposed (Solís Neffa, 2000) following the criteria of Stebbins (1971).
Proximal and historical nonexclusive hypotheses were proposed to explain the spatial patterns of the cytotypes in this species (Solís Neffa, 2000; Solís Neffa and Fernández, 2001; Solís Neffa et al., 2004). Given that the widespread latitudinal and longitudinal range of T. sidoides encompasses a wide diversity of climates and ecological regions with contrasting precipitation regimes, it was first hypothesised that the geographical separation of the cytotypes was due to differences in habitat requirements. However, taking into account the relative frequency and geographical distribution of the cytotypes, it was also hypothesised that the distribution of extant cytotypes might reflect the Quaternary geomorphologic and climatic changes that affected most of the current geographical range of T. sidoides. Consequently, restricted diploids may be relictual populations that have survived these periods of change, while such historical events could provide favourable conditions for the establishment of polyploids, with the wider distribution of tetraploids being the result of range expansion (Solís Neffa and Fernández, 2001; Solís Neffa et al., 2004; Elías et al., 2011).
Additionally, the integration of cytogeographic and genetic divergence data together with past niche modelling provided further insights into the geographic context of the processes driving divergence and range expansion in T. sidoides complex (Solís Neffa et al., 2022). The projected model for the mid-Pliocene (~3.3-3.0 Myr), revealed four areas of highest climatic suitability for a possible ancestor of T. sidoides. The main one, in the NW of the current species range (following the direction of main orographic systems) were coincident with the proposed refugial centres for T. sidoides complex based on plastid genome diversity (Speranza et al., 2007, Moreno et al., 2018). This area was also proposed as the ancestral geographic area for T. sidoides (Moreno et al., 2018). Additionally, three smaller suitable areas were located in the Pampean ranges and in NE Argentina as well as in the SE of the current range, along the Serranías del Este (Uruguay) (Solís Neffa et al., 2022). The spreading of large areas of semi-arid conditions in the Chaquean Domain concomitantly with the final uplift of the Andes (Gregory-Wodzicki, 2000) would have acted as a barrier to gene flow among populations of T. sidoides, promoting the divergence between the two main lineages by the time of the Great Patagonian Glaciations (~1.17 Myr). Further diversification of lineages within the two main clades of T. sidoides complex mostly coincides with colder periods, suggesting that cooling and drying during Pleistocene (Villagrán and Hinojosa, 1997; Rabassa et al., 2011) may have caused the isolation and divergence of ancestral T. sidoides complex populations (Moreno et al., 2018).
The predicted distribution of diploids during interglacial periods suggests a more continuous area of diploids than at present, probably along the most elevated terrains (Moreno et al., 2018; Solís Neffa et al., 2022). The evolutionary history of the complex from the Last Glacial Maximum up to present day mostly involved the range expansion of the allopatrically differentiated populations to their current limits. During the Mid-Holocene, the warmer and wetter climate would have promoted the westward reduction of the area of diploids and the range expansion of tetraploids. Semiarid conditions during the Late Holocene (Iriondo and García, 1993) may have determined the progressive retraction of the suitable area for diploids, while the subhumid climate period would have favoured the persistence of diploids in the inter-Andean valleys (Solís Neffa et al., 2022).
EVOLUTIONARY SIGNIFICANCE OF AUTOPOLYPLOIDY IN T. SIDOIDES
Cytogenetic data together with past niche modelling as well as cytogeographic and genetic divergence analyses provided strong evidence that T. sidoides complex is in an active process of intraspecific allopatric diversification at diploid level since the early Pleistocene. Further diversification of the complex involved the emergence of independent polyploid series in each morphologically divergent lineage.
Polyploidy did not produce extensive morphological changes in T. sidoides, so diploids and polyploids within each subspecies and morphotypes can only be distinguished from each other on the basis of detailed analysis of microcharacters (Panseri, 2012) and quantitative traits (Solís Neffa, 2000). These results together with cytogenetic (Solís Neffa, 2000; Roggero Luque et al., 2015) and molecular data (Moreno et al., 2015) suggested that autopolyploidy did not contributed significantly to the diversification of T. sidoides. Instead, changes associated with genome size variation and the gigas effect were recorded in the polyploid series of the complex (Solís Neffa, 2000). All these chromosome doubling-induced changes may have had an adaptive significance that may promote an increase range of subspecies and morphotypes through the expansion of polyploids arising from multiple diversified diploid populations of T. sidoides (Solís Neffa et al., 2022). In this sense, despite the fact that polyploid series of each subspecies and morphotype of T. sidoides inhabits the same ecoregion, the cytotypes tend to be spatially segregated at a more local scale (Solís Neffa and Fernández, 2001; Solís Neffa et al., 2004; Elías et al., 2011; Solís Neffa et al., 2022). A clear example was demonstrated in the serrano morphotype of subspecies pinnatifida (Elías et al., 2011), in which diploids and polyploids, although inhabiting the same ecoregion, are segregated along altitudinal and bioclimatic gradients at regional/local scales. The wider distribution of tetraploids compared to their related diploids in each polyploid series of T. sidoides complex supports the hypothesis that autopolyploids may have been more effective colonisers of new ecological niches.

