










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMultequina
versión On-line ISSN 1852-7329
Multequina vol.18 no.2 Mendoza jul./dic. 2009
ARTÍCULO ORIGINAL
Asteráceas de importancia económica y ambiental. Primera parte. Sinopsis morfológica y taxonómica, importancia ecológica y plantas de interés industrial
Asteraceae of economic and environmental importance. First part. Morphological and taxonomic synopsis, environmental importance and plants of industrial value
Luis A. Del Vitto1 y E. M. Petenatti1
1 Herbario y Jardín Botánico UNSL, Cátedras Farmacobotánica y Famacognosia, Facultad de Química, Bioquímica y Farmacia, Universidad Nacional de San Luis, Ej. de los Andes 950, D5700HHW San Luis, Argentina. lvitto@unsl.edu.ar.
RESUMEN
Las Asteráceas incluyen gran cantidad de especies útiles (medicinales, agrícolas, industriales, etc.). Algunas han sido domesticadas y cultivadas desde la Antigüedad y otras conforman vastas extensiones de vegetación natural, determinando la fisonomía de numerosos paisajes. Su uso etnobotánico ha ayudado a sustentar numerosos pueblos. Hoy, unos 40 géneros de Asteráceas son relevantes en alimentación humana y animal, fuentes de aceites fijos, aceites esenciales, forraje, miel y polen, edulcorantes, especias, colorantes, insecticidas, caucho, madera, leña o celulosa. Otras son importantes malezas y/o plantas tóxicas para el hombre y el ganado, algunas causan alergia y otras resultan ornamentales. En terapéutica son usados gran número de metabolitos secundarios sintetizados por Asteráceas. Se ofrece una síntesis de las Asteráceas útiles y dañinas, taxonómicamente actualizadas, considerando usos etnobotánicos y propiedades fitoquímicas y farmacológicas, con énfasis en las entidades silvestres, cultivadas o adventicias que viven en Argentina. En esta primera parte se discuten los caracteres generales de la familia, hábitat y dispersión, fitoquímica, y los géneros y especies de valor industrial y de mayor importancia ecológica y ambiental, así como el estado de conservación de sus miembros, considerados a nivel global, todo ello a la luz de la actual sistematización de la Familia.
Palabras clave: Botánica económica; Etnobotánica; Plantas útiles; Plantas nocivas
SUMMARY
Asteraceae include many useful species (medical, agricultural, industrial, etc.). Some have been domesticated and cultivated since ancient times and other up vast tracts of natural vegetation, determining the physiognomy of many landscapes. Their ethnobotanical use has sustained a lot of peoples all over the world. Today, some 40 genera are relevant for human and animal feeding, sources of eating oils, essential oils, fodder, honey and pollen, sweeteners, spices, dyes, insecticides, rubber, wood, firewood or cellulose. Others are major weeds and/or plants toxic to humans and livestock, some cause allergies and others are valuable ornamentals. A great number of therapeutic secondary metabolites are synthesized by Asteraceae, while others acts as insectifuge. It provides a synthesis of the Asteraceae useful and harmful Asteraceae, taxonomically updated, considering ethnobotanical uses and phytochemical and pharmacological properties, with emphasis on wild, cultivated or weed entities lived in Argentina. In this first part discusses the general characteristics of the family, habitat and dispersal, phytochemistry, genera and species of industrial, ecological and environmental importance, and conservation status of its members, taken globally, all this in the light of the current systematization of the Family.
Key Words: Economic botany; Ethnobotany; Noxious plants; Useful plants
INTRODUCCIÓN
La Familia de plantas Asteraceae Bercht. & J. Presl (1820) corresponde al Orden Asterales, Suborden Asteridae, y está caracterizada por sus inflorescencias racimosas en capítulos, con flores individuales epíginas rodeadas de 1-varias hileras de brácteas involucrales, sobre el receptáculo común en que remata el escapo o rama florífera (Weberling, 1989). Por dicho carácter es considerada un grupo natural y uniforme, altamente evolucionado, y recibe el nombre alternativo Compositae Giseke (1792).
Comparada con otras grandes familias de plantas (como Poáceas y Fabáceas) tiene menor valor económico para el Hombre, aunque incluye una cantidad de especies útiles desde diversos puntos de vista. El uso etnobotánico de muchas de ellas ha ayudado al progreso y sustento de gran número de pueblos en todo el mundo (Parodi, 1934; Guillaumin, 1948; Dawson, 1960), safisfaciendo sus necesidades de alimento, forraje, leña, medicinas, etc.
Como otras plantas, algunas Asteráceas han sido sometidas desde la Antigüedad al complejo proceso de domesticación (Parodi, 1938; Dempewolf et al., 2008,) luego del establecimiento de las prácticas agrícolas básicas (Parodi, 1935, 1966; Harris, 1967, 1969; Darlington, 1969). Hoy son cultivos de importancia (Elzebroeck & Wind, 2008) cuyos ancestros y parientes silvestres pueden ser empleados en el mejoramiento genético (Vavilov, 1926/1951; Hawkes, 1969; Darlington, 1973).
Desde el punto de vista estrictamente económico, unas 40 especies tienen importancia directa en alimentación humana (hortalizas y "semillas" oleaginosas) e indirectamente por productos obtenidos por la industria. Otras especies silvestres tienen potencial nutricional (Wilkes, 1977), muchas son de interés tecnológico u ornamental, y centenares rinden metabolitos secundarios de uso farmacéutico o industrial (Wagner, 1977; Evans, 2002) o aportan néctar y polen para la producción apícola (Howes, 1953; Pellett, 1976), forraje para la producción ganadera (Boelcke, 1964), etc. Muchas Asteráceas son pioneras u oportunistas, y gran número resultan malezas de cultivos y/o tóxicas para el ganado (Marzocca et al., 1979; Ragonese & Milano, 1984). Además, numerosas especies tienen un rol destacado en la estructura y funcionamiento de los ecosistemas (Heywood et al., 1977; Hind et al., 1996).
El objeto de este trabajo es presentar una sinopsis de los grupos utilitarios y nocivos de Asteráceas, refiriendo la sistemática del grupo al más moderno sistema de clasificación y estableciendo la importancia de esta Familia en las diversas actividades humanas.
Importancia de las Asteráceas a nivel global y en la Argentina
Es la familia de Fanerógamas con mayor diversidad a nivel mundial. Representa 8-10% de la flora global y abarca 22.750 taxones específicos e infraespecíficos, con 1.620 géneros (Stevens, 2007) incluyendo algunos de los más numerosos como Senecio (c. 1.250 especies), Hieracium (c. 1.000) y Helichrysum (c. 600; Mabberley, 1998).
Es de distribución subcosmopolita, pues está representada en todos los continentes excepto la Antártida y el hinterland de Groenlandia (Stevens, 2007). Ocupa hábitats diversos, desde el nivel del mar hasta el límite altitudinal de la vegetación. La mayor diversidad se halla en zonas tropicales, subtropicales y templadas; abundan en biomas de montaña y en regiones áridas y semiáridas. La distribución continental está correlacionada con algunos representantes de ciertas tribus (Bohm & Stuessy, 2001).
En Argentina es la más numerosa de las familias de plantas superiores con 1.500 especies y unos 200 géneros, es decir 16% del total de plantas vasculares del país, con 26% de endemismos (Cabrera, 1961; Zuloaga & Morrone, 1999, Zuloaga et al., 1999; Katinas et al., 2007). En sectores andinos y patagónicos constituyen hasta 20% de la flora (Cabrera, 1957, 1971, 1974). Interesan tanto por su amplia distribución y abundancia como por la hipótesis que considera la Patagonia como probable lugar de origen de la familia (Stuessy et al., 1996). Contribuciones de autores locales han permitido conocer la diversidad de la familia en el país (Cabrera, 1961, 1965, 1971, 1974, 1978; Ariza, 1994-2005; Hunziker, 1994; Zuloaga & Morrone, 1999, etc.) y en Chile (Moreira-Muñoz & Muñoz-Schick, 2007).
Caracteres diacríticos de la Familia
La constancia de caracteres morfoanatómicos, químicos, moleculares y adaptativos tiene gran valor sistemático (Carlquist, 1961; Hummel & Stäsche, 1962; Hegnauer, 1964; Metcalfe & Chalk, 1983; Bohm & Stuessy, 2001; Panero & Funk, 2002; Funk et al., 2005), aún en híbridos (Rollins, 1944). Destacan la presencia/ausencia de canales secretores, tejido laticífero, engrosamiento secundario, principios químicos "marcadores", tipología de capítulos, verticilos de protección y sexuales, granos de polen, indumento, etc., secuencias de nucleótidos, biótipos y tipos funcionales y adaptaciones morfofisiológicas.
Predominan las hierbas anuales (terófitos), bienales (protohemicriptófitos) o perenes (hemicriptófitos, geófitos), y en alta montaña abundan sufrútices enanos (caméfitos pulvinados) y subarbustos erguidos o rastreros (caméfitos sufrutescentes y caméfitos reptantes, respectivamente); en ambientes xéricos predominan subarbustos (caméfitos) y arbustos (nanofanerófitos), muchos de ellos rizomatosos o radicantes; algunos géneros tropicales y subtropicales están representados por arbolitos (microfanerófitos), incluyendo "árboles en roseta", "candelabriformes" o "palmeriformes", y más raramente árboles elevados (megafanerófitos hasta 30 m alto y 1 m diámetro), e incluso trepadoras o apoyantes herbáceas (fanerófitos herbáceos) o leñosas (lianas).
En algunos géneros (Achyrocline, Gamochaeta, p.p. Helichrysum, Hypochaeris, Podolepis, Senecio, Vittadinia) existe simbiosis micorrícica, tanto vesículo-arbuscular (VAM) como ectomicorrícica (Newman & Reddell, 1987; Smith & Read, 1997), con hongos de los géneros Glomus (VAM) y Laccaria (ectomicorrizante).
Los tallos son erguidos, postrados o escandentes, raro áfilos; cuando herbáceos con un anillo de haces vasculares concéntricos, cada uno protegido por un casquete de fibras. Las hojas son alternas (raro opuestas), con frecuencia en roseta basal, generalmente pecioladas, a veces sésiles o atenuadas en pseudopecíolo, decurrentes sobre los tallos; las láminas son simples o 1-, 2- ó más veces pinnati- o palmatilobas, -fidas o -sectas, raro compuestas; los estomas son generalmente anomocíticos. Los tricomas son glandulosos y/o eglandulosos y las estructuras secretoras son externas e internas; su rica tipología resulta útil en taxonomía y sobre todo en el control de calidad de hierbas y otras plantas útiles (Uphof & Hummel, 1962); en algunos grupos los canales laticíferos se distribuyen por toda la planta (Fahn, 1979; Metcalfe & Chalk, 1983; Esau, 1987).
Las flores son epíginas, perfectas, diclinas (pistiladas o estaminadas) o neutras; los sépalos están ausentes o modificados en cerdas, escamas, aristas o pelos simples o ramificados, constituyendo un papus o vilano (Cabrera, 1978) que ayuda a la diseminación de los frutos y tiene funciones defensivas (Stuessy & Garver, 1996); los pétalos son connatos, formando corolas (3) 5-meras, actinomorfas (tubulosas, pentadentadas) o zigomorfas (liguladas, bilabiadas, filiformes); el androceo es sinantéreo, con (4) 5 estambres alternando con los lóbulos de la corola, los filamentos se insertan en el tubo corolino y las anteras son introrsas, connatas, formando un tubo alrededor del estilo; el polen es variable y la exina presenta notable esculturación (Erdtman, 1952-1971, 1969; Skvarla et al., 1977) que fija las micrósporas al cuerpo de los polinizadores (Markgraf et al., 1981; Lane, 1996), aunque en pocos casos están adaptados a la polinización anemófila (Bolick, 1990). El ovario es ínfero, 2-carpelar, 1-locular y 1-ovulado, con óvulo anátropo de placentación basal; el estilo es filiforme, apicalmente dividido en 2 ramas estigmáticas papilosas o provistas de pelos colectores subapicales; en la antesis el crecimiento progresivo del estilo a través del "tubo anteral" empuja el polen hacia arriba (por ello la flor perfecta actúa inicialmente como masculina, luego se despliegan las ramas estigmáticas resultando funcionalmente femenina, y finalmente esas ramas se repliegan hacia abajo, tocando a veces el polen remanente y permitiendo una autofecundación tardía; Ladd, 1994); los frutos son secos, en cipsela (Roth, 1977), generalmente fusiformes u obcónicos, a veces rostrados, a menudo provistos de papus (los adaptados a la dispersión anemócora), a veces sin él o provistos de cerdas espinosas (los adaptados a la dispersión zoócora; Fahn & Werker, 1972); las semillas se presentan en número de 1 por fruto, son exalbuminadas y a veces su testa es oscura a causa de la "fitomelanina" (Pandey & Dhakal, 2001), con embrión recto y germinación epígea (Corner, 1976; Johri, 1984).
Las inflorescencias elementales son del tipo indefinido (racimosas), pauci- a multifloras (raramente unifloras), en capítulos formados por un receptáculo (común a todas las flores) plano a convexo, a veces cónico y hasta columnar, paleáceo o no, a veces cerdoso, otras foveolado, rodeado por un involucro de 1-5 (15 a más) series de brácteas o filarias, a veces con calículo; con (1) 5-300 ó más flores individuales isomorfas o heteromorfas en un mismo capítulo, que puede ser radiado (flores periféricas liguladas, pistiladas o neutras y flores del disco tubulosas, perfectas o estaminadas), capitado (con todas las flores liguladas, perfectas) o eradiados, en cuyo caso pueden ser discoides (todas las flores son actinomorfas, tubulosas, perfectas), disciformes (con 1-varias series de flores filiformes, pistiladas y flores centrales tubulosas, perfectas, a veces estaminadas) o radiantes (con todas las flores actinomorfas, las periféricas con corolas más grandes, lobadas y expandidas, perfectas, pistiladas o neutras, y las centrales con corolas menores, perfectas). A menudo los capítulos están agrupados a su vez en inflorescencias mixtas, cimiformes, corimbiformes, racemiformes o espiciformes (Harris, 1995).
Aunque muchas Asteráceas sólo se reproducen por semillas, buena parte presentan también multiplicación vegetativa. La diversificación sexual y el mecanismo de presentación del polen inducen tipos de reproducción muy variados, con flores monoclinas, diclinas o neutras que determinan capítulos homógamos o heterógamos, dioecia estricta o facultativa, ginomonoecia, ginodioecia, hasta poligamia, etc. y permiten expresar una cantidad casi ilimitada de combinaciones adaptables a muy diversas condiciones ambientales. Por ello pueden ser tanto autógamas como alógamas, presentándose con frecuencia autoincompatibilidad del tipo esporofítico homomórfico (Hiscock & Tabah, 2003; Allen& Hiscock, 2008).
La multiplicación vegetativa es importante en especies, líneas o clones productivos que presentan baja tasa de producción de semillas viables, o en híbridos naturales o artificiales de baja fertilidad; tiene lugar mediante rizomas, raíces gemíferas o tuberosidades radicales acompañadas de la porción basal de tallo; por vía artificial se suma la división de matas en plantas vivaces, las estacas en leñosas o semileñosas, los esquejes, etc. Numerosas especies presentan apomixis obligada (Davis, 1967).
El número cromosómico básico sería x = 8, que es el de la Subfamilia más primitiva, Barnadesioideae (Stuessy et al., 1996); los números hallados son x = 2 - (5-20) - 120, y hasta más de 400 en Olearia albida Hook. f. de Nueva Zelanda (Smith-White, 1968; Watanabe & Smith-White, 1987); los más frecuentes son x = 8, 9 y 12 (Solbrig, 1977; Watanabe, 2008); las semejanzas o variaciones numéricas intra- e interespecíficas favorecen o impiden, respectivamente, la hibridación (Sundberg & Stuessy, 1990). Las condiciones citogenéticas influyen en la especiación, presentándose alo- o autopoliploidía, y variantes metabólicas que llevan a la síntesis de metabolitos secundarios bioactivos, a menudo útiles como "marcadores" quimiotaxonómicos (Bohm & Stuessy, 2001).
Muestra la mayor plasticidad adaptativa entre las familias de Magnoliopsida y se considera la más evolucionada de ese grupo; la escasez de registros fósiles indica una relativa juventud filética, presentando a la vez caracteres "primitivos" y "avanzados"; por ello es difícil establecer interrelaciones basadas sólo en caracteres morfológicos; recientes estudios moleculares han permitido formular un sistema filogenético tentativo que mejora la concepción taxonómica de la familia (Panero & Funk, 2002; Funk et al., 2005).
Hábitat y Adaptaciones
La amplia distribución y gran plasticidad adaptativa han generado diversidad de bioformas y tipos funcionales. Las variaciones morfoanatómicas son resultado de adaptaciones correlacionadas con hábitos normales de crecimiento y con la especialización ecológica inducida mayormente por el ambiente, hallándose ejemplos emblemáticos (Metcalfe & Chalk, 1983; Bell, 1993). Los géneros Eupatorium, Vernonia, Senecio y Sonchus muestran un casi completo espectro de formas biológicas, desde hierbas anuales (incluso efímeras) hasta fanerófitos, con grupos hidrofíticos, helofíticos, mesofíticos y xerofíticos (Raunkiaer, 1934; Stebbins, 1977).
Las estrategias adaptativas de Asteráceas a condiciones ambientales extremas son diversas (Solbrig, 1961, 1993; Böcher & Lyshede, 1972; Laetsch, 1974; Seddon, 1974, Smith & Turner, 1975, Ruthsatz, 1978, Ancibor, 1980; van der Pihl, 1982; Orshan, 1986; Vogel, 1990 a,b; Fahn & Cutler, 1992; Petenatti & Del Vitto, 2000): adaptaciones al frío y la sequía (formas vegetativas, época de floración, indumento, posición de estomas, formas de crecimiento, modalidades de germinación, caducidad foliar, microfilia y hasta afilia); sindrome de polinización predominantemente entomófilo; diseminación prevalentemente anemócora (raro zoócora); espinescencia, acaulia o paquicaulia; crecimiento en cojín y adaptación a intensa irradiación solar, a bajas temperaturas y a vientos intensos; suculencia; producción de látex, aceites esenciales o resinas, como respuesta defensiva ante predatores o adaptativa a condiciones extremas; ciclo biosintético "C4" en algunos géneros o especies, correlacionado con anatomía "Kranz"; heliofilia o esciofilia; etc.
La composición química y su importancia en el aprovechamiento y la taxonomía
El conocimiento bioquímico del grupo ha aportado datos relevantes a la taxonomía y ha explicado o facilitado el empleo de las mismas en las actividades económicas. Ya Hegnauer (1964) estableció los caracteres fitoquímicos generales, sustentados y ampliados por Harborne (1977), Mabry & Bohlmann (1977), Seaman (1982), Bohm & Stuessy (2001) y Aniszewski (2007). En general, el grupo está caracterizado por la presencia de ácidos iso- y clorogénico, isoflavonoides, lactosas sesquiterpénicas, alcoholes triterpénicos pentacíclicos, aceites esenciales (con predominio de terpenoides), alcaloides (especialmente pirrolizidínicos) y diversos derivados acetilénicos, mientras que carece de taninos verdaderos y de iridoides (Bohlmann, 1977). La acumulación de ciertos metabolitos primarios y secundarios determina la utilidad o aplicación de las plantas. La síntesis de glúcidos en último término conduce a la formación de fructanos del tipo inulina (Hegnauer, 1964) acumulados en órganos subterráneos (Bruneton, 1995) y semillas; son la principal reserva hidrocarbonada, de gran digestibilidad y reemplazan el almidón. La síntesis de ciclitoles también caracteriza a las Asteráceas, especialmente L-inositol y su isómero esciloinositol (Plouvier, 1963; Hegnauer, 1977).
Las semillas de Asteráceas almacenan glúcidos (particularmente como inulina), prótidos (como proteína amorfa) y lípidos. Los aceites seminales son ricos en ácidos grasos de cadenas cortas, sobre todo linoleico y esteárico y en menor cantidad oleico, linolénico y palmítico (Smith, 1985); algunas son fuente industrial de aceites comestibles.
Los metabolitos secundarios aislados de Asteráceas son muy variados. Algunos flavonoides y aceites volátiles (con di- y triterpenos) son comunes a casi todas las especies (Harborne, 1977; Mabry & Bohlmann, 1977; Seaman et al., 1990; Bohm & Stuessy, 2001) y 2 grupos de sustancias son "marcadores quimiotaxonómicos" de la Familia: las lactonas sesquiterpénicas y los compuestos poliacetilénicos (Bohlmann et al., 1973; Herz, 1977; Mabry & Bohlmann, 1977; Sörensen, 1977; Reaman, 1982). En general estas plantas carecen de alcaloides, salvo los de núcleo pirrolizidínico (Aniszewski, 2007). Finalmente, es llamativa la ausencia de iridoides, aminoácidos no proteicos y taninos verdaderos.
Sistematización de las Asteráceas
Inicialmente fue dividida en 2 subfamilias: Asteroideae Lindl. Y Cichorioideae Kitam. Según el sistema de Bentham, que data de 1873, corresponden 12 tribus a la primera y una sola a la segunda. Pero desde los trabajos de Bremer (1987) y Jansen & Palmer (1987), evidencias moleculares han servido de base para una nueva sistematización de la familia, determinando la existencia de varios clados supragenéricos. Bremer & Jansen (1992) establecieron la mayor antigüedad filética del taxón basal de la familia, un grupo finalmente designado Barnadesioideae con rango de Subfamilia; su notoria presencia en la flora argentina (en los Andes y el sur del país) ha sustentado la hipótesis del origen patagónico de la familia Asteráceas (Stuessy et al., 1996). Cuando Bremer (1994, 1996) reinterpretó los caracteres macromorfológicos de los grupos, resultó un sistema de 3 subfamilias: Barnadesioideae, Asteroideae y Cichorioideae, con 17 tribus. Estudios filogenéticos moleculares (sobre todo la secuenciación del ADN cloroplastidial por Baldwin et al., 2002; Eldenäs et al., 1999; Jansen et al., 1991, 1992; Kim et al., 1992; Kim & Jansen, 1995) llevaron a escindir la Subfamilia Cichorioideaey revalidar antiguos conceptos genéricos o crear nuevos géneros. Todo ello condujo a la proposición de clasificaciones cada vez más detalladas (APG II, 2003), como la sugerida por Panero & Funk (2002), dividiendo la Familia en 10 Subfamilias y 35 Tribus. Thorne & Reveal (2007) validaron el nombre Carduoideae en lugar de Cichorioideae, que ha mostrado ser un taxón parafilético, y expandieron el número de tribus a 25, con 3 nuevas tribus identificadas por análisis molecular (Baldwin et al., 2002), aunque mantuvieron Heliantheae, que se considera un grupo polifilético. Ampliando la base de datos moleculares, Panero & Funk (2002) y Funk et al. (2005) produjeron un nuevo esquema tentativo, aceptado parcialmente por Jeffrey (2007), con 24 tribus dispuestas en 5 subfamilias. Recientemente se elevó el número de subfamilias y tribus, confirmando el carácter monofilético de las subfamilias Barnadesioideae y Asteroideae, mientras que los clados ubicados entre las Cichorioideae de Bremer (1994) y las Carduoideae de Thorne & Reveal (2007) pasaron a integrar alguna de estas tres subfamilias: Mutisioideae, Carduoideae y Cichorioideae; además, han sido establecidas otras subfamilias monotípicas, representadas por géneros como Corymbium, Gymnarrhena, y Hecastocleis, cada una de linajes monofiléticos. Así, en las recientes clasificaciones sugeridas por Panero & Funk (2007, 2008) se reconocen 12 subfamilias estrictamente monofiléticas, que comprenden 40 tribus, según muestra la Tabla 1.
Tabla 1. Sistematización de la Familia Asteraceae según Panero & Funk (2008) y otros
Table 1. Systematization of the Family Asteraceae according to Panero & Funk (2008) and others

Asteráceas útiles y perjudiciales
A los fines prácticos, las Asteráceas utilitarias o perniciosas para las actividades humanas han sido agrupadas aquí en 13 conjuntos, mencionando los géneros (y en algunos casos las especies) de mayor importancia, en base a la experiencia transmitida desde las más tempranas o clásicas menciones bibliográficas (D. Parodi, 1886; Correa, 1926; Bois, 1927; Parodi, 1934, 1939; Cabrera, 1939; Guillaumin, 1948) hasta las más actualizadas, ya sea de carácter general (Bailey, 1949, 1954; Hill, 1952; Uphof, 1968; Usher, 1974; Allaby, 1992; Wickens, 2004) como regional (Pittier, 1908, 1971; Pérez Arbeláez, 1947/1978; Baranov, 1967; Cárdenas, 1969; Rizzini & Mors, 1976; Huang & Ling, 1996; Boer & Ella, 2000; Cabrera et al., 2000; Van Wyk, 2000), e incluso los alimentos o forrajes de emergencia que han servido para paliar las épocas de hambruna (De Castro, 1952; Purdue University, 2008), y las que regionalmente son reconocidas por nuestra experiencia personal, e incluso algunas evaluaciones económico-ambientales (Godoy et al., 1993). Algunos taxones son empleados con múltiples propósitos y son mencionados en diversos grupos.
1. Plantas de Valor Industrial
Entre las Asteráceas de las que se obtienen productos industriales, destacan las oleaginosas (que además tienen grandes perspectivas para la elaboración de combustibles como el biodiesel), unas pocas plantas cauchíferas, otras muchas con metabolitos intensamente amargos y una con una importante sustancia edulcorante.
1.1. Oleaginosas: La reserva de los frutos ("semillas") de muchas especies de Asteráceas es fuente de aceite de considerable importancia en alimentación humana y animal (Vaughan, 1970; Weiss, 1999; Van der Vossen & Mkamilo, 2007). Entre ellas destaca el "girasol" (Helianthus annuus L.), planta anual originaria de Norteamérica, cultivada en casi todo el mundo templado y que ha sufrido un fuerte proceso de selección (Seiler & Rieseberg, 1997; Burke et al., 2005); benefician sus frutos ("pipas"), que se consumen crudos o tostados, salados o no; son ricos en aceite comestible, que se emplea puro o en mezcla; mientras las semillas de formas salvajes y ramificadas contienen 22-36% de aceite en peso, las de los híbridos comerciales cultivados hoy llegan a 40-50% (Bruneton, 1995; Burke et al., 2005; Paniego et al., 2006); la "torta" remanente de la extracción, rica en prótidos, integra parte fundamental de dietas animales.
Los frutos del "cártamo", "alazar" o "falso azafrán" (Carthamus tinctorius L.) rinden aceite comestible ("aceite de cártamo") con alto contenido en ácido linoleico, que también ha sido usado en medicina. De los frutos de "madí" (Madia sativa Mol.), nativa del sur de Argentina y Chile, también se beneficia el aceite; su cultivo y aprovechamiento como oleaginosa por los indígenas data de época precolombina (Parodi, 1940; Dawson, 1960). Por su parte, en Oriente se desarrolló el cultivo de "ramtil" o "Niger seed" (Guizotia abyssinica (L. f.) Cass.), una hierba anual, norafricana, cuyo aceite seminal es comestible e industrial, los frutos son usados en mezclas para alimentar pájaros y el follaje se emplea como verdura.
Los aceites de alta calidad de "girasol" y "cártamo" tienen bajo tenor de ácidos grasos saturados y alta concentración de insaturados; algunos cultivares mutagénicos de ambas especies rinden otros tipos de aceites. Muchos ácidos grasos interesantes y nuevos para la industria se hallan en los aceites de otras Asteráceas, tales como los dienólicos conjugados de Dimorphoteca, los acetilénicos (predominantemente vernólico y crepenínico) de Crepis, los epoxi obtenidos de Vernonia y de Stokesia (Wagner, 1977; IENICA, 2007) y los que están formados mayormente por ácidos grasos trans, obtenidos de algunas especies de Asterothamnus, Artemisia, Heteropappus, Saussurea y Youngia (Tsevegsüren et al., 2000).
Los llamados "aceites" de "caléndula" (Calendula officinalis L.) y de "diente de león" (Taraxacum officinale Web. in Wigg.) tienen aplicación en cosmética y en realidad son maceraciones de inflorescencias de esas plantas en aceite de oliva virgen y otros (Worwood, 1991). Por otra parte, las oleaginosas en general han abierto un amplio campo de aplicación industrial en la elaboración de combustibles ("biodiesel"), sobre todo las especies de grandes rendimientos agrícolas.
En la Tabla 2 se da una lista de los géneros más conocidos de Asteráceas con "semillas" oleíferas.
Tabla 2. Géneros con semillas oleíferas
Table 2. Genera witho oil-bearing seeds

1.2. Edulcorantes: Stevia rebaudiana (Bertoni) Bertoni es una planta originaria de Paraguay y actualmente cultivada en cierta escala para la obtención de un glicósido diterpénico, el esteviósido, una sustancia 300 veces más dulce que la sacarosa y sólo 20% menos dulce que la sacarina. El rendimiento es de 65 g de esteviósido/1.000 g de materia seca (Wagner, 1977) y su empleo se ha ido incrementando, sobre todo por la toxicidad de algunos edulcorantes artificiales, integrándoselo a alimentos funcionales (Madrigal & Sangronis, 2007).
1.3. Con Principios Amargos: Las sustancias amargas son productos del metabolismo secundario muy frecuentes en las Asteráceas. Pese a ello sólo pocas especies, como Artemisia absinthium L. y Centaurea benedicta L., han alcanzado importancia por este motivo en farmacia e industria alimentaria. El "ajenjo", debe su sabor a lactonas sesquiterpénicas como el guaianólido dimérico absintina, por lo cual es empleado para elaborar bases de licores aperitivos como "vermouth"; en cuanto a Centaurea benedicta, una lactona de tipo germacreno, esterificada, llamada cnicina, le confiere un sabor amargo menos intenso; ambas son usadas en la industria farmacéutica como estimulantes del apetito (aperitivos) y digestivos (Wagner, 1977; Evans, 2002). En Sudamérica austral la industria licorera se basa en unas pocas especies para elaborar los aperitivos denominados "amargos" (tanto alcohólicos como analcohólicos), especialmente a partir de las Asteráceas Pluchea sagittalis (Lam.) Cabrera, "lucera", y las "marcelas" (Achyrocline spp., Gnaphalium spp.) y "vira-viras" (Gnaphalium spp., Senecio spp.), que a su vez tienen usos en medicina popular por su acción eupéptica.
1.4. Sacaríferas: Otras Asteráceas se emplean para la obtención industrial de polisacáridos. Como ha quedado dicho, estas plantas se caracterizan por acumular inulina, un polisacárido compuesto por 20-30 unidades de fructosa, mucho más digestible que otros carbohidratos, y mejor tolerado incluso por los diabéticos, para quienes se prepara un pan especial con estos productos. Además, la inulina se usa para la elaboración técnica de fructosa. En menor escala, raíces y rizomas de Inula y de Helianthus tuberosus L., tuberosidades radicales de Dahlia spp., raíces de Taraxacum officinale Web. ex Wigg. y de Arctium lappa L., además de las de Cichorium intybus L., son usadas con cierta frecuencia como fuentes alternativas de carbohidratos.
1.5. Cauchíferas: Algunas Asteráceas rinden una cantidad apreciable de látex, que es empleado limitadamente en la industria del caucho (Báez et al., 1948; Sívori & Caso, 1952; Marzocca, 1964.a; Lewinsohn, 1991; Boer & Ella, 2000). Para obtener caucho industrialmente se emplean sobre todo dos especies: "kok-saghyz" (Taraxacum bicorne Dahlst.), una planta nativa del centro de Rusia con 8-10% de látex en la raíz, y que fuera intensamente estudiada durante la segunda guerra mundial como fuente alternativa ante la escasez del caucho producido por Hevea brasiliensis (Willd. ex A. Juss.) Muell. Arg. (Whaley, 1947), incluso con posibilidades ciertas de una mecanización de la extracción (Marzocca & Fanti, 1952); y "guayule" (Parthenium argentatum A. Gray), un arbusto de las regiones áridas de México, que se hibrida naturalmente con otras especies silvestres (Rollins, 1944; Ray, 1993; National Academy of Sciences, 2002). Los cauchos obtenidos de ambas especies se usan para mezclar con caucho sintético, al que confieren mayor elasticidad, necesaria en ciertas aplicaciones industriales de este producto. También son cauchíferas, aunque con menor productividad, "krym-saghyz" (Taraxacum megalorhizon Hand-Mazz.), y "tau-saghyz" (Scorzonera tau-saghyz Lipschitz & Bossé). En EE.UU. se ha estudiado también la obtención de caucho a partir de especies de Solidago (los "goldenrods"), pero con muy bajo rendimiento. Por último, debe indicarse que Chrysothamnus nauseosus (Pallas) Britt. es otra potencial fuente de caucho, llamado "chrysil" (Hall & Goodspeed, 1919).
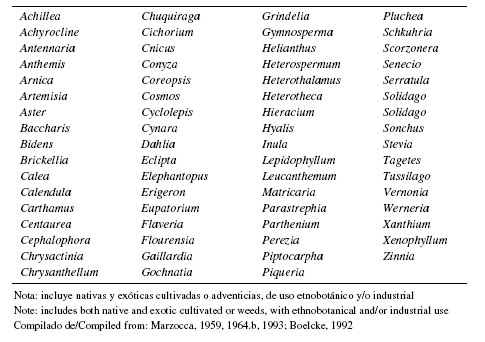
1.6. Tintóreas y Taníferas: Las plantas constituyen las principales fuentes de sustancias colorantes y tintóreas, y entre ellas las Asteráceas juegan un importante papel (Cannon & Cannon, 2003; Cardon, 2007), acrecentado porque la industria pone cada día más interés en los pigmentos de origen natural, tanto sea por razones ambientales como sanitarias y bromatológicas. Por su parte, la industria del curtido siempre ha requerido de las sustancias tánicas y los taninos derivados de algunas Asteráceas leñosas comienzan a ser relevantes (Torres Romero, 1983). Además, es importante el uso etnobotánico que siempre se ha hecho de las Asteráceas como plantas tintóreas y taníferas (Marzocca, 1959, 1993; Torres Romero 1983).
Algunas Asteráceas contienen tintes empleados industrialmente o en arte culinario, como el ya mencionado "cártamo" (Carthamus tinctorius L.), el "aciano" (Centaurea cyanus L., también ornamental) y la "vara de oro" (Solidago virgaurea L.). El alto contenido en flavonoides de numerosas Asteráceas nativas y exóticas las ha convertido en plantas útiles desde este punto de vista, desde la antigüedad (Hill, 1952; Bailey, 1954).
En la Tabla 3 se da una lista de los géneros con representantes tintóreos, colorantes y taníferos mencionados para la Argentina.
Tabla 3. Géneros con especies tintóreas y taníferas, silvestres y cultivadas en Argentina
Table 3. Genera with dyeing and tanner species, wild and cultivated in Argentina
Importancia ecológica y status de conservación
Numerosas Asteráceas tienen un papel destacado en la estructura y funcionamiento de los ecosistemas; en efecto, vastas extensiones de vegetación natural y seminatural están dominadas por Asteráceas, o en su caso acompañan las especies principales en las asociaciones dominantes, configurando con frecuencia la fisonomía de los ambientes naturales, lo que les confiere un valor ecológico y ambiental inapreciable. En muchos casos alcanzan importantes valores de cobertura, y cumplen funciones fundamentales en la retención y estabilización del suelo, en el incremento de su contenido de materia orgánica, en la producción de oxígeno, la depuración de la atmósfera, etc.
Actualmente un gran número de Asteráceas son ruderales o se comportan como malezas, y por ello abundan en áreas disturbadas o en cultivos agrícolas en todo el mundo, pero a su vez existe un significativo número de especies (particularmente en regiones montañosas, tanto tropicales como subtropicales y hasta templadas) que son endemismos restringidos, y que a causa de dramáticas modificaciones en el hábitat (particularmente debidas a la acción antrópica) se ven amenazadas o en peligro de extinción. Así, en la Lista Roja de plantas compilada por la Unión Internacional de Conservación de la Naturaleza (IUCN, 2009) se hallan representados 147 géneros de Asteráceas, que abarcan 475 especies con diversas calificaciones de amenaza de extinción. Pese a que se puede considerar aún muy incompleto el conocimiento de la flora y el estado de conservación de las plantas de vastas regiones del globo, las cifras mencionadas son llamativas, puesto que prácticamente el 2% de las especies y poco más del 9% de los géneros de la Familia Asteráceas conocidas en todo el mundo se encuentran incluidas en estos registros. De ellas, 4 especies se consideran completamente extintas (EX; las abreviaturas son las establecidas como normativa por la IUCN); 2 ya se han extinguido en la naturaleza (EW), aunque se conservan ejemplares en cultivo; 65 se encuentran críticamente amenazadas de extinción (CR); 91 están amenazadas (EN); 174 se hallan en condición de vulnerabilidad (VU); 42 han sido calificadas como casi amenazadas (NT); 49 en estado que provoca preocupación (LC); y las 22 restantes presentan bajo riesgo, repartiéndose en este caso entre bajo riesgo/prácticamente amenazadas (16) y bajo riesgo/preocupación (6); finalmente, de 26 de ellas se presume que están amenazadas de una u otra forma (DD), pero por el momento se carece de datos fidedignos para establecer indubitablemente su status (Mace & Lande, 1991; Mace et al., 1992; IUCN, 2009).
En la Tabla 4 se da una lista de los géneros de Asteráceas que incluyen taxones infragenéricos amenazados de extinción, y que constan en las Listas Rojas de la IUCN.
Tabla 4. Géneros de Asteráceas con número de taxones amenazados de extinción de cada uno, registrados a nivel mundial
Table 4. Genera of Asteraceae with number of threatened taxa of each, registered worldwide

BIBLIOGRAFÍA
1. ALLABY, M. (ed.), 1992. The Concise Oxford Dictionary of Botany. 2º ed. Oxford, Univ. Press. vi, 442 pp. [ Links ]
2. ALLEN, A.M. & S.J. HISCOCK, 2008. Evolution and Phylogeny of Selfincompatibility Systems in Angiosperms. En: Franklin-Tong (ed.), Self Incompatibility in Flowering Plants. Berlin/Heidelberg, Springer. Pp. 73-101. [ Links ]
3. ANCIBOR, E., 1980. Estudio anatómico de la puna de Jujuy. II. Bol. Soc. Argent. Bot. 19: 157-202. [ Links ]
4. ANISZEWSKI, T., 2007. Alkaloids, Secrets of Life. Amsterdam, Elsevier. 334 pp. [ Links ]
5. APG II (Angiosperm Phylogeny Group). 2003. An update of the APG classification for the orders and families of flowering plants. Bot. J. Linn. Soc. 141: 399-436. [ Links ]
6. ARIZA-ESPINAR, L., 1994-2005. Pródromo de la flora fanerogámica de Argentina Central, 1: 1-65 (1994); 2: 1-111 (2000): , 3: 1-139 (2005). Córdoba, Museo Botánico. [ Links ]
7. BÁEZ, C.R., H.G. FISCHER & E.M. SÍVORI, 1948. Plantas caucheras. Revta. Fac. Agron. La Plata 26 (2): 182-200. [ Links ]
8. BAILEY, L.H., 1949. Manual of cultivated plants. New York, Macmillan. 1116 pp. il. [ Links ]
9. BAILEY, L.H., 1954. The Standard Cyclopedia of Horticulture. 17º impr. New York, Macmillan. 3 vols. [ Links ]
10. BALDWIN, B.G., B.L. WESSA & J.L. PANERO, 2002. Nuclear rDNA evidence for major lineages of helenioid Heliantheae (Compositae). Syst. Bot. 27: 161-98. [ Links ]
11. BARANOV, A.I., 1967. Wild vegetables in Manchuria. Econ. Bot. 21:140-55. [ Links ]
12. BELL, A.D., 1993. Plant form. Oxford, Univ. Press. 341 pp. [ Links ]
13. BÖCHER, T.W. & O.B. LYSHEDE, 1972. Anatomical studies in xerophytic apophyllous plants, II. Biol. Skr., Danske Vid. Selsk. 18 (4): 1-137. [ Links ]
14. BOELCKE, O., 1964. Forrajeras. En: Parodi (dir.), Encicl. Argent. Agric. Jard. 2, 2da. parte: 707-87. Buenos Aires, Acme. [ Links ]
15. BOER, E. & A.B. ELLA (eds.), 2000. Plant Resources of South East Asia 18. Plants producing exudates. Leiden, Backhuys. 189 pp. [ Links ]
16. BOHLMANN, F., 1977. Summary of the chemistry of the Compositae. En: Heywood, Harborne & Turner (eds.), The Biology and Chemistry of the Compositae, 2: 1097-1104. London, Academic Press. [ Links ]
17. BOHLMANN, F., T. BURKHARDT & C. ZDERO (eds.), 1973. Naturally occuring Acetylenes. London/New York, Academic Press. 547 pp. [ Links ]
18. BOHM, B.A. & T.F. STUESSY, 2001. Flavonoids of the Sunflower Family (Asteraceae). Vienna, Springer. 831 pp. [ Links ]
19. BOIS, D., 1927. Les plantes alimentaires chez tous les peuples et a travers les ages. Histoire, utilisation, culture. Phanérogames Legumières. Paris, Lechevalier. 593 pp. [ Links ]
20. BOLICK, M.R., 1990. The pollen surface in wind-pollination with emphasis on the Compositae. En: Hesse & Ehrendorfer (eds.), Pl. Syst. & Evol. Suppl. 5: 39-51. Wien/New York, Springer. [ Links ]
21. BREMER, K. 1987. Tribal interrelationships of the Asteraceae. Cladistics 3: 210-53. [ Links ]
22. BREMER, K., 1994. Asteraceae: Cladistics and Classification. Portland, Timber Press. 752 pp. [ Links ]
23. BREMER, K. 1996. Major clades and grades of the Asteraceae. En: Hind & Beentje (eds.), Compositae: Systematics, 1: 1-7. Kew, Royal Bot. Gardens. [ Links ]
24. BREMER, K. & R.K. JANSEN, 1992. A new subfamily of the Asteraceae. Ann. Missouri Bot. Gard. 79: 414-15. [ Links ]
25. BRUNETON, J., 1995. Pharmacognosy, phytochemistry, medicinal plants. 2º ed. Paris. Lavoisier/Intercept. 700 pp. [ Links ]
26. BURKE, J.M., S.J. KNAPP & L.H. RIESEBERG, 2005. Genetic consequences of selection during the evolution of cultivated sunflower. Genetics 171: 1933-40. [ Links ]
27. CABRERA, A.L., 1939. Las Compuestas útiles cultivadas en la República Argentina. Bol. Direcc. de Agric., Ganadería e Ind. (Buenos Aires) 1939: 5-31. [ Links ]
28. CABRERA, A.L., 1957. La vegetación de la Puna argentina. Revista Invest. Agríc. (Buenos Aires) 11 (4): 317-412. [ Links ]
29. CABRERA, A.L., 1961. Compuestas Argentinas: Clave para la determinación de los géneros. Revista Mus. Argent. Ci. Nat. Bernardino Rivadavia, Bot. 2: 291-362. [ Links ]
30. CABRERA, A.L., 1965. Compositae. En: Cabrera (dir.), Flora Prov. Buenos Aires. Colecc. Ci. INTA 4 (6): 1-443. Buenos Aires. [ Links ]
31. CABRERA, A.L., 1971. Compositae. En: Correa (dir.), Flora Patagónica. Colecc. Ci. INTA 8 (7): 1-451. Buenos Aires. [ Links ]
32. CABRERA, A.L., 1974. Compositae. En: Burkart (ed.), Flora Ilustrada de Entre Ríos. Colecc. Ci. INTA 6 (6a): 106-554. Buenos Aires. [ Links ]
33. CABRERA, A.L., 1978. Compositae. En: Cabrera (dir.), Flora de la provincia de Jujuy. Colecc. Ci. INTA 12 (10): 1-726. Buenos Aires. [ Links ]
34. CABRERA, A.L., J.V. CRISCI, G. DELUCCHI, S.E. FREIRE, D.A. GIULIANO, L. IHARLEGUI, L. KATINAS, A.A. SÁENZ, G. SANCHO & E. URTUBEY, 2000. Catálogo ilustrado de las Compuestas (= Asteraceae) de la Provincia de Buenos Aires, Argentina. COBIOBO nº 2/PROBIOTA nº 1. La Plata. 136 pp. [ Links ]
35. CANNON, J. & M. CANNON, 2003. Dye Plants and Dyeing, 2º ed. Portland, Timber Press. 128 pp. [ Links ]
36. CÁRDENAS, M., 1969. Manual de plantas económicas de Bolivia. Cochabamba, Icthus. 421 pp.. [ Links ]
37. CARDON, D., 2007. Natural dyes. Sources, tradition, technology and science. London, Archetype Books. 500 pp. [ Links ]
38. CARLQUIST, S., 1961. Comparative plant anatomy. New York, Holt/ Rinehart/Winston. ix, 146 pp. [ Links ]
39. CORNER, E.J.H. 1976. The seeds of Dicotyledons. Cambridge, Univ. Press. 2 vols. [ Links ]
40. CORREA, M.P., 1926. Diccionario das plantas uteis do Brasil e das exoticas cultivadas. Río de Janeiro, Imp. Nacional. 2 vols. [ Links ]
41. DARLINGTON, C.D., 1969. The silent millennia in the origin of agriculture. En: Ucko & Dimbleby, The domestication and exploitation of plants and animals. London, Duckworth. Pp. 67-72. [ Links ]
42. DARLINGTON, C.D., 1973. Chromosome botany and the origins of cultivated plants. 3º ed. rev., New York, Hafner. [ Links ]
43. DAVIS, G.L., 1967. Apomixis in the Compositae. Phytomorphology 77: 270-77. [ Links ]
44. DAWSON, G., 1960. Los alimentos vegetales que América dio al mundo. Mus. La Plata, Ser. Técn. Didáct. 8: 1-68. [ Links ]
45. DE CASTRO, J., 1952. The geography of hunger. London, Gollancz. 288 pp. [ Links ]
46. DEMPEWOLF, H., L.H. RIESEBERG & Q.C. CRONK, 2008. Crop domestication in the Compositae: a family-wide trait assessment. Genet. Resour. Crop Evol. 55: 1141-57. [ Links ]
47. ELDENÄS, P. , M. KÄLLERSJÖ & A.A. ANDERBERG , 1999. Phylogenetic placement and circumscription of tribes Inuleae s. str. and Plucheeae (Asteraceae). Molec. Phylog. & Evol. 13: 50-8. [ Links ]
48. ELZEBROEK, T. & K. WIND (eds.), 2008. Guide to cultivated plants. Wallingford, CABI Publ. 540 pp. [ Links ]
49. ERDTMAN, G., 1969. Handbook of Palynology. Copenhagen, Munskgaard. 846 pp. [ Links ]
50. ERDTMAN, G., 1952-1971. Pollen Morphology and Plant Taxonomy; an introduction to palynology. Stockholm/Waltham, Almqvist & Wiksell/ Chronica Bot. 4 vols. [ Links ]
51. ESAU, K., 1987. Anatomía de las plantas con semilla. Buenos Aires, Hemisferio Sur. 512 pp. [ Links ]
52. EVANS, W.C., 2002. Trease & Evans´Pharmacognosy. 15º ed. Edimburg, Saunders.585 pp. [ Links ]
53. FAHN, A., 1979. Secretory Tissues in Plants. London, Academic Press. 302 pp. [ Links ]
54. FAHN, A. & D.F. CUTLER, 1992. Xerophytes. Berlin/Stuttgart, Gebr. Bornträger.176 pp. [ Links ]
55. FAHN, A. & E. WERKER, 1972. Anatomical mechanisms of dispersal. En: Kozlowski (ed.), Seed Biology, 1, Chap. IV: 151-221. New York, Academic Press. [ Links ]
56. FUNK, V.A ., R.J. BAYER, S. KEELEY, R. CHAN, L. WATSON, B. GEMEINHOLZER, E.E. SCHILLING, J.L. PANERO, B.G. BALDWIN, N.T. GARCÍA JACAS, A. SUSANNA, & R.K. JANSEN. 2005. Everywhere but Antarctica: using a supertree to understand the diversity and distribution of the Compositae. Biol. Skr. 55: 343-73. [ Links ]
57. GODOY, R., R. LUBOWSKY & A. MARKANDYA, 1993. A method for the economic valuation of non-timber tropical forest products. Econ. Bot. 47: 220-33. [ Links ]
58. GUILLAUMIN, A., 1948. Les plantes sauvages. Biologie et utilisation. Paris, Payot. 219 pp. [ Links ]
59. HALL, M.M. & T.H. GOODSPEED, 1919. Chrysil, a new rubber from Chrysothamnus nauseosus. Univ. Calif. Publ. Bot. 7: 159-78, 183-264, 265-78. [ Links ]
60. HARBORNE, J.B., 1977. Flavonoid profiles in the Compositae. En: Heywood, Harborne & Turner (eds.), The Biology and Chemistry of the Compositae, 1: 359-84. London, Academic Press. [ Links ]
61. HARRIS, D.R., 1967. New light on plant domestication and the origins of agriculture: a review. Geogr. Rev. 57: 90-107. [ Links ]
62. HARRIS, D.R., 1969. Agricultural systems, ecosystems and the origins of agriculture. En: Ucko & Dimbleby, The domestication and exploitation of plants and animals. London, Duckworth. Pp. 3-15. [ Links ]
63. HARRIS, E.M., 1995. Inflorescence and floral ontogeny in Asteraceae: a synthesis of historical and current concepts. Bot. Rev. 61: 93-278. [ Links ]
64. HAWKES, J.G., 1969. The ecological background of plant domestication. En: Ucko & Dimbleby, The domestication and exploitation of plants and animals. London, Duckworth. Pp. 17-29. [ Links ]
65. HEGNAUER, R., 1964. Chemotaxonomie der Pflanzen, 3, Dicotyledoneae, Part I (Chemical Series, 18). Stuttgart, Birkhauser. 743 pp. [ Links ]
66. HEGNAUER, R., 1977. The chemistry of the Compositae. En: Heywood, Harborne & Turner (eds.), The Biology and Chemistry of the Compositae, 1: 283-335. London, Academic Press. [ Links ]
67. HERZ, W., 1977. Sesquiterpene lactones in the Compositae. En: Heywood, Harborne & Turner (eds.), The Biology and Chemistry of the Compositae, 1: 337-357. London, Academic Press. [ Links ]
68. HEYWOOD, V.H., J.B. HARBORNE & B.L. TURNER, 1977. An overture to the Compositae. En: Heywood, Harborne & Turner (eds.), The Biology and Chemistry of the Compositae, 1: 1-20. London. Academia Press. [ Links ]
69. HILL, A.E., 1952. Economic Botany. 2º ed. Tokyo, McGraw-Hill/Kogakusha. 560 pp. [ Links ]
70. HIND, D.J.N., H.J. BEENTJE & P.D.S. CALIGARI (eds.), 1996. Compositae: Biology and Utilization. Kew, Royal Bot. Gardens. 2 vols. 1673 pp. [ Links ]
71. HISCOCK, S.J. & D.A. TABAH, 2003. The different mechanisms of sporophytic self-incompatibility. Phil. Trans. R. Soc. London, Ser. B, 358: 1037-45. [ Links ]
72. HOWES, F.N., 1953. Plantas melíferas. Flora silvestre y cultivada, de valor para la vida del colmenar y la cosecha de miel. Barcelona, Reverté. 326 pp. [ Links ]
73. HUANG, Y.-P. & Y.-R. LING, 1996. Economic Compositae in China. En: Caligari & Hind (eds.), Compositae: Biology and utilization. 2. Kew, Royal Bot. Gardens. Pp. 431-51. [ Links ]
74. HUMMEL, K. & K. STÄSCHE, 1962. Die Verbreitung der Haartypen in den natürlichen Verwandschaftsgruppen. En: Linsbauer, Handbuch der Pflanzenanatomie, 4 (5): 272-291. Berlin, Gebr. Bornträger. [ Links ]
75. HUNZIKER, A.T. (dir.), 1994-. Flora Fanerogámica Argentina (Asteraceae: L. Ariza-Espinar, coord. y AA.VV., fascículos 13-14, 28, 29, 31, 45-48, 59-63, 66, 81-83). Buenos Aires/Córdoba, Proflora/CONICET. [ Links ]
76. IENICA (Interactive European Network for Industrial Crops and their Applications), 2007. Summary report for European Union, Crops for specialty chemicals. En: http://www.ienica.net, consultado 30.III.2009. [ Links ]
77. IUCN, 2009. Lista de géneros de Asteráceas amenazados de extinción. En: http://www.iucnredlist.org/search/ asteraceae. Consultado 6.II.2009. [ Links ]
78. JANSEN, R.K. & J.D. PALMER, 1987. A chloroplast DNA inversion marks an ancient evolutionary split in the sunflower family, Asteraceae. Proc. Natl. Acad. Sci. USA 84: 5818-22. [ Links ]
79. JANSEN, R.K., H.J. MICHAELS & J.D. PALMER, 1991. Phylogeny and character evolution in the Asteraceae based on chloroplast DNA restriction site mapping. Syst. Bot. 16: 98-115. [ Links ]
80. JANSEN, R.K., H.J. MICHAELS, R.S. WALLACE, K.J. KIM, S.C. KEELEY, L.E. WATSON & J.D. PALMER, 1992. Chloroplast DNA variation in the Asteraceae: Phylogenetic and Evolutionary Implications. En: Soltis, Soltis & Doyle (eds.), Molecular Systematics of Plants, New York, Chapman & Hall. Pp. 252-79. [ Links ]
81. JEFFREY, C., 2007. Compositae: Introduction with key to tribes. En: Kadereit & Jeffrey (eds.), Families and Genera of Vascular Plants. VIII, Flowering Plants, Eudicots, Asterales. Pp. 61-87. Berlin, Springer. [ Links ]
82. JOHRI, B.M. (ed.), 1984. Embryology of Angiosperms. Berlin, Springer. 819 pp. [ Links ]
83. KATINAS, L., D.G. GUTIÉRREZ, M.A. GROSSI & J.V. CRISCI, 2007. Panorama de la Familia Asteraceae (= Compositae) en la República Argentina. Bol. Soc. Argent. Bot. 42 (1-2): 113-29. [ Links ]
84. KIM, K.-J. & R.K. JANSEN, 1995. ndhF sequence evolution and the major clades in the sunflower family. Proc. Natl. Acad. Sci. USA 92: 10379-83. [ Links ]
85. KIM, K.-J., R.K. JANSEN, R.S. WALLACE, H.J. MICHAELS & J.D. PALMER, 1992. Phylogenetic implications of rbcL sequence variation in the Asteraceae. Ann. Missouri Bot. Gard. 79: 428-45. [ Links ]
86. LADD, P.G. 1994. Pollen presenters in flowering plants - form and function. Bot. J. Linnean Soc. 115: 165-195. [ Links ]
87. LAETSCH, W.M., 1974. The C4 syndrome: a structural analysis. Ann. Rev. Plant Physiol. 25: 27-52. [ Links ]
88. LANE, M., 1996. Pollination biology of Compositae. En: Caligari & Hind (eds.), Compositae: Biology and Utilization. Kew, Royal Bot. Gardens. 2: 61-80 [ Links ]
89. LEWINSOHN, T.M. 1991. The geographical distribution of plant latex. Chemoecology 2: 64-68. [ Links ]
90. MABBERLEY, D.J., 1998. The Plant Book. 2º ed. Cambridge, Univ. Press. 858 pp. [ Links ]
91. MABRY, T.J. & F. BOHLMANN, 1977. Summary of the chemistry of the Compositae. En: Heywood, Harborne & Turner (eds.), The Biology and Chemistry of the Compositae. 2: 1097-1104. London, Academia Press. [ Links ]
92. MACE, G.M. & R. LANDE. 1991. Assessing extinction threats: toward a re-evaluation of IUCN threatened species categories. Conservation Biology 5: 148-157. [ Links ]
93. MACE, G.M., N. COLLAR, J. COOKE, K.J. GASTON, J.R. GINSBERG, N. LEADER-WILLIAMS, M. MAUNDER & E.J. MILNERGULLAND, 1992. The development of new criteria for listing species on the IUCN Red List. Species 19: 16-22. [ Links ]
94. MADRIGAL, L. & E. SANGRONIS, 2007. La inulina y derivados como ingredientes claves en alimentos funcionales. Arch. Latinoam. Nutric. 57 (4): 2007. [ Links ]
95. MARKGRAF, V., H.L. D'ANTONI & T.A. AGER, 1981. Modern Pollen Dispersal of Argentina. Palynology 5: 43-63. [ Links ]
96. MARZOCCA, A., 1959. Historia de plantas tintóreas y curtientes. Buenos Aires, INTA/Min. Agric. Ganad. Nación. 234 pp. [ Links ]
97. MARZOCCA, A., 1964a. Plantas cauchíferas. En: Parodi (dir.), Encicl. Argent. Agric. Jard., 2, 2da. parte. Buenos Aires, Acme. Pp. 1201-5. [ Links ]
98. MARZOCCA, A., 1964b. Plantas tintóreas. En: Parodi (dir.), Encicl. Argent. Agric. Jard., 2, 2da. parte. Buenos Aires, Acme. Pp. 1183-94. [ Links ]
99. MARZOCCA, A., 1993. Index de plantas colorantes, tintóreas y curtientes. Manual de las especies de Argentina. Ser. nº 9, Buenos Aires, Academia Nac. Agronomía y Veterinaria. 326 pp. [ Links ]
100. MARZOCCA, A. & O.D. FANTI, 1952. Extracción mecánica de caucho de "kok-saghyz". Revista IDIA (Buenos Aires) 52: 14-6. [ Links ]
101. MARZOCCA, A., O.J. MÁRSICO & O. DEL PUERTO, 1979. Manual de malezas. Buenos Aires, Hemisferio Sur. 580 pp. [ Links ]
102. METCALFE, C.R. & L. CHALK, 1983. Anatomy of the Dicotyledons. 2º ed.,Vol. 2: 783-803. Oxford, Clarendon Press. [ Links ]
103. MOREIRA-MUÑOZ, A. & M. MUÑOZ-SCHICK, 2007. Classification, diversity, and distribution of Chilean Asteraceae: implications for biogeography and conservation. Diversity Distrib. 13: 818-828. [ Links ]
104. NATIONAL ACADEMY OF SCIENCES, 2002. Guayule: An Alternative Source of Natural Rubber. New York, Minerva. 108 pp. [ Links ]
105. NEWMAN, E.I. & P. REDDELL, 1987. The distribution of mycorrhizas among families of vascular plants. New Phytol. 106: 745-51. [ Links ]
106. ORSHAN, G., 1986. Plant form as describing vegetation and expressing adaptation to environment. Annali di Botanica 44: 7-38. [ Links ]
107. PANDEY, A.K. & M.R. DHAKAL, 2001. Phytomelanin in Compositae. Current Science 80 (8): 933-40. [ Links ]
108.PANERO, J.L. & V.A. FUNK, 2002. Toward a phylogenetic subfamilial classification for the Compositae (Asteraceae). Proc. Biol. Soc. Wash. 115: 909-922. [ Links ]
109.PANERO, J.L. & V.A. FUNK, 2007. New Infrafamilial taxa in Asteraceae. Phytologia 89: 356-360. [ Links ]
110. PANERO, J.L. & V.A. FUNK, 2008. The value of sampling anomalous taxa in phylogenetic studies: Major clades of the Asteraceae revealed. Molec. Phylogen. & Evol. 47: 757-82. [ Links ]
111. PANIEGO, N., R. HEINZ & H.E. HOPP, 2006. Sunflower. En: Genome Mapping & Molecular Breeding, Vol. II (Oilseeds): 155-179. Berlin, Springer. [ Links ]
112. PARODI, D., 1886. Notas sobre algunas plantas usuales del Paraguay, de Corrientes y de Misiones. Buenos Aires, Coni. Xxxvii, 123 pp. [ Links ]
113. PARODI, L.R., 1934. Las plantas indígenas no alimenticias cultivadas en la Argentina. Revista Argent. Agron. 1 (3): 165-212. [ Links ]
114. PARODI, L.R., 1935. Relaciones de la agricultura prehispánica con la agricultura argentina actual. Acad. Nac. Agron. Vet. Buenos Aires 1: 115-67. [ Links ]
115.PARODI, L.R., 1938. El proceso biológico de la domesticación vegetal. Rev. Arg. Agron. 5 (1): 1-24. [ Links ]
116.PARODI, L.R., 1939. Géneros de plantas cultivadas representados en la flora indígena del país. Physis 18: 255-68. [ Links ]
117. PARODI, L.R., 1940. A propósito de los vegetales de América que han conquistado el mundo. Revista Geográfica Americana 14 (82): 39-40. [ Links ]
118. PARODI, L.R., 1966. La agricultura aborigen argentina. Cuadernos de América nº 4, Buenos Aires, Eudeba. 48 pp. [ Links ]
119. PELLETT, F.C., 1976. American Honey Plants. Hamilton (U.S.A.), Dadant. 467 pp. [ Links ]
120. PÉREZ ARBELÁEZ, E., 1947/1978. Plantas útiles de Colombia. Bogotá, Lit. Arco. 210 pp. [ Links ]
121. PETENATTI, E.M. & L.A. DEL VITTO, 2000. Estructura Kranz en las especies argentinas de Flaveria (Asteraceae-Helenieae). Kurtziana 28 (2): 251-57. [ Links ]
122. PITTIER, H., 1908. Ensayo sobre las plantas usuales de Costa Rica. Washington, McQueen. 176 pp. [ Links ]
123. PITTIER, H., 1971. Manual de las plantas usuales de Venezuela y su Suplemento. Caracas, Fund. E. Mendoza. 620 pp. [ Links ]
124. PLOUVIER, V., 1963. Distribution of aliphatic polyols and cyclitols. En: Swain (ed.), Chemical Plant Taxonomy. London/New York, Academic Press. Pp. 313-36. [ Links ]
125. PURDUE UNIVERSITY, 2008. Famine foods, Asteraceae. En: http:// www.hort.purdue.edu/newcrop/faminefoods/ff_families/ ASTERACEAE.html. Consultado 18.II.2009. [ Links ]
126. RAGONESE, A.E. & V.A. MILANO, 1984. Vegetales y substancias tóxicas de la Flora Argentina. En: Kugler (dir.), Encicl. Argent. Agric. Jard., 2, fasc. 8-2: 1-413. Buenos Aires, Acme. [ Links ]
127. RAUNKIAER, C., 1934. The life forms of plants and statistical plant geography. Oxford, Clarendon Press. 632 pp. [ Links ]
128. RAY, D.T. 1993. Guayule: A source of natural rubber. En: Janick & Simon (eds.), New crops. New York, Wiley. Pp. 338-43. [ Links ]
129. RIZZINI, C.T. & W.B. MORS, 1976. Botânica econômica brasileira. São Paulo, Ed. Pedagógica e Universitária. 207 pp. [ Links ]
130. ROLLINS, R.C., 1944. Evidence for natural hybridity between guayule (Parthenium argentatum) and mariola (P. incanum). Amer. J. Bot. 31: 93-9. [ Links ]
131. ROTH, I., 1977. Fruits of Angiosperms. En: Linsbauer, Handbuch der Pflanzenanatomie, Spez Teil, Bd. 10, T. 1: 1-675. Berlin/Stuttgart, Gebr. Bornträger. [ Links ]
132. RUTHSATZ, B., 1978. Las plantas en cojín de los semidesiertos andinos del noroeste argentino. Darwiniana 21 (2-4): 491-539. [ Links ]
133. SEAMAN, F.C., 1982. Sesquiterpene lactones as taxonomic characters in the Asteraceae. Bot. Rev. 48: 121-595. [ Links ]
134. SEAMAN, F.C., H. BOHLMANN, C. ZDERO & T.J. MABRY, 1990. Diterpenes of Flowering Plants. Compositae (Asteraceae). New York, Springer. 638 pp. [ Links ]
135. SEDDON, G., 1974. Xerophytes, xeromorphs and sclerophylls: the history of some concepts in ecology. Biol. J. Linn. Soc. 6 (1): 65-87. [ Links ]
136. SEILER, G.J. & L.H. RIESEBERG, 1997. Systematics, origin and germplasm resources of the wild and domesticated sunflower. En: Schneiter (ed.), Sunflower Technology and Production. Madison, Amer. Soc. Agronomy. Pp. 21-66. [ Links ]
137. SÍVORI, E.M. & O.H. CASO, 1952. Contenido de caucho en plantas de la flora Argentina. Publ. Téc. (n.s.) nº 68: 157-165. Buenos Aires, Inst. Bot. Agrícola. [ Links ]
138. SKVARLA, J.J., B.L. TURNER, V.C. PATEL & A.S. TOMB, 1977. Pollen morphology in the Compositae and in morphological related families. En: Heywood, Harborne & Turner (eds.): The biology and chemistry of the Compositae, 1: 141-248. New York, Academic Press. [ Links ]
139. SMITH, B.N. & B.L. TURNER, 1975. Distribution of Kranz syndrome among Asteraceae. Amer. J. Bot. 62 (5): 541-5. [ Links ]
140. SMITH, C.R., 1985. Unusual seed oils and their fatty acids. En: Pryde (ed.), Fatty acids. Champaign, Amer. Oil Chemists Soc. 29-47. [ Links ]
141. SMITH, S.E. & D.J. READ, 1997. Mycorrhizal symbiosis. 2º ed. San Diego, Academic Press. 605 pp. [ Links ]
142. SMITH-WHITE, S., 1968. Brachycome lineariloba: A species for experimental cytogenetics. Chromosoma 23 (3): 359-64. [ Links ]
143. SOLBRIG, O.T., 1961. Leaf and modal anatomy of some Andean Compositae: Astereae. J. Arnold Arb. 42 (3): 276-94. [ Links ]
144. SOLBRIG, O.T., 1977. Chromosomal cytology and evolution in the family Compositae. En: Heywood, Harborne, Turner (Eds.), The Biology and Chemistry of the Compositae, 1: 269-81. London, Academic Press. [ Links ]
145. SOLBRIG, O.T. 1993. Plant traits and adaptive strategies: Their role in ecosystem function. En: Schulze & Mooney (eds.), Biodiversity and Ecosystem Function. Berlin, Springer. Pp. 97-116. [ Links ]
146. SÖRENSEN, N.A., 1977. Polyacetylenes and conservatism of chemical characters in the Compositae. En: Heywood, Harborne & Turner (eds.), The Biology and Chemistry of the Compositae, 1: 385-409. London, Academic Press. [ Links ]
147. STEBBINS, G.L. 1977. Flowering Plants Evolution above the species level. Cambridge, Harvard Univ. Press. 399 pp. [ Links ]
148. STEVENS, P.F., 2007. Angiosperm Phylogeny Website. Version 8, June 2007. En: http://www.mobot.org/MOBOT/research/APweb/orders/ asteralesweb. htm#Asteraceae. Consultado 27.III.2008. [ Links ]
149. STUESSY, T.F. & D. GARVER, 1996. The defensive role of pappus in heads of Compositae. En: Caligari & Hind (eds.), Compositae:Biology and Utilization. Kew, Royal Bot. Gardens. Pp. 81-91. [ Links ]
150. STUESSY, T.F., T. SANG & M.L. DEVORE, 1996. Phylogeny and biogeography of the subfamily Barnadesioideae with implications for early evolution of the Compositae. En: Hind & Beentje (eds.), Compositae: Systematics. 1. Kew, Royal Bot. Gardens. Pp. 463-90. [ Links ]
151. SUNDBERG, S. & T.F. STUESSY, 1990. Isolating mechanisms and implications for modes of speciation in Heliantheae (Compositae). Pl. Syst. Evol. Suppl. 4: 77-97 [ Links ]
152. THORNE, R.F. & J.L. REVEAL, 2007. An updated classification of the class Magnolipsida ("Angiospermae"). Bot. Rev. 73(2): 67-182. [ Links ]
153. TORRES ROMERO, J.H., 1983. Contribución al conocimiento de las plantas tánicas registradas en Colombia. Biblioteca J.G. Triana nº 2: 1-175.. Bogotá, Univ. Nac. Colombia/Colciencias. [ Links ]
154. TORRES ROMERO, J.H., 1983b. Contribución al conocimiento de las plantas tintóreas registradas en Colombia. Biblioteca J.G. Triana nº 3: 1-205. Bogotá, Univ. Nac. Colombia/Colciencias [ Links ]
155. TSEVEGSÜREN, N., K. AITZETMÜLLER, L. BRÜHL & G. WERNER, 2000. Seed oil fatty acids of Mongolian compositae. J. High Resol. Chromatogr. 23 (5): 360-66. [ Links ]
156. UPHOF, J.C.T, 1968. Dictionary of economic plants. Lehrer, Neth., Cremer. 591 pp. [ Links ]
157. UPHOF , J.C.T. & K. HUMMEL, 1962. Plant hairs. En: Handbook der Pflanzenanatomie 4 (5). Berlin, Gebr. Bornträger. Pp. 1-292. [ Links ]
158.USHER, G., 1974. A dictionary of plants used by man. London, Constable. 619 pp. [ Links ]
159. VAN DER Pihl, L., 1982. Principles of Dispersal in Higher Plants. 3º ed. Berlin/New York, Springer. 214 pp. il. [ Links ]
160. VAN DER VOSSEN, H.A.M. & G.S. MKAMILO, 2007. Plant Resources of Tropical Africa (PROTA), Vol. 14: Vegetable oils. Nairobi, Kenia, PROTA Foundation. 84 pp. [ Links ]
161. VAN WYK, B.-E., 2000. People's Plants, A Guide to the Useful Plants of Southern Africa. Pretoria, Briza Publ. 351 pp. [ Links ]
162. VAUGHAN, J.G., 1970. The Structure and Utilization of Oil Seeds. London, Chapman & Hall. 192 pp. [ Links ]
163. VAVILOV, N.I., 1926/1951. Estudios sobre el origen de las plantas cultivadas. Buenos Aires, Acme. xxii, 185 pp. il. [ Links ]
164. VOGEL, S., 1990a. Radiación adaptativa del síndrome floral en las familias neotropicales. Bol. Acad. Nac. Cs. Córdoba 59 (1-2): 5-30. [ Links ]
165. VOGEL, S., 1990b. The role of scent glands in pollination. On the structure and function of Osmophores. Washington, Smithsonian Inst./NSC. 203 pp. [ Links ]
166. WAGNER, H., 1977. Pharmaceutical and economic uses of the Compositae. En: Heywood, Harborne & Turner (eds.), The Biology and Chemistry of the Compositae, 1: 411-33. London, Academic Press. [ Links ]
167. WATANABE, K., 2008. Index to Chromosome Numbers in Asteraceae. En: htpp://www.asteraceae.cla.kobe-u.ac.jp./index.html. Consultado 22.XII.2008. [ Links ]
168. WATANABE, K. & S. SMITH-WHITE, 1987. Phyletic and evolutionary relationships of Brachyscome lineariloba (Compositae). Plant Syst. Evol. 157: 121-41. [ Links ]
169. WEBERLING, F., 1989. Morphology of flowers and inflorescences. Cambridge, Univ. Press. xx, 405 pp. il. [ Links ]
170. WEISS, E.A. (ed.), 1999. Oilseed Crops. 2º ed. Oxford, Blackwell Sci. 364 pp. [ Links ]
171. WHALEY, W.G., 1947. Russian dandelion (kok-saghyz) an emergency source of natural rubber. Washington, U.S. Gov. Print. Office. Misc. Publ. n° 618. [ Links ]
172. WICKENS, G.E., 2004. Economic Botany. Principles and Practices. Dordrecht, Boston, Kluwer Acad. Publ. 556 pp. [ Links ]
173. WILKES, G., 1977. Nature crops and wild food plants. Ecologist 7: 312-17. [ Links ]
174. WORWOOD, V.A., 1991. The Complete Book of Essential Oils and Aromatherapy. Novato, Cal., New World Library. 426 pp. [ Links ]
175. ZULOAGA, F.O. & O. MORRONE (eds.), 1999. Catálogo de las plantas vasculares de la República Argentina, II. Dicotyledoneae. Monogr. Syst. Bot. Missouri Bot. Gard. 74: xxii, 1-1269. [ Links ]
176. ZULOAGA, F.O., O. MORRONE & D. RODRÍGUEZ, 1999. Análisis de la biodiversidad en plantas vasculares de la Argentina. Kurtziana27: 17-167. [ Links ]
Recibido: 5/2009
Aceptado: 11/2009

