










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkMultequina
versión On-line ISSN 1852-7329
Multequina vol.19 no.2 Mendoza dic. 2010
ARTÍCULO ORIGINAL
Echinopsis leucantha (Gillies ex Salm-Dyck) Walp. (Cactoideae). Interacciones con plantas nodrizas en el desierto central argentino
Echinopsis leucantha (Gillies ex Salm-Dyck) Walp. (Cactoideae). Interactions with nurse plants in the central argentine desert
Martín Almirón1 y Eduardo Martínez Carretero1
1Geobotánica y Fitogeografía IADIZA (CONICET) CC 507, 5500 Mendoza - Argentina malmiron@mendoza-conicet.gob.ar
RESUMEN
En el presente estudio se examinó la distribución espacial de E. leucantha mediante el análisis de patrones espaciales bi-variados, en relación a la disponibilidad de sitios de instalación bajo los arbustos en tres unidades fisiográficas diferentes (geotopos). Se analizó la influencia de los arbustos en la radiación fotosintéticamente activa y en la temperatura a nivel del suelo. A escalas inferiores (0-1,6m) E. leucantha se encontró asociado en los tres geotopos a Larrea cuneifolia y/o a Zuccagnia punctata. Respecto a estos arbustos, la asociación encontrada mostró una alta relación con el gradiente de radiación solar fotosintéticamente activa encontrado desde el centro hacia la periferia de los arbustos. En relación a otros arbustos, la distribución fue aleatoria. El efecto de los arbustos en la temperatura del suelo no influyó de modo claro en la distribución espacial de E. leucantha.
Palabras clave: Asociación interespecífica; Heterogeneidad ambiental; Wiegand- Moloney's O(r) statistic
Summary
In this study we examined the spatial distribution of E. leucantha, through the analysis of bivariate spatial patterns, with respect to available installation sites under bushes in three different physiographic units (geotopes). The influence of shrubs in the photosynthetic active radiation and temperature at ground level was evaluated. A lower scales (0 to 1.6 m) E. leucantha was associated in three geotopes with Larrea cuneifolia and/or Zuccagnia punctata. Regarding to these shrubs, the association found showed a high relation to the gradient of photosynthetic active solar radiation found recorded from the center to the periphery of the bushes. In relation to other shrubs the distribution was random. The effect of shrubs on soil temperature did not influence in the spatial distribution of E. leucantha.
Key words: Interespecific associations; Environmental heterogeneity; Wiegand- Moloney's O(r) statistic
INTRODUCCIÓN
Muchas cactáceas de sistemas áridos y semiáridos se desarrollan bajo el dosel de los arbustos ya que éstos atenúan la radiación solar y consecuentemente la temperatura del suelo. Además, los arbustos ofrecen refugio contra herbívoros y suelen mejorar el contenido de nutrientes en el suelo (Turner et al., 1966, Jaksic & Fuentes, 1980; Valiente-Banuet et al., 1991, Valiente-Banuet & Excurra, 1991, Zuñiga et al., 2005). Por otro lado, se han detectado efectos negativos (alelopatía, competencia por recursos) entre los arbustos y las cactáceas (McAuliffe, 1984; Franco & Nobel, 1989; Flores-Martínez et al., 1994; Cazón et al., 2000; Cazón et al., 2002). El balance dinámico entre estas interacciones y los mecanismos de dispersión determinan la distribución de las cactáceas en estos ambientes.
López & Valdivia (2007) sugieren que las cactáceas columnares (no opuntioideas) de la Prepuna boliviana se desarrollan casi exclusivamente bajo arbustos, mientras que las de tipo opuntioideo pueden desarrollarse en áreas descubiertas. Las de tipo globoso serían indiferentes a los arbustos.
Teniendo en cuenta que la estructura del estrato arbustivo (considerado como sitio potencial de instalación) depende de la heterogeneidad de ambientes a escala de paisaje, se evaluó la distribución espacial de E. leucantha respecto a cada especie de arbusto encontrado en tres geotopos diferentes mediante el análisis de patrones espaciales bi-variados propuesto por Wiegand & Moloney (2004).
En cada geotopo se registró la radiación fotosintéticamente activa y la temperatura del suelo, bajo y entre los arbustos, con el fin de caracterizar los posibles sitios de instalación.
MATERIAL Y MÉTODO
ÁREA DE ESTUDIO
El estudio fue realizado en la localidad de Matagusanos (31°13'17"S—68°39'7"W) provincia de San Juan, Argentina. La precipitación media anual es de 116,5mm, (el 72% durante el período estival). La temperatura promedio anual es de 17,2º C, la temperatura máxima absoluta de 42,8º C y la mínima absoluta de -7,3º C. (Servicio Meteorológico Nacional, 1958). La fisonomía dominante es la de matorral, con cobertura media del 20%. Las especies dominantes son Larrea cuneifolia Cav., Bulnesia retama Cav., Zucaggnia punctata Cav., Larrea divaricata Cav. y Bougainvillea spinosa (Cav.) Heimerl., entre otras. A la escala de trabajo el paisaje es heterogéneo con planicies y laderas de exposición E y W, siempre con intensos procesos erosivos hídricos.
Echinopsis leucantha (Gillies ex Salm-Dyck) Walp., es una cactácea de tipo columnar característica de la Provincia Fitogeográfica del Monte (Kiesling, 2003) que alcanza <100cm de altura y <25cm de diámetro. De septiembre a marzo desarrolla flores blancas infundibuliformes. Los frutos son dehiscentes, con pulpa dulce y numerosas semillas negras y rugosas de 9,45 ±0,35mg de peso.
DISEÑO DEL MUESTREO
La distribución espacial de E. leucantha se evaluó en los tres geotopos dominantes: Llanura aluvial (A), Ladera de exposición Este (E) y Ladera de exposición Oeste (W) (Tabla 1).
Tabla 1. Caracterización ambiental de cada geotopo y especies arbustivas encontradas
Table 1. Environmental characterization of each geotope and shrubby species found

En cada geotopo se establecieron aleatoriamente ocho parcelas de 6x10m. Cada parcela se dividió en 1500 cuadros para obtener las coordenadas cartesianas de cada cactus como así también la forma, posición y tamaño de cada arbusto según la metodología propuesta por Wiegand et al. (2006) para análisis espacial de patrones con forma y tamaño finito.
Para interpretar el micro ambiente bajo los arbustos se registró la radiación fotosintéticamente activa (PAR) (radiómetro digital: QMSS-S, Apogee®) y la temperatura a 7cm del suelo (termómetros de alcohol de respuesta rápida), en los mismos arbustos y al mediodía en tres posiciones respecto de la proyección de sombra del arbusto: al centro del arbusto, al medio y al extremo de la proyección de su sombra, durante cuatro días representativos de las cuatro estaciones del año.
Con el fin de obtener una referencia del suelo descubierto, se promediaron los valores de PAR y temperatura tomados aleatoriamente en cuatro puntos sin cobertura vegetal dentro de cada parcela.
Para caracterizar al arbusto promedio de cada especie por geotopo se midió la altura y los diámetros mayor y menor de copa de todos los arbustos encontrados en las parcelas.
ANÁLISIS DE DATOS
Para determinar la diferencia media anual de temperatura y de radiación fotosintéticamente activa (ΔTemp y ΔPAR, respectivamente) bajo los diferentes arbustos en cada geotopo, se restaron los valores observados al valor medio de referencia en suelo descubierto de cada parcela, luego se promediaron los valores obtenidos en las cuatro estaciones. Se analizaron mediante ANOVA factorial y Test de Tukey (p<0,05) los ΔTemp y ΔPAR por geotopo entre las diferentes especies y posiciones respecto de la proyección de sombra.
Mediante el Software Programita® se analizó la distribución espacial de E. leucantha respecto a cada especie arbustiva, comparándose los valores de la función empírica Wiegand-Moloney's O(r) (Wiegand & Moloney, 2004) con un modelo nulo de distribución aleatoria tipo Poisson. Para determinar las bandas de confianza del 99%, se realizaron 99 simulaciones de Monte Carlo (Bailey & Gatrell, 1995; Haase, 1995; Martens, 1997) tomando el 5to mayor y el 5to menor valor como límites de confianza (Wiegand et al., 2006). Gráficamente, si la distribución empírica se observa dentro de los límites de confianza se asume que no hay diferencias significativas con el modelo de distribución Poisson aleatorio, cuando los valores observados se distribuyen sobre el límite superior indican atracción, mientras que los valores observados bajo el límite inferior indican repulsión (Diggle, 1983).
RESULTADOS
La composición florística, altura, cobertura media y dominancia específica de los arbustos fue diferente en los tres geotopos (Tabla 1).
En los tres ambientes se observó un gradiente de ΔTemp (A= F: 3,07; p<0,0484; E= F: 6,41; p:0,0019; W= F: 30,43; p<0,0001) y ΔPAR (A= F: 13,48; p<0,0001; E= F: 21,15; p<0,0001; W= F: 115,60; p<0,0001) diferente respecto de la posición, que disminuyó desde el centro de los arbustos hacia el extremo de la proyección de la sombra (Tabla 2).
Tabla 2. ANOVA de ΔTemp (ºC) y ΔPAR (μmol/m2*s) en los tres geotopos en función del factor "posición" Letras distintas indican diferencias significativas para Test de Tukey (p<0,05)
Table 2. ANOVA of ΔTemp (ºC) and ΔPAR (μmol/m2*s) in the three geotopes in function of "position" factor Different letters indicate significant differences for Tukey Test (p<0.05)

El factor "especie" resultó significativo para ΔTemp en todos los geotopos (A= ΔTemp: F: 24,19; p<0,0001; E= F: 10,18; p=0,0001; W= F: 8,81; p=0,0032) y para ΔPAR sólo en la llanura aluvial (F: 3,25; p<0,022). En la Llanura aluvial bajo B. retama el suelo estaba significativamente más fresco y sombrío (Tabla 3).
Tabla 3. ANOVA de ΔTemp (ºC) y ΔPAR (μmol/m2*s) en los tres geotopos en función del factor "especie" Letras distintas indican diferencias significativas para Test de Tukey (p<0,05)
Table 3. ANOVA of ΔTemp (ºC) y ΔPAR (μmol/m2*s) in the three geotopes in function of "specie" factor different letters indicate significant differences for Tukey Test (p<0.05)
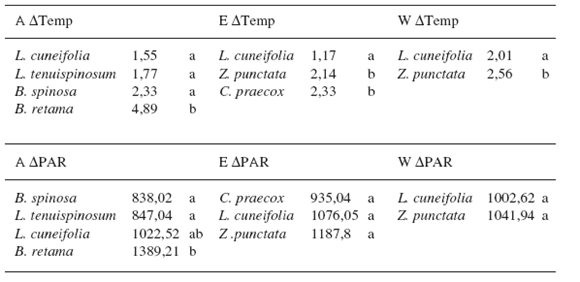
En (E) y (W) la especie más sombría fue Z. punctata, aunque sin diferencias con las otras arbustivas (Tabla 3). Respecto a ΔTemp, en (W) Z. punctata mostró el mayor valor, significativamente diferente de L. cuneifolia. En (E) las especies que generaron microambientes más frescos fueron C. praecox y Z. punctata (Tabla 3).
En todos los geotopos E. leucantha se asoció positivamente a algún arbusto: Con L. cuneifolia en A y en W, entre 0-1m de distancia y con Z. punctata entre 0-1,6m de distancia en E y entre 0-1,2 m en W. Respecto a otros arbustos (en A= B. spinosa, B. retama y L. tenuispinosum; en E= L. cuneifolia y C. praecox), no hubo evidencia significativa para rechazar el modelo de distribución Poisson aleatorio. Por último, en E se encontraron menos cactus que los esperados por el modelo Poisson aleatorio respecto a L. cuneifolia en distancias superiores a los 2,6m y respecto a Z. punctata entre los 1,6 y 2m en W (Figura 1).

Figura 1. Análisis de la distribución espacial de E. leucantha respecto a los arbustos encontrados en los tres geotopos: Llanura aluvial (A), ladera oeste (W) y ladera este (E). En trazo grueso se observa la función empírica Wiegand-Moloney's O(r) y en trazo fino el intervalo de confianza generado por el 5to mayor y menor valor de 99 simulaciones de Monte Carlo del modelo nulo Poisson aleatorio
Figure 1. Spatial distribution analysis of E. leucantha respect to shrubs found in the three geotopes: alluvial plain (A), West hillside (W), and East hillside (E). Bold line shows the empirical function Wiegand-Moloney's O(r), and thin lines show the confidence interval generated by the 5th highest and lowest value of 99 Monte Carlo simulations of random Poisson null model
DISCUSIÓN Y CONCLUSIONES
Las diferencias físicas entre los geotopos (pendiente, cobertura, exposición) y entre los arbustos (tamaño, cobertura, dominancia), ofrecieron una alta heterogeneidad ambiental para la instalación y desarrollo de E. leucantha. El efecto de los arbustos atenuando la radiación solar y la temperatura del suelo fue similar a lo encontrado por otros autores (Franco & Nobel, 1989; Reyes-Olivas et al., 2002). El gradiente microclimático observado desde el centro hacia la periferia de los arbustos fue el esperado, considerando que el centro del arbusto suele ser la posición más protegida, o menos expuesta.
El patrón de distribución de E. leucantha respecto a L. cuneifolia y Z. punctata evidencia la fuerte agregación espacial con estos arbustos, similar a lo encontrado por otros autores para otras cactáceas de tipo globoso y columnar (Valiente-Banuet & Ezcurra, 1991; de Viana et al., 2000; Mandujano et al., 2002; Reyes Olivas et al., 2002; López & Valdivia, 2007). Posiblemente esta asociación se deba a que L. cuneifolia y Z. punctata atenúan la radiación solar en rangos intermedios (1002,62—1187,8 μmol/m2.s) comparados con otros arbustos que filtran más (B. retama) o menos (B. spinosa, L. tenuispinosum, C. praecox) radiación solar. La temperatura bajo las diferentes especies arbustivas no influyó de modo claro en distribución de E. leucantha. Quizás esta temperatura se relacione a otras variables independientes de los arbustos, como la velocidad del viento, corrientes convectivas locales, tipo y color de suelo, etc.
Si bien Méndez et al. (2004), aplicando el test G de presencia/ausencia, halló asociación positiva entre ésta cactácea y arbustos del género Larrea, la aplicación de modelos bi-variados permitió interpretar la distribución de E. leucantha como un proceso dependiente de la escala espacial. Los valores empíricos de la función Wiegand-Moloney's O(r), que evidencian asociación positiva con los arbustos (Figura 1), decrecen a medida que aumenta la escala espacial considerada hasta valores próximos a los diámetros menores de los arbustos asociados (Tabla 1). Esta asociación coincide en general con el gradiente de ΔTemp y ΔPAR encontrado bajo los arbustos. A escalas mayores no se puede rechazar la hipótesis de distribución Poisson aleatoria, pues la curva empírica se ubicó entre los intervalos de confianza (Figura 1). Por ello, se puede concluir que E. leucantha además de encontrarse preferentemente bajo L. cuneifolia y Z. punctata, es más probable encontrarlo en el centro del arbusto que hacia la periferia. Estos resultados enfatizan la necesidad de estudiar las posibles asociaciones planta-nodriza a nivel específico (Arriaga et al., 1993; de Viana et al., 2000), considerándolas como procesos dependientes de la escala.
Las asociaciones encontradas en este trabajo pueden ser el resultado de la atenuación de la radiación solar bajo L. cuneifolia y Z. punctata sumado a mecanismos de dispersión diferencial intermediados por frugívoros, como ocurre con otras cactáceas con frutos de características similares (Steenbergh & Lowe, 1977; Silva, 1988; Wendelken & Martin, 1988; Soriano et al., 1991; Silvius, 1995; de Viana et al., 2000). Además, debe tenerse en cuenta que las semillas son fotoblásticas positivas y la aparente dormancia de E. leucantha (Ortega-Baes et al., 2010) puede influir en el modelo de distribución general actual.
AGRADECIMIENTOS
A Mariano Ariza, Juan Pablo Cáceres J. y Juan Aguilera por la ayuda en las tareas de campo. A los revisores anónimos por las sugerencias.
BIBLIOGRAFÍA
1. CAZÓN, A., M. DE VIANA & J. GIANELLO, 2000. Identificación de un compuesto alelopático de Baccharis boliviensis (Asteraceae) y su efecto en la germinación de Trichocereus pasacana (Cactaceae). Biol. Trop. 48: 47-51. [ Links ]
2. CAZÓN, A., M. DE VIANA & J. GIANELLO, 2002. Comparación del efecto fitotóxico de aleloquímicos de Baccharis boliviensis (Asteraceae) en la germinación de Trichocereus pasacana (Cactaceae). Ecología Austral 12: 73-78. [ Links ]
3. DE VIANA, M., S. SUHRING & B. MANLY, 2000. Application of randomization methods to study the association of Trichocereus pasacana (Cactaceae) with potential nurse plants. Plant Ecol. 156:193-197. [ Links ]
4. DIGGLE, P., 1983. Statistical analysis of spatial point patterns. Academic Press, London. [ Links ]
5. FRANCO, A. & P. NOBEL, 1989. Effect of nurse plants on the microhabitat and growth of cacti. J. Ecol. 77: 870-886. [ Links ]
6. FLORES-MARTÍNEZ, A., E. EZCURRA & S. SÁNCHEZ-COLÓN, 1994. Effect of Neobuxbaumia tetetzo on growth and fecundity of its nurse plant Mimosa luisiana. J. Ecol. 82: 325-330. [ Links ]
7. JAKSIC, F. & E. FUENTES, 1980. Why are native herbs in the Chilean matorral more abundant beneath bushes: microclimate or grazing? J. Ecol. 68: 665-669. [ Links ]
8. KIESLING, R., 2003. Flora de San Juan, República Argentina, Vol II. Estudio Sigma Editores. [ Links ]
9. LÓPEZ, R. & S. VALDIVIA, 2007. The importance of shrub cover for four cactus species differing in growth form in an Andean semi-desert. J. Veg. Sci.18: 263-270. [ Links ]
10. MCAULIFFE, J., 1984. Sahuaronurse tree associations in the Sonoran Desert: competitive effects of Sahuaros. Oecología 64. 319-321. [ Links ]
11. MANDUJANO, M., A. FLORES MARTINEZ, J. GOLUBOV & E. EZCURRA, 2002. Spatial distribution of three globose cacti in relation to different nurse-plant canopies and bare areas. The Southwestern Naturalist 47(2):162-168. [ Links ]
12. MÉNDEZ, E., J. GUEVARA & O. ESTEVEZ, 2004. Distribution of cacti in Larrea spp. shrublands in Mendoza, Argentina. J. Arid Environ. 58: 451-462. [ Links ]
13.ORTEGA-BAES, P., M. APARICIO-GONZÁLEZ, G. GALÍNDEZ, P. DEL FUEYO, S. SÜHRING & M. ROJAS-ARÉCHIGA, 2010. Are cactus growth forms related to germination responses to light? A test using Echinopsis species. Acta Oecologica. 36: 339-342. [ Links ]
14. REYES-OLIVAS, A., E. GARCÍAMOYA & L. LÓPEZMATA, 2002. Cacti-shurb interaction in the coastal desert of northern Sinaloa, Mexico. J. Arid Environ. 52: 431-445. [ Links ]
15. SERVICIO METEOROLÓGICO NACIONAL, 1958. Estadísticas climatológicas 1941-1950. Publicación B1. [ Links ]
16. SILVA, W.,1988. Ornitocoria em Cereus peruvianus (cactaceae) na Serra do Japi, estado de Sao Paulo. Revista Brasileira de Biologia 48 (2):381-389. [ Links ]
17. SILVIUS, K., 1995. Avian consumers of cardon fruits (Stenocereus griseus: Cactaceae) on Margarita Island, Venezuela. Biotropica 27(1): 96-105. [ Links ]
18. SORIANO, P., M. SOSA & O. ROSSELL, 1991. Hábitos alimentarios de Glossophaga longirostris Miller (Chiroptera: Phyllostomidae) en una zona árida de Los Andes venezolanos. Revista de Biología Tropical 39(2):263-268. [ Links ]
19. STEENBERGH, W. & C. LOWE, 1977. Ecology of the Sahuaro: II, Reproduction, Germination, Establishment, Growth and survival of the young plant. National Park Service Scientific Monograph 8, Gob. Printing Office, Washington, D.C. [ Links ]
20.TURNER, R., S. ALCORN, G. OLIN & J. BOOTH, 1966. The influence of shade, soil and water on Saguaro seedling establishment. Bot. Gaz.127: 95-102. [ Links ]
21. VALIENTE-BAUNET, A., A. BOLOGNA ROCRAVENNA, O. BRIONES, E. EZCURRA, M. ROSAS, H. NÚÑEZ, G. BARNARD & E. VAZQUEZ, 1991. Spatial relation between cacti and nurse shrubs in a semi-arid Environment in central Mexico. Journal of Vegetation Science. 2 (1): 15-20. [ Links ]
22. VALIENTE-BAUNET, A. & E. EZCURRA, 1991. Shade as a cause of the association between the cactus Neobouxbaumia tetetzo and the nurse plant Mimosa luisana in the Tehuacan Valley, México. J Ecol. 79: 961-971. [ Links ]
23. WENDELKEN, P & R. MARTIN, 1988. Avian consumption of the fruit of the cacti Stenocereus eichlamii and Pilosocereus maxonii in Guatemala. American Midland Naturalist 119:235-243. [ Links ]
24. WIEGAND, T. & K. MOLONEY, 2004. Rings, circles and nullmodels for point pattern analysis in ecology. Oikos 104: 209-229. [ Links ]
25. WIEGAND, T., W. KISSLING, P. CIPRIOTTI & M. AGUIAR, 2006. Extending point pattern analysis to objects of finite size and irregular shape. J. Ecol. 94: 825-837. [ Links ]
26. ZUÑIGA, B., G. MALDA & H. SUZÁN, 2005. Interacciones Planta - nodriza en Lophophora diffusa (Cactaceae) en un desierto subtropical de México. Biotrópica 37: 351-356. [ Links ]
Recibido: 6/2010
Aceptado: 9/2010

