










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
On-line version ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.11 no.2 Ciudad Autónoma de Buenos Aires Dec. 2009
PALEONTOLOGÍA
Primeros registros fósiles de pejerreyes (Teleostei: Atheriniformes) en el Pleistoceno Medio de la provincia de Buenos Aires, Argentina
Sergio Bogan1, Martín L. de los Reyes2 & Marcos M. Cenizo1,2
1Área Paleontología. Fundación de Historia Natural "Félix de Azara". Departamento de Ciencias Naturales y Antropología. CEBBAD - Universidad Maimónides. Valentín Virasoro 732 (C1405BDB) Ciudad Autónoma de Buenos Aires, Argentina. sergiobogan@yahoo.com.ar.
2Facultad de Ciencias Naturales y Museo, UNLP. Museo de La Plata. Paseo del Bosque, s/nº (1900). La Plata, Buenos.
delossreyes@yahoo.com.ar; cenizomarcos@yahoo.com.ar
Resumen: En este trabajo se dan a conocer restos fósiles de pejerreyes recuperados en secuencias sedimentarias correspondientes al Piso-Edad Bonaerense (Pleistoceno Medio) de la localidad de Centinela del Mar, provincia de Buenos Aires, Argentina. Los únicos registros fósiles previos de pejerreyes sudamericanos se restringen a tres localidades de edad Miocena de Argentina y Chile. La presencia de diversos caracteres osteológicos diagnósticos permitió referir al género Odontesthes algunas de las estructuras óseas aquí tratadas. Estos registros brindan nuevos datos acerca de la composición de las ictiofaunas pleistocénicas de Argentina, constituyendo los primeros restos fósiles de pejerreyes procedentes de la región pampeana.
Palabras clave: Pejerreyes; Atherinomorpha; Sorgentinini; Odontesthes; Pleistoceno; Centinela del Mar; Argentina.
Abstract: First fossil records of silversides (Teleostei: Atheriniformes) from the Middle Pleistocene of Buenos Aires province, Argentina. This paper presents the fossil remains of silversides recovered in sedimentary sequences corresponding to Centinela del Mar district Bonaerian Stage-Age (Middle Pleistocene), Buenos Aires province, Argentina. The few only previously described fossil silversides are restricted to three Chilean and Argentinean Miocene localities. The silversides studied here present several diagnostic osteological characters that allow us to refer the remains to the genus Odontesthes. These records give us new data on the composition of the pleistocenic ictiofaunas of Argentina, besides representing the first fossil records of silversides from the Pampean region.
Key words: Silversides; Atherinomorpha; Sorgentinini; Odontesthes; Pleistocene; Centinela del Mar; Argentina.
INTRODUCCIÓN
Los pejerreyes distribuidos actualmente en el continente americano son peces Atheriniformes incluidos en la familia Atherinopsidae. Dicha familia está integrada por las subfamilias Menidiinae y Atherinopsinae, ambas endémicas de las aguas continentales y marinas.
La subfamilia Atherinopsinae fue establecida por Fowler en 1903 (Schultz, 1948) y agrupa a la mayoría de los pejerreyes del Nuevo Mundo. Esta subfamilia cuenta con dos tribus: Atherinopsinini (Atherinops, Atherinopsis, Colpichthys y Leuresthes) distribuida en América del Norte y Sorgentinini (Basilichthys y Odontesthes) en América del Sur (White, 1985; Dyer, 1997, 1998, 2006). De acuerdo a la hipótesis filogenética planteada por Dyer (1997) ambas tribus serían monofiléticas.
El género Odontesthes Evermann & Kendall 1906, de gran distribución geográfica y amplitud ecológica, incluye al menos 19 especies, con representantes tanto de aguas marinas litorales y estuarinas como así también dulceacuícolas (Dyer, 2000, 2006). Las formas marinas se distribuyen desde el extremo austral de Tierra del Fuego hasta Piura (Perú) por el Pacífico, y hasta Santos (sur de Brasil) por el Atlántico, incluyendo las costas de las Islas Malvinas. Las formas de agua dulce se extienden desde Patagonia (río Santa Cruz, Argentina) hasta el norte del estado de Río Grande do Sul (Brasil) (White, 1985; Dyer, 1998, 2000, 2003, 2006).
Basilichthys (Girard, 1855), el género hermano de Odontesthes, cuenta con cinco especies, todas dulceacuícolas, distribuidas en ríos y lagos al oeste de los Andes desde Lambayeque (Perú) hasta la isla de Chiloé (Chile) (Dyer, 2006).
El criterio biogeográfico seguido por Dyer (1998, 2000, 2006) indica que la tribu Sorgentinini sería un grupo primitivamente dulceacuícola, constituyendo el ambiente marino una condición derivada para el grupo. Esta apreciación se fundamenta en el hecho de que las especies del género Basilichthys, como así también las especies basales de Odontesthes, son dulceacuícolas. Tanto las especies incluidas en el género Odontesthes como aquellas reunidas en el género Basilichthys constituirían grupos monofiléticos (Dyer, 1997).
El registro fósil de pejerreyes sudamericanos es sumamente exiguo. Su presencia ha sido referida sólo para tres localidades de edad miocena: Puesto Galván en Argentina (Bocchino, 1971), y Cerro La Mina y El Tallón en Chile (Rubilar, 1994).
El objetivo del presente trabajo es dar a conocer nuevos materiales asignados al género Odontesthes recuperados en secuencias sedimentarias asignables a la parte más alta del Piso-Edad Bonaerense (Pleistoceno Medio, sensu Cione& Tonni, 2005) de la localidad de Centinela del Mar, provincia de Buenos Aires, Argentina.
MATERIALES Y MÉTODOS
El esquema lito-bioestratigráfico adoptado en este trabajo es el propuesto por Cione & Tonni (1999, 2005). El concepto aplicado de "litofacies" corresponde a Reading & Levell (1996).
Las medidas fueron tomadas con calibre de resolución 0,01 mm. Las comparaciones osteológicas fueron efectuadas siguiendo la bibliografía específica (White, 1985; Dyer, 1997, 2006) y asistidas por ejemplares osteológicos de las siguientes especies actuales de pejerreyes: Odontesthes bonariensis (6), O. argentinensis (5), O. perugiae (10), O. incisa (3), O. humensis (2), O. nigricans (3), O. smitti (2) y O. hatcheri (4). Los materiales comparativos antes citados pertenecen a la colección personal de uno de los autores (S.B.).
Abreviaturas. LS, longitud estándar; LT, longitud total; MLP, Colección División Paleontología de Vertebrados, Museo de La Plata, La Plata, Argentina.
GEOLOGÍA Y EDAD
La localidad de Centinela del Mar (S 38º 26´ - O 58º 14´) se ubica geográficamente en el litoral atlántico bonaerense (Fig. 1), partido de General Alvarado, provincia de Buenos Aires. Aquí se levanta un acantilado litoral con una extensión aproximada de 2,85 km cuya altura fluctúa entre 5 y 15 metros. El mismo es limitado al sudoeste por la confluencia de los arroyos El Pescado, Nutria Mansa y Malacara. La sucesión estratigráfica es relativamente homogénea, al menos en sus niveles superiores, y en todo el perfil se destaca la presencia de "horizontes guías" de posible extensión regional. En la secuencia expuesta se encuentra representado un conjunto de litofacies referibles al intervalo Plioceno Tardío-Pleistoceno Tardío (de los Reyes et al., 2006).
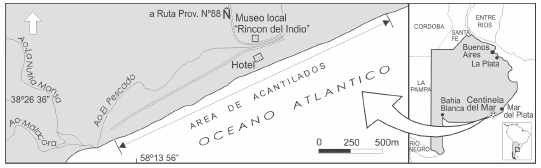
Fig. 1. Ubicación geográfica de la localidad Centinela del Mar
Los materiales aquí descriptos provienen de dos litofacies diferentes, ubicadas verticalmente en el sector medio del acantilado (Litofacies G e I, Fig. 2).
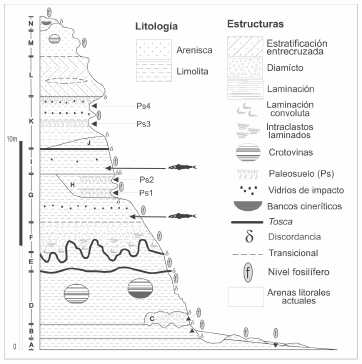
Fig. 2. Perfil estratigráfico integrado de la localidad de Centinela del Mar
Sólo una vértebra (MLP 04-V-2-227) de pejerrey fue recuperada de la litofacies G. Dicha unidad está representada por 1 a 2 m de limos loessoides arenosos a areno-arcillosos castaños oscuros y masivos. La litofacies G integra una sucesión facial con la litofacies infrayacente (F), vinculada a fenómenos de remoción en masa, correspondiente a una dinámica de tipo torrencial con una disminución de la energía depositacional hacia el techo de la sucesión.
El resto de los ejemplares fueron exhumados de la litofacies I. Dicha unidad se encuentra representada por limos loessoides arcillosos, verdes a verde-grisáceos, muy plásticos, cuya potencia fluctúa entre los 0,5 y 2 m de espesor. Los mismos incluyen pequeños rodados líticos (<1mm), como así también restos escasos de vidrios de impacto. En la mayor parte de la secuencia expuesta, una inconformidad erosiva separa la citada unidad I de la inferior (G), no obstante, en el extremo noreste de la exposición la litofacies I descansa sobre la H (Fig. 2). Por otra parte, el techo de la unidad I posee un desarrollo perfectamente horizontal en todo el perfil, y se halla coronado por mantos de concreciones calcáreas nodulares y tabulares. Sobre el mismo se encuentra depositada la unidad K en franca discordancia erosiva, con excepción de un acotado sector de la barranca (sector del hotel) en donde la unidad J descansa sobre la unidad I. La génesis de este depósito estaría vinculada a llanuras de inundación, con la presencia de cuerpos lénticos permanentes, algunos de relativa profundidad. Esta idea es sustentada tanto por la evidencia sedimentológica como por la naturaleza tafonómica y taxonómica de la tafocenosis exhumada (Bogan et al., 2006; Bogan et al., 2009; Cenizo e Ibáñez, 2006).
Sobre la unidad sedimentaria I se encuentra depositada la unidad K interpretada como un ambiente de mezcla (Isla et al., 2000; Schultz et al., 2004; Bogan & Cenizo, 2008), representando facies estuariales de la transgresión marina Belgranense (=Fm. Pascua, sensu Isla et al., 2000), correlacionable con el interglacial Sangamoniano o Tirreniano alrededor de 125 ka (estadío isotópico 5e).
Cabe señalar que la secuencia de litofacies EN ha sido referida al cron de polaridad normal Brunhes, y por lo tanto más joven que 780 ka AP (Heil et al., 2002; Soibelzon et al., en prensa). De esta manera, las litofacies G e I estarían incluidas dentro del intervalo cronológico 780-125 ka correspondiente al Piso-Edad Bonaerense (Pleistoceno Medio, vease Cione & Tonni, 2005)
SISTEMÁTICA
Orden ATHERINIFORMES Rosen, 1964
Familia ATHERINOPSIDAE Fowler, 1903
Subfamilia ATHERINOPSINAE Fowler, 1903
Tribu SORGENTININI Pianta de Risso& Risso, 1953
Género Odontesthes Evermann & Kendall, 1906
Odontesthes sp.
Material referido. Premaxilar izquierdo (MLP 04-V-2-223; Fig. 3, K y L).
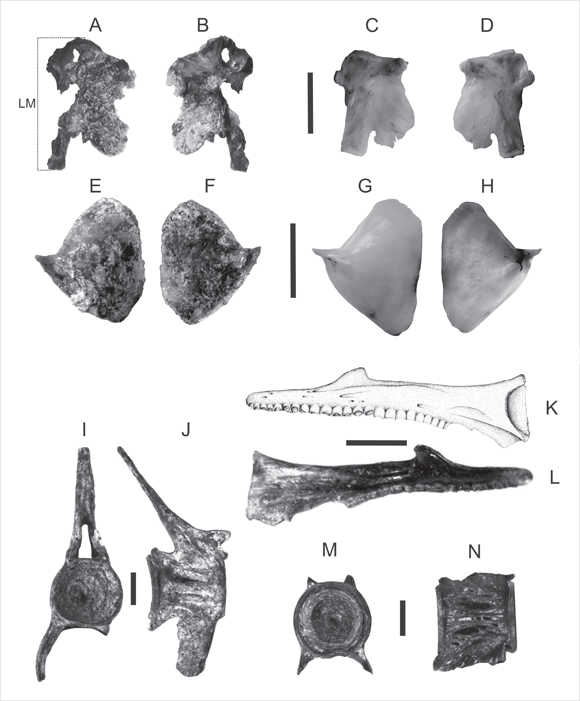
Fig. 3. Hiomandibular derecho (MLP 04-V-2-222) A: vista lateral externa, B: vista lateral interna. Hiomandibular derecho (comparativo) Odontesthes bonariensis C: vista lateral externa, D: vista lateral interna. Opérculo izquierdo (MLP 04-V-2-224), E: vista lateral externa, F: vista lateral interna. Opérculo izquierdo (comparativo) O. bonariensis G: vista lateral externa, H: vista lateral interna. Premaxilar (MLP 04-V-2-223), K: vista lateral externa, L: vista lateral interna. Vértebra caudal (MLP 04-V-2-226), I: vista cara frontal, J: vista cara lateral. Vértebra caudal (MLP 04-V-2-227), M: vista cara frontal, N: vista cara lateral. La escala equivale a 10 mm. en A-H y a 5 mm. en K-N.
Descripción. Se trata de un elemento curvo que cuenta con un proceso ascendente y un proceso posterior (o porción distal del brazo alveolar). El proceso ascendente presenta la base estrecha y su porción apical se halla incompleta. El proceso posterior de la rama del premaxilar, si bien se encuentra fracturado, exhibe un progresivo ensanchamiento hacia la porción distal. En la parte inferior del premaxilar se ubican varias filas de alvéolos donde los dientes se implantarían de forma algo desordenada.
Discusión. La condición dilatada del proceso posterior es considerada un carácter derivado para Atherinopsidae (Schultz, 1948; Dyer, 1997). El proceso ascendente con base estrecha es un carácter común para las especies del género Odontesthes (Dyer, 1997; Bemvenuti, 2005) y permite distinguirlo de las especies del género hermano Basilichthys. En las especies de este último género, la base del proceso ascendente es notablemente amplia debido a la reducida protractilidad de las mandíbulas (De La Hoz & Tosti- Croce, 1981; Dyer, 1997).
Material referido. Opérculo izquierdo (MLP 04-V-2-224; Fig. 3, E y F).
Descripción. Se trata de un hueso laminar que cuenta con un proceso dorsal. En el ángulo anterodorsal de la cara interna del opérculo se encuentran dos engrosamientos que convergen en el cóndilo hiomandibular. Es de destacar la presencia en este ángulo de por lo menos una fenestra visible desde la cara externa del opérculo. El borde dorsal del opérculo no es continuo con el flanco laminar del proceso dorsal y el borde posterodorsal es convexo.
Discusión. La presencia de un proceso dorsal es característica de los Atheriniformes (Dyer, 1997). En el opérculo descrito, el borde dorsal no es continuo con el flanco laminar del proceso dorsal, siendo ésta una condición derivada característica de los Atherinopsidae, con excepción de O. hatcheri, O. perugiae y O. retropinnis, quienes conservan el carácter plesiomórfico de los Atheriniformes (Dyer, 1997).
La manifestación de fenestras en el ángulo anterodorsal es considerada una sinapomorfía del género Odontesthes (White, 1985; Dyer, 1997) y está presente en todas las especies con excepción de O. incisa (White, 1985; Dyer, 1997) y O. perugiae (Bemvenuti, 2005). Es importante remarcar que ninguna de las especies del género Basilichthys presenta estas fenestras (White, 1985; Dyer, 1997).
La presencia de un borde posterodorsal convexo es considerada por Dyer (1997) como una condición derivada típica de las especies del género Odontesthes. En Basilichthys el borde posterodorsal es cóncavo (Dyer, 1997).
Material referido. Hiomandibular derecho (MLP 04-V-2-222; Fig. 3, A y B).
Descripción. El ejemplar MLP 04-V-2-222 es el hiomandibular más completo que pudo recuperarse en esta localidad. Cuenta con un proceso opercular, un proceso laminar metapterigóide y dos procesos superiores de articulación con los huesos esfenótico y pterótico del neurocráneo. La porción superior se encuentra muy ventifacteada y meteorizada, por hallarse aflorando en la barranca. En la cara ventral se encuentra parte del foramen superior cubierto por concreciones, y en el proceso inferior de la cara dorsal se observa claramente el foramen de la ramificación mandibular próximo al sector de unión del hiomandibular con el simpléctico. La parte izquierda del proceso inferior se halla rota sobre y paralelamente al canal de la ramificación hioidea del nervio hiomandibular, permitiendo observar parte del recorrido de dicho nervio e incluso el tabique donde se divide en dos ramas. Esto último nos permite verificar que la ramificación del nervio se produce en el interior del hueso.
La longitud máxima (LM) del ejemplar es de 22 mm, tomada desde el punto más elevado de la carilla de articulación del proceso posterior, en línea recta hasta la carilla distal de contacto con el hueso simpléctico del proceso inferior del hiomandibular (carilla simpléctica, Fig. 3 A).
Discusión. El nervio hiomandibular penetra por el foramen superior de la cara interna del hueso hiomandibular, dividiéndose en el interior de este hueso y emergiendo ramificado por la cara dorsal con una rama mandibular y una rama hioidea. Esta es una condición derivada para las especies del género Odontesthes, con excepción de O. hatcheri que al igual que las especies del género Basilichthys presenta la condición plesiomórfica, produciéndose la ramificación del nervio hiomandibular en el exterior del hueso (Dyer, 1997; Bemvenuti, 2005). La longitud del hueso hiomandibular registrada en el ejemplar MLP 04-V-2-222 denota que el mismo perteneció a un pez de gran porte. En la actualidad, la especie de Odontesthes que presenta mayor longitud es O. bonariensis alcanzando 520 mm de longitud estándar (Dyer, 2003). Creemos interesante señalar que un ejemplar de O. bonariensis de 384 mm de LS y 450 mm de LT, colectado en el Río de la Plata, presenta un hueso hiomandibular de 20,5 mm (la longitud del hueso fue tomada en los mismos puntos que en el ejemplar fósil). Con este ejemplo no pretendemos señalar una filiación con O. bonariensis, simplemente consideramos pertinente remarcar que el ejemplar MLP 04-V-2-222 debió pertenecer a un pejerrey muy grande (posiblemente mayor a 400 mm LS.).
Sorgentinini indet.
Material referido. Vértebras caudales (MLP 04-V-2-227, MLP 04-V-2-226, MLP 04-V-2- 231 y MLP 04-V-2-232; Fig. 3: I, J, M y N).
Descripción. Los elementos recuperados (MLP 04-V-2-227, MLP 04-V-2-226, MLP 04-V-2- 231 y MLP 04-V-2-232) permiten notar la presencia de modificaciones en los arcos hemales. Si bien ninguna de las piezas halladas se encuentra completa, es posible advertir una expansión de los arcos hemales y un notable ensanchamiento de las paredes de los mismos.
Discusión. Las expansiones advertidas conformarían parte de un embudo hemal. La presencia de un embudo hemal es característica de los pejerreyes de la subfamilia Atherinopsinae (Schultz, 1948). Por otra parte, el ensanchamiento de las paredes de los arcos hemales en las vértebras caudales es un carácter exclusivo de los pejerreyes sudamericanos Sorgentinini (White, 1985, Dyer 1997). A diferencia de estos últimos, las especies de los cuatro géneros de la tribu Atherinopsinini presentan un embudo hemal desarrollado a partir de la bifurcación de las apófisis de las espinas hemales (White, 1985, Dyer 1997).
El embudo hemal es una estructura especializada que contiene la porción terminal de la vejiga natatoria. Sólo O. incisa y O. nigricans carecen de la citada estructura (Pianta de Risso & Risso, 1953; De Buen, 1953; Dyer, 1997). No obstante, Bemvenuti (2005) identifica un embudo hemal en los ejemplares de O. incisa del sur de Brasil, aunque dicha estructura presenta en esta especie paredes laterales muy estrechas. Cabe aclarar que coincidimos con la observación de Bemvenuti, puesto que advertimos la presencia de un embudo hemal con paredes laterales muy estrechas en ejemplares de O. incisa colectados en la costa bonaerense.
Atheriniformes indet.
Material referido. Fragmento de hiomandibular derecho (MLP 04-V-2-243); fragmento de hiomandibular derecho (MLP 04-V-2-244); fragmento de frontal derecho (MLP 04-V-2-241); fragmento de frontal izquierdo (MLP 04-V-2-243); fragmento de cleitro (MLP 04-V-2-239); epihial derecho (MLP 04-V-2-240); 6 vértebras dorsales (MLP 04-V-2-233, MLP 04-V-2-234, MLP 04-V-2- 235, MLP 04-V-2-236, MLP 04-V-2-237 y MLP 04- V-2-238); 3 vértebras caudales (MLP 04-V-2-228, MLP 04-V-2-229 y MLP 04-V-2-230).
Discusión. Los materiales referidos a Atheriniformes indeterminados fueron cotejados con las especies actuales del género Odontesthes utilizadas como material comparativo, corroborándose una gran afinidad en las características morfológicas de ambos conjuntos. En este sentido, consideramos muy probable la correspondencia entre el material recuperado en Centinela del Mar y alguna especie del género Odontesthes. No obstante, no fue posible confirmar esta condición dado que los materiales analizados carecen de caracteres sinapomórficos, ya sea debido a su estado de fragmentación o al escaso interés que los estudios sistemáticos prestaron a los elementos anatómicos aquí considerados.
CONCLUSIONES
La presencia de diversos caracteres osteológicos diagnósticos permite referir al género Odontesthes parte de las estructuras óseas recuperadas. Si bien no fue posible una asignación específica, resulta importante destacar la presencia en el material estudiado de varios caracteres que permiten, en primera instancia, excluir la posibilidad de que se trate de las especies dulceacuícolas Odontesthes hatcheri, O. perugiae y O. retropinnis, como así tampoco las marinas O. incisa y O. nigricans.
La exclusión de O. hatcheri resulta particularmente importante debido a que se trata de la única especie dulceacuícola del género que habita en la actualidad la subregión Austral (Dyer, 1998). Según Dyer (2000) esta especie se encuentra distribuida desde el río Santa Cruz, (provincia de Santa Cruz, Argentina) hasta el río San Juan y Desaguadero (provincia de San Juan y Mendoza, respectivamente, Argentina). En la provincia de Buenos Aires se halla presente en la cuenca del río Colorado (Ringuelet et al., 1967).
De acuerdo a Dyer (1998), O. hatcheri se encuentra "aislada" en Patagonia desde el Mioceno Medio de las demás especies dulceacuícolas de abolengo brasílico.
En tal sentido, consideramos relevante señalar que la localidad de Centinela del Mar se ubica a 300 km al sur del río Salado, siendo en la actualidad O. bonariensis la única especie dulceacuícola propia de ríos y ambientes lénticos pampásicos con una amplia distribución en estas latitudes. Ringuelet (1975) señala que O. bonariensis es un pejerrey de gran tamaño típico de aguas templadas (que no sufren un excesivo calentamiento en el verano) con un rango de tolerancia de temperaturas entre los 0° y 34° C, con un óptimo para su normal desarrollo de 17° C. Durante los primeros años de vida, su espectro dietario se constituye principalmente de pequeños organismos planctónicos, complementado secundariamente por crustáceos, insectos y pequeños moluscos. Al alcanzar mayores dimensiones integra a su dieta pequeños peces.
Desde el punto de vista paleontológico, los únicos registros fósiles previos de pejerreyes sudamericanos se restringen a tres localidades de edad Miocena de Argentina y Chile.
La primera de ellas es Puesto Galván, departamento Cushamén, provincia de Chubut, Argentina. De esta localidad se describió un ejemplar casi completo referido por Bocchino (1971) a Basilichthys aff. regius. Es importante señalar que para el momento en que fue descripto dicho ejemplar, el conocimiento de la ictiofauna argentina presentaba grandes confusiones respecto de la correspondencia taxonómica de los pejerreyes que habitan este territorio. Tal confusión se debe, en gran medida, a una revisión realizada por Lahille (1929) quien refirió todas las especies vivientes de pejerreyes argentinos al género Basilichthys. Posteriormente, este criterio fue seguido y difundido por muchos ictiólogos y naturalistas argentinos (e.g. Pozzi & Bordalé, 1935; Marrero, 1950; Ringuelet et al., 1967, 1975). Dyer (1998) hace mención al material de Puesto Galván confirmando taxativamente que se trata de una especie del género Odontesthes señalando, además, la gran afinidad que presenta con O. hatcheri y O. retropinnis.
Un segundo taxón fue referido para la misma localidad al género de Perciformes Istiophorus sp. (Bocchino, 1971), pero una revisión posterior de carácter preliminar (Arratia & Cione, 1996), señala que dicho espécimen no corresponde al género Istiophorus y lo refiere a la familia "Atherinidae".
La segunda y tercera localidades se encuentran en el sur de Chile, éstas son Cerro La Mina y El Tallón. Los materiales fueron recuperados en unidades sedimentarias fluviolacustres correspondientes al Mioceno (Formación Cura-Mallín) y asignados a la familia "Atherinidae" (Rubilar, 1994). Es importante resaltar que Rubilar (1994) señala que los elementos óseos recuperados no permiten una asignación genérica por carecer de caracteres diagnósticos.
Los conocimientos sobre la dinámica paleobiogeográfica y paleoecológica de las comunidades ictícolas continentales del neógeno argentino son aún fragmentarios. De igual manera, las hipótesis filogenéticas actuales cuentan con escaso sustento en el registro fósil. Los restos aquí comunicados brindan nuevos datos acerca de la composición de las ictiofaunas pleistocénicas de Argentina, constituyendo los primeros ejemplares fósiles de pejerreyes procedentes de la región pampeana.
AGRADECIMIENTOS
En especial a A. Almirón y J. Casciotta por la bibliografía aportada, a F. Zorzi por las modificaciones gramaticales y sintácticas sugeridas, a U.F.J. Pardiñas por la lectura del manuscrito, a F. Agnolin por sus valiosos comentarios y a F. Goin por colaborar en la obtención de las imágenes. Nuestro agradecimiento se hace extensivo a A. C. Larriestra, E.P. Tonni, G. Scillato-Yané, S. Lorenzini, C. Canelo y familia de los Reyes, tanto como a la Fundación de Historia Natural "Félix de Azara", Universidad Maimónides. Este trabajo fue realizado en el marco del proyecto UNLP 1000-03215-2005.
BIBLIOGRAFÍA
1. Arratia, G. & A. Cione. 1996. The Record of Fossil Fishes of Southern South America. En: G. Arratia (ed.), Contributions of Southern South America to Vertebrate Paleontology. Müncher Geowissen. Abhand. Reihe A. Geol. Pal. 30: 9-72. [ Links ]
2. Bemvenuti, M.A. 2005. Osteologia comparada entre as espécies de Peixes-rei Odontesthes. Evermann& Kendall (Osteichthyes, Atherinopsidae) do sistema lagunar Patos-Mirim, no extremo sul do Brasil. Rev. Bras. de Zool., 22(2): 293-305. [ Links ]
3. Evermann, B.W. & W.C. Kendall. 1906. Notes on a collection of fishes from Argentina, South America, with descriptions of three new species. Proc. U.S. Nat. Mus. 31: 67-108. [ Links ]
4. Bocchino, A.R. 1971. Algunos peces fósiles del denominado Patagoniano del oeste de Chubut, Argentina. Ameghiniana 7 (1): 52-62. [ Links ]
5. Bogan, S., M.M. Cenizo, L.M. de los Reyes & N.E. González. 2006. Primera ictiofauna continental para el Pleistoceno Medio, Centinela del Mar, provincia de Buenos Aires, Argentina. 9° Congreso Argentino de Paleontología y Bioestratigrafía, Academia Nacional de Ciencias, Córdoba. Resúmenes: p.95. [ Links ]
6. Bogan, S. & M.M. Cenizo. 2008. Sobre la presencia de un Sciaenidae (Teleostei: Perciformes) en el Belgranense (Fm. Pascua, Pleistoceno superior) de Centinela del Mar, Buenos Aires (Argentina). Stvdia Geol. Salamanticensia (España) 44 (2) 213-220. [ Links ]
7. Bogan, S.; M.L. de los Reyes & M.M. Cenizo. 2009. Primer registro del género Jenynsia Günther, 1866 (Teleostei: Cyprinodontiformes) en el Pleistoceno medio tardío de la provincia de Buenos Aires (Argentina). Papéis Avulsos de Zoologia (Brasil) vol. 49 (5): 81-86. [ Links ]
8. Cenizo, M.M. & M.B. Ibañez. 2006. Primer registro para el género Oxyura Bonaparte, 1928 (Anatidae, Anseriformes) en el Pleistoceno Medio-Tardío de la provincia Buenos Aires. Ameghiniana 43 (4): 29R. [ Links ]
9. Cione, A.L. & E.P. Tonni. 1999. Biostratigraphy and chronological scale of upper-most Cenozoic in the Pampean Area, Argentina. En: E.P. Tonni & A.L. Cione (eds.), Quaternary Vertebrate Paleontology in South America. Quaternary South American and Antarctic Peninsula 12: 23-51. [ Links ]
10. Cione, A.L. & E.P. Tonni. 2005. Bioestratigrafía basada en mamíferos del Cenozoico superior de la región pampeana. En: R. de Barrio, R.O. Etcheverry, M.F. Caballé & E. Llambías (eds.), Geología y Recursos Minerales de la Provincia de Buenos Aires XVI Congreso Geológico Argentino (La Plata), Relatorio 11: 183-200. [ Links ]
11. De Buen, F. 1953. Los pejerreyes (familia Atherinidae) en la fauna Uruguaya, con descripción de nuevas especies. Universidade Sao Paulo, Bol. Inst. Ocea. 4 (12): 3-80. [ Links ]
12. De la Hoz, E. & E. Tosti-Croce. 1981. Osteología de Basilichthys australis Eigenmann (Teleostei - Atherinidae). An. Mus. Hist. Nat. Valp. 14: 229-245. [ Links ]
13. De los Reyes, L.M, M.M. Cenizo, F. Agnolin, S. Lucero, S. Bogan, R. Lucero, U.F.J. Pardiñas, F. Prevosti, F.& A. Scanferla. 2006. Aspectos paleofaunísticos y estratigráficos preliminares de las secuencias pliopleistocénicas de la localidad Centinela del Mar, provincia de Buenos Aires, Argentina. 9° Congreso Argentino de Paleontología y Bioestratigrafía, Academia Nacional de Ciencias. Resúmenes: p. 105. [ Links ]
14. Dyer, B.S. 1997. Phylogenetic revision of Atherinopsinae (Teleostei, Atheriniformes, Atherinopsidae), with comments on the systematics of the South American freshwater fish genus Basilichthys Girard. Museum Zoology, University of Michigan, Misc. Publ. 185: 1-64. [ Links ]
15. Dyer, B.S. 1998. Phylogenetic systematics and historical biogeography of the Neotropical silverside family Atherinopsidae (Teleostei, Atheriniformes). En: L.R. Malabarba, R.E. Reis, R.P. Vari, Z.M. Lucena & C.A.S. Lucena (eds), Phylogeny and Classification of Neotropical Fishes, pp. 519-536, Porto Alegre. [ Links ]
16. Dyer, B.S. 2000. Revisión sistemática de los pejerreyes de Chile (Teleostei, Atheriniformes). Estud. Oceanol. 19: 99-127. [ Links ]
17. Dyer, B.S. 2003. Family Atherinopsidae (Neotropical Silversides). En: R.E. Reis, S.O. [ Links ]
18. Fowler, H.W. 1903. Descriptions of new, little known and typical Atherinidae. Proc. Acad. Nat. Sci., Philadelphia 55: 727-742. [ Links ]
19. Girard, C. 1855. Abstract of a report to Lieut. Jas. M. Gilliss, U.S.N., upon the fishes collected during the U.S.N. Astronomical Expedition to Chili. Proc. Acad. Nat. Sci., Philadelphia 7(6): 197-199. [ Links ]
20. Kullander, S.O. & C.J. Ferraris (eds.), Check list of the Freshwater Fishes of South and Central America, pp. 515-525. Brasil. [ Links ]
21. Kullander, S.O. & C.J. Ferraris (eds.) 2006. Systematic revision of the South American silversides (Teleostei, Atheriniformes). Biocell 30 (1): 69-88. [ Links ]
22. Heil, C., J. King, M.A. Zárate & P.H. Schultz. (2002). Paleomagnetic and environmental magnetic studies of Pampeano Loess Deposits from Centinela del Mar, Argentina. EOS Trans. AGU Suppl. 83 GP71A-0974 [ Links ]
23. Isla, F.I., N.W. Rutter, J.E. Schanck & M.A. Zárate. 2000. La transgresión belgranense en Buenos Aires. Una revisión a cien años de su definición. Rev. Cuat. y Cienc. Amb. 1: 3-14. [ Links ]
24. Marrero, A. 1950. Flechas de Plata, atherínidos argentinos, pejerreyes y laterinos. Buenos Aires, 157 pp. [ Links ]
25. Lahille, F. 1929. El pejerrey. Bol. Min. Agr. Nac., Buenos Aires 28 (3): 261-395 [ Links ]
26. Pianta de Risso, E.N. & F.J.J. Risso. 1953. El "Cornalito" Sorgentinia incisa (Jenyns) n. g. y su ubicación sistemática (Atherinidae, Sorgentininae nueva subfamilia). Trab. Mus. Tres Arroyos, 1(1): 5-25. [ Links ]
27. Pozzi, A.J. & L.F. Bordalé. 1935. Cuadro sistemático de los peces marinos de la República Argentina. An. Soc. Cient. Arg. 120: 145-189. [ Links ]
28. Reading, H.G. & B.K. Levell. 1996. Control on the sedimentary rock record. En: H.G. Reading (ed.), Sedimentary Environments: Processes, Facies and Stratigraphy (3º edición). Blackwell Science, pp. 5- 35. [ Links ]
29. Ringuelet, R.A. 1975. Zoogeografía y ecología de los peces de aguas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América del Sur. Ecosur 2(3): 1-122. [ Links ]
30. Ringuelet, R.A., A.A. de Arámburu & R.H. Arámburu. 1967. Los peces argentinos de agua dulce. Comisión de Investigaciones Científicas de la Provincia de Buenos Aires, La Plata, 602 pp. [ Links ]
31. Rubilar, A. 1994. Diversidad ictiológica en depósitos continentales miocenos de la Formación Cura- Mallín, Chile (37-39° S): implicancias paleográficas. Rev. Geol. Chile 21 (1): 3-29. [ Links ]
32. Schultz, L.P. 1948. A revision of six subfamilies of atherine fishes, with descriptions of new genera and species. Proc. U.S. Nat. Mus. 98 (3220): 1-48. [ Links ]
33. Schultz, P.H., M. Zárate, R. Hames, B. C. Koeberl, T. Bunch, D. Storzer, P.R. Renne & J. Wittke. 2004. The Quaternary impact record from the Pampas, Argentina. Earth and Planetary Science Letters 219: 221-238. [ Links ]
34. Soibelzon, E., F.J. Prevosti, J.C. Bidegain, Y., Rico, D.H. Verzi & E.P. Tonni. (en prensa). Correlation of Cenozoic sequences of southeastern Buenos Aires province: biostratigraphy and magnetostratigraphy. Quat. Int. [ Links ]
35. White, B.N. 1985. Evolutionary relationships of the Atherinopsinae (Pisces: Atherinidae). Nat. Hist. Mus. Los Angeles County, Contrib. Sci. 368:1-20. [ Links ]
Recibido: 9-II-2008
Aceptado: 4-XI-2009

