










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
versión On-line ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.11 no.2 Ciudad Autónoma de Buenos Aires dic. 2009
ZOOLOGÍA
Estimación de la masa corporal de las especies de osos fósiles y actuales (Ursidae, Tremarctinae) de América del Sur
Leopoldo Héctor Soibelzon1 & Viviana Beatriz Tarantini2
1CONICET, Departamento Científico Paleontología de Vertebrados, Museo de La Plata, Paseo del Bosque, 1900 La Plata, Buenos Aires, Argentina.
2Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, 122 y 60, 1900 La Plata, Buenos Aires, Argentina.
Resumen: Contar con estimaciones precisas de la masa corporal de los taxones fósiles es esencial para realizar reconstrucciones paleobiológicas. Empleando relaciones demostradas entre la masa corporal y medidas anatómicas en mamíferos modernos algunos autores han propuesto ecuaciones predictivas a fin de estimar la masa de los taxones fósiles. En este trabajo calculamos la masa corporal de los osos vivientes y fósiles de América del Sur (Tremarctos ornatus y Arctotherium respectivamente) aplicando 61 ecuaciones alométricas seleccionadas a 19 medidas craneanas, dentarias y postcranenas. Adicionalmente estimamos el rango de masa de las cinco especies de Arctotherium por simple comparación de tamaño con el tamaño y peso de los osos actuales. Luego, por extrapolación a base de similitud morfológica con T. ornatus estimamos la masa de Arctotherium a fin de saber si nuestras estimaciones obtenidas con las ecuaciones alométricas eran posibles. También pusimos a prueba las ecuaciones empleadas utilizando un espécimen de T. ornatus de masa conocida. Nuestros resultados indican que las mejores ecuaciones para predecir la masa de Arctotherium son las basadas en medidas del fémur (8 ecuaciones) y craneanas (una ecuación). Por otra parte, las mejores ecuaciones para predecir la masa de T. ornatus son aquellas basadas en medidas del cráneo (una ecuación), fémur (dos ecuaciones) y húmero (tres ecuaciones).
Palabras clave: Masa corporal; Arctotherium; Tremarctos ornatus; América del Sur; Pleistoceno.
Abstract: Body mass estimation of extinct and extant South American bears (Ursidae, Tremarctinae). Precise estimates of body mass in fossil taxa are elementary to paleobiological reconstruction. Using demonstrated relationships between body mass and anatomical measurements in modern mammals, several authors proposed prediction equations to estimate the body mass of fossil taxa. We calculate the body mass of extant and fossil South American bears (Tremarctos ornatus and Arctotherium respectively) applying 61 selected allometric equations to 19 cranial, dental and postcranial measurements. We also estimate the body mass ranges of the five Arctotherium species by simple size comparison to size and weight of extant bears. Extrapolating on the basis of geometric similitude with T. ornatus we further estimated the body mass of Arctotherium in order to know if our estimations, bias published equations, were feasible. Also we test the prediction equations results with a specimen of Tremarctos ornatus of known body mass. We found that the best prediction equations for all Arctotherium species are based on femoral (8 equations) and cranial (one equation) measurements. In turn the best prediction equations for T. ornatus are based on cranial (one equation), first lower molar (one equation), femoral (two equations) and humeral measurements (three equations).
Key words: Body mass; Arctotherium; Tremarctos ornatus; South America; Pleistocene.
INTRODUCCIÓN
El objetivo de este trabajo es estimar la masa de las especies de Arctotherium y la de Tremarctos ornatus (Ursidae, Tremarctinae) utilizando ecuaciones de regresión tomadas de la literatura (e.g. Van Valkenburg, 1990, Christiansen, 1999, Egi, 2001), aplicadas a medidas relevadas sobre diferentes estructuras anatómicas (e.g. cráneo, m1 y elementos del estilo y zeugopodio), discutir los resultados obtenidos y la utilidad relativa de las ecuaciones descriptas. A tal fin se han recopilado 61 ecuaciones (que utilizan 19 medidas diferentes) y estas se han aplicado a 72 ejemplares de Arctotherium y Tremarctos ornatus.
La estimación de la masa de las especies fósiles es fundamental para la realización de inferencias paleobiológicas (e.g. tasa de natalidad, duración del período gestacional, de la lactancia y de los cuidados parentales) y paleoecológicas (e.g. posición del taxón en las redes tróficas, densidad poblacional). Por su parte, también es importante poder realizar estimaciones de la masa de individuos de especies actuales a partir de restos del esqueleto hallados en el campo; sobre todo en especies como T. ornatus donde la mayoría de los ejemplares en colección carecen de datos de masa y las posibilidades de captura a fin del relevamiento de este tipo de datos es extremadamente baja.
Los osos fósiles que habitaron América del Sur pertenecen a un grupo diverso de carnívoros pequeños a gigantescos que habitan América desde el Mioceno (Soibelzon et al., 2005) y tienen un único representante actual, el oso de anteojos (T. ornatus). En el registro fósil de América del Sur se encuentran representados por el género Arctotherium y sus cinco especies: A. angustidens, A. vetustum, A. wingei, A. tarijense y A. bonariense (Soibelzon, 2002, 2004). Estas, en conjunto con las especies de Arctodus (A. simus y A. pristinus) de América del Norte, conforman el clado de los osos de rostro corto (Soibelzon, 2002) o short-faced bears.
El registro fósil de Arctotherium comienza en el Ensenadense y finaliza en el Lujanense comprendiendo el lapso Pleistoceno Inferior a Holoceno Inferior; el grupo se distribuye desde el extremo norte de América del Sur hasta el extremo sur (Venezuela, Bolivia, Brasil, Uruguay, Argentina y sur de Chile; Soibelzon et al., 2005, Soibelzon & Rincón, 2007). En el Blanquense (Plioceno Superior) de El Salvador se comunicó la presencia de cf. Arctotherium (Soibelzon et al., 2008).
Las especies fósiles Arctotherium angustidens, Arctodus simus de América del Norte y Ursus spelaeus (Ursidae, Ursinae) de Europa, todas registrados durante el Pleistoceno, se encuentran entre los mamíferos continentales del Orden Carnivora de mayor masa corporal (Christiansen, 1999). Entre los mamíferos carnívoros sólo algunos creodontes, el mesoníquido gigante del Eoceno Andrewsarcus, el hyaenodonte Megistotherium y probablemente Panthera atrox pudieron haber igualado en tamaño a los más grandes úrsidos (Christiansen, 1999).
La masa de los osos actuales varía entre 27 y 800 kg (Nowak, 1999), encontrándose en un extremo Ursus malayanus (27 a 67 kg) y en el otro U. maritimus (150 a 800 kg). Según Nowak (1999) el rango de masas de las otras especies actuales es: T. ornatus (60 a 175 kg); Ursus thibetanus (65 a 110 kg), U. americanus (92 a 270 kg), U. arctos (95 a 324 kg), U. ursinus (55 a 145 kg). El registro del individuo más robusto, entre las especies actuales, corresponde a un macho de U. maritimus, cazado en 1960 en el noroeste de Alaska, cuya masa fue de 1002 kg (Wood, 1981). Cuando se trata de tamaño o masa corporal es necesario tener en cuenta que el dimorfismo sexual en tamaño en los Ursidae alcanza hasta el 20% (Nowak, 1999).
Existen estimaciones previas de la masa de algunos osos de rostro corto. La masa de Arctodus simus fue estimada en 590-630 kg para los individuos de mayor tamaño (Kurtén, 1967); en 700 a 800 kg para los machos, con individuos de hasta o algo más de 1000 kg (Christiansen, 1999) y la de Arctodus sp. (=Arctotherium sp.) en 500 kg (Fariña, 1996) y 308 kg (Fariña et al., 1998). Con respecto a la estimación realizada por Fariña et al. (1998) es importante aclarar que si bien los autores expresan que su estimación se basa en medidas cráneo-dentales tomadas sobre el individuo MACN 9645, este corresponde a varios restos del miembro posterior, sin elementos dentarios y/o craneales asociados.
Más recientemente, Soibelzon (2002) en su trabajo de Tesis Doctoral realizó estimaciones preliminares de la masa de las especies de Arctotherium aplicando las ecuaciones descriptas por Van Valkenburgh (1990) y Anyonge (1993) y discutió los resultados obtenidos por él y otros autores. Los resultados obtenidos por Soibelzon (2002) se presentan aquí ampliados y enriquecidos por el uso de mayor cantidad de ecuaciones alométricas y por la mayor cantidad de restos fósiles hallados y estudiados durante los años transcurridos.
Dentro del género Arctotherium los extremos de tamaño se encuentran en las especies A. angustidens y A. wingei. La primera con individuos de tamaño gigantesco, más grandes que los mayores individuos de Ursus spelaeus y de tamaños comparables a los más grandes individuos de Arctodus simus, y la segunda (A. wingei) con los individuos más pequeños, con tamaños comparables a los observados en U. americanus (92 a 270 kg; Nowak, 1999) y Tremarctos floridanus (Soibelzon, obs. pers.). De esta manera se pueden considerar tres categorías de tamaño a fin de agrupar a las especies de Artctotherium: tamaño pequeño a mediano (A. wingei y A. vetustum), mediano a grande (A. tarijense y A. bonariense) y grande a gigante (A. angustidens).
La estimación de la masa de los taxones fósiles se basa en métodos indirectos, uno de ellos es la aplicación de ecuaciones de regresión. Las ecuaciones se obtienen a partir del estudio de la relación entre la masa de individuos actuales y algunos de sus caracteres cuantitativos lineales (e.g. longitud o ancho del húmero, tibia o molares) o de área (e.g. área de superficies articulares de huesos largos) y pueden ser aplicadas a la estimación de la masa de los individuos fósiles con ciertas restricciones.
Una de las principales restricciones de este tipo de métodos es la interpolación más allá del rango original de los datos, lo cual puede generar valores con una incertidumbre asociada muy alta, ya que no hay pruebas de que estos modelos lineales se continúen más allá del rango original de los datos utilizados para su generación (Egi, 2002). Esta aclaración es necesaria porque muchos de los ejemplares de Arctotherium (especialmente dentro de A. angustidens) presentan tamaños que escapan al rango de datos con que las ecuaciones se construyeron originalmente.
Otra restricción está relacionada con las proporciones de los elementos anatómicos del o los taxones utilizados para construir las ecuaciones y las de los taxones fósiles objeto de las estimaciones, más allá de su relación filogenética (véase Gingerich, (1990). En este sentido, también debe tenerse en cuenta que existen algunas diferencias en algunas de las proporciones del cráneo y los molares entre Arctotherium y las especies de Ursus (sobre las que se construyen las ecuaciones para Ursidae). Por esta razón, algunos valores obtenidos para individuos gigantes con algunas ecuaciones resultan irreales y no deben ser considerados como posibles.
Abreviaturas institucionales. CEHA: Colección del Centro de Estudios del Hombre Austral, Chile. GALY: Colección del Grupo de Arqueología del Liceo de Young, República Oriental del Uruguay. GP: Instituto de Geociencias, Universidade de Sao Paulo, Brasil. IGC: Instituto de Geociencias, Universidade Federal de Minas Gerais, Brasil. MACN: Colección de Paleontología de Vertebrados, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Argentina. MACN-zool: Colección de Mastozoología, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Argentina. MARC: Museo y Archivo Regional Castelli, Argentina. MBLUZ: Museo de Biología de la Universidad del Zulia, Venezuela. MHJ: Museo Histórico de Junín, Argentina. MHNLP: Museo de Historia Natural de La Paz, Bolivia. MLP: Colección Paleontología de Vertebrados, Museo de La Plata, Argentina. MLP-M: Colección de mastozoología, Museo de La Plata, Argentina. MMMP: Museo Municipal de Mar de Plata "Lorenzo Scaglia", Argentina. MMPH: Museo Municipal "Punta Hermengo", Argentina. MNHNP-PAM: Museum National de Histoire Naturelle, Colección del Pampeano; Francia. MPD: Museo Paleontológico de Daireux, Argentina. MPS: Museo Paleontológico de San Pedro, Argentina. MPV: Museo Paleontológico de Valencia, España. NHM: Natural History Museum, London, Inglaterra. PIMUZ: Paläontologisches Institut und Museum der Universität Zürich, Suiza. UZM: Zoological Museo University of Copenhagen, Dinamarca. VF: Museo Royo y Gómez, Universidad Central de Venezuela, Venezuela.
Materiales examinados
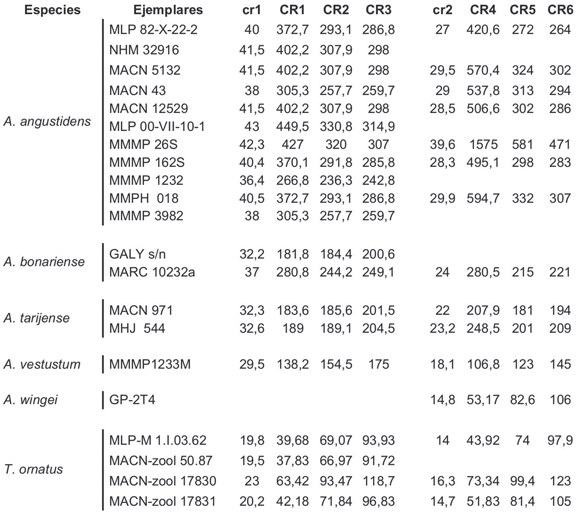
A. angustidens: NHM 32916; NHM 32971; NHM 32972; MACN 43; MACN 851; MACN 12529; MACN 17986; MACN 5132; MACN 2275; MACN 16260; MACN 9609; MARC 10232 a 2/242; MHNLP s/n; MLP 10-81; MLP 10-15; MLP 10- 34; MLP 10-21; MLP 09-I5-1; MLP 97-II-2-1; MLP 00-VII-10-1; MLP 00-VII-15-1; MLP 35-IX- 26-4; MLP 82-X-22-2; MLP 99-X-5-1; MLP 80-XII- 1-1; MMPH 018; MMMP 26S, MMMP 162S, MMMP 1232; MMMP 3981; MMMP 3982; MMMP 3983; MNHNP-PAM 179; MPS 60; MPV 614. A. bonariense: CALY s/n; MACN 18599; MACN 2668; MACN 13971; MACN 9645; MACN 9646; MACN 9647; MARC 10232a2/242; MMMP1441, PIMUZ 4215. A. tarijense: CEHA.; MACN 1458; MACN 2667; MACN 8581; MACN 14614; MACN 971; MHJ 544; MLP 83-I-15-6; MMMP 311S; MPD s/n. A. vetustum: MACN 1201; MACN 17486; MMMP 1233M. A. wingei: UZM 5721-22; UZM 5249; MACN 1453; MBLUZP-5.119; MBLUZP-5.119; MBLUZP- 5.121; GP-2T4; ICG-10. Arctotherium sp.: MACN 12558; MACN 11443; MMMP 311S.
Para más datos sobre los ejemplares arriba citados, véanse Soibelzon (2002, 2004), Soibelzon et al. (2005) y Soibelzon y Rincón (2007).
Tremarctos ornatus: MACN-zool 50.87, MACN-17830, MACN-zool 17831, MLP-M 1.I.03.62.
Metodología
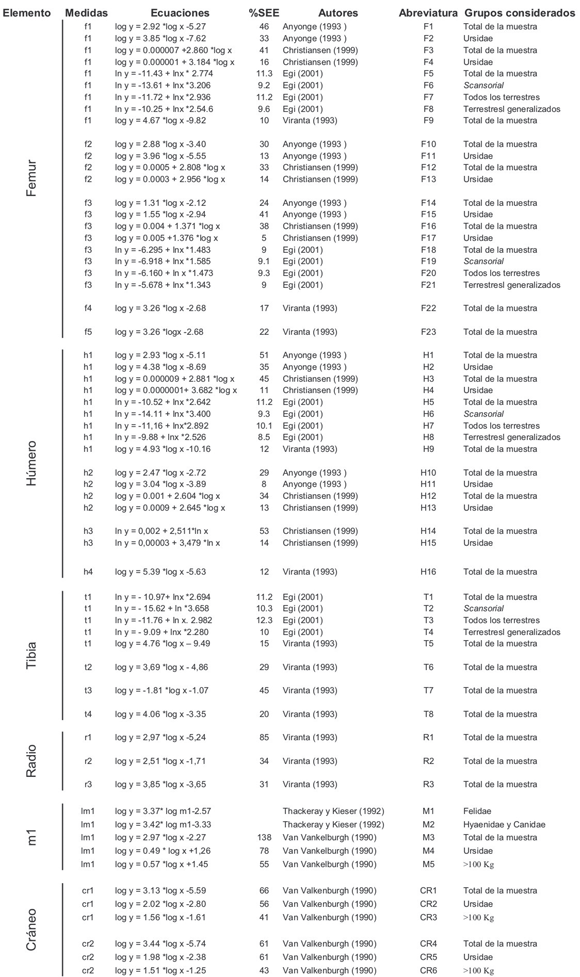
Abreviaturas. cr1 y cr2 medidas relevadas sobre el cráneo, f1a f5 medidas relevadas sobre el fémur; h1 a h4 medidas relevadas sobre el húmero; lm1 longitud del m1; r1 a r3 medidas relevadas sobre el radio; t1 a t4 medidas relevadas sobre la tibia. CR1 a CR8 ecuaciones obtenidas por diferentes autores sobre medidas del cráneo; F1 a F23 ecuaciones obtenidas por diferentes autores sobre medidas del fémur; H1 a H13 ecuaciones obtenidas por diferentes autores sobre medidas del húmero; M1 a M6 ecuaciones obtenidas por diferentes autores sobre medidas del primer molar inferior; R1 a R3 ecuaciones obtenidas sobre medidas de radio; T1 a T7 ecuaciones obtenidas por diferentes autores sobre medidas de la tibia (véase Materiales y Métodos). Error de predicción (% PE) y error estándar de la estimación (% SEE).
Las masas se estimaron a base de ecuaciones de regresión propuestas por diferentes autores (véase Tabla 1) que utilizan como variable independiente medidas tanto dentarias como craneanas y post-craneanas.
Tabla 1. Lista de las ecuaciones utilizadas en el presente estudio y de sus correspondientes abreviaturas, medidas de los elementos anatómicos utilizados al construir las ecuaciones (f1 a f5; h1 a h4; t1 a t4; r1 a r3 y lm1; véase materiales y Métodos) autores, grupos considerados en la construcción de cada ecuación y su porcentaje de error (% SEE). Falta el % PE, en muchos casos es mejor indicador que el % SEE.
Medidas consideradas. a) Lineales. Para el húmero, fémur, tibia y radio se tomaron la longitud total, la circunferencia y el ancho transversal de la diáfisis. Medidas del fémur: f1: longitud total; f2: circunferencia de la diáfisis tomada al 50% de su longitud; f4: diámetro máximo sobre los cóndilos; f5: ancho transversal tomada al 50% de su longitud. Medidas del húmero: h1: longitud total; h2: circunferencia de la diáfisis tomada al 50% de su longitud; h3: diámetro transverso máximo de la epífisis distal; h4: ancho transversal tomado al 50% de su longitud. Medidas del radio: r1: longitud total; r2: diámetro máximo sobre la superficie articular proximal; r3: diámetro máximo sobre la superficie articular distal. Medidas de la tibia: t1: longitud total; t2: diámetro transverso máximo de la superficie articular proximal; t3: diámetro transverso máximo de la superficie articular distal (véanse Anyonge, 1993; Viranta, 1994; Christiansen, 1999 y Egi, 2001). Sobre el cráneo se relevó la longitud cóndilo-basal y distancia entre el borde anterior de la órbita y los cóndilos occipitales (cr1 y cr2 respectivamente; (véase Van Valkenburg, 1990). Finalmente, se utilizó la longitud mesio-distal del m1 (lm1) (véanse Van Valkenburg, 1990 y Thackeray & Kieser, 1992). b) De área. El cálculo de áreas de la superficie articular distal de fémur y tibia (f3 y t4 respectivamente; véase Ruff, 1990).
Las medidas fueron tomadas con calibre Weyersberg 1/20 y/o Vernier 0-500 X 0.5 mm y se encuentran expresadas en mm.
Ecuaciones de regresión. Anyonge (1993) utiliza una muestra de 28 especies de carnívoros (Mamalia, Carnivora) a la que divide en cuatro grupos: total de la muestra (28 especies), cánidos (7 especies), úrsidos (4 especies) y félidos (14 especies) y obtiene diferentes ecuaciones de regresión para cada uno de los cuatro grupos utilizando como variables independientes medidas de fémur y húmero (Tabla 1 ecuaciones F1, F2, F10, F11, F14, F15, H1, H2, H10 y H11).
Egi (2001) utiliza una muestra de 47 especies agrupadas en 7 conjuntos según los hábitos locomotores: total de la muestra (47 especies), arborícolas (9 especies), buenos trepadores (scansorial, 14 especies), todos los terrestres (16 especies), terrestres generalizados (6 especies), terrestres semi fosoriales (6 especies) y cursoriales (8 especies) y terrestres semi acuéticos (4 especies), esta última, no utilizada en nuestro trabajo. El autor utiliza el modelo RMA (reduce major axis) en lugar del modelo de los mínimos cuadrados (de uso general), porque considera que subestima las masas cuando se aplica en especies con individuos de gran tamaño. Este modelo toma los valores transformados en logaritmos naturales (Tabla 1 ecuaciones F5 a F8, F18 a F21, H5 a H8, T1 a T4).
Christiansen (1999) utilizó una muestra de 30 especies de carnívoros (incluídos Canidae, Hyaenidae, Mustelidae, Procyonidae y Viverridae) para estimar la masa de Arctodus simus y Ursus spelaeus (Ursidae) a partir de medidas del fémur y húmero. Aplica las ecuaciones a tres grupos: el total de la muestra (n=30), Ursidae (n=4) y Felidae (n=9). (Tabla 1 ecuaciones F3, F4, F16, F17, H3, H4, H12 a H15).
Thackeray y Kieser (1992) utilizaron la longitud mesio-distal del m1 para construir dos ecuaciones de regresión, una para felidos (26 individuos) y otra para hiénidos y cánidos (26 individuos) (Tabla 1 ecuaciones M1 y M2).
Van Valkenburgh (1990) recurrió a medidas del cráneo y el m1 de 72 especies de carnívoros y obtiene ecuaciones de regresión para el total de la muestra (incluye grupo de Dasyuridae, n=2; Procyonidae, n=5; Viverridae, n=4), para Cánidae (n=14), Ursidae (n=8), Mustelidae (n=19) y Felidae (n= 16). También presenta ecuaciones aplicables a cuatro grupos de tamaño: menores a 6 kg., 6 a 10 kg, 10 a 100 kg. y mayores que 100 kg (Tabla 1 ecuaciones M3 a M5 y CR1 a CR6).
Viranta (1994) construyó ecuaciones de regresión a fin de estimar la masa de Ursus spelaeus sobre medidas del húmero, fémur y tibia de Ursus arctos (n= 14), U. maritimus (n=14) y U. americanus (n=3) (Tabla 1 ecuaciones F9, F22, F23, H9, H16, T5 a T8, R1 a R3).
Todas las ecuaciones fueron aplicadas también a cuatro ejemplares de oso de anteojos (Tremarctos ornatus) el único Tremarctinae viviente y por lo tanto el úrsido más cercano filogenéticamente a Arctotherium. Asimismo T. ornatus comparte varias características anatómicas con Arctotherium, algunas de los cuales podrían incidir sobre las estimaciones (e.g. acortamiento del rostro). La masa de los individuos adultos varía entre 60 y 175 kg (Nowak, 1999). Lamentablemente el rango de masas de T. ornatus solo se superpone parcialmente con el de las especie pequeñas y medianas de Arctotherium (A. wingei y A. vetustum y en menor medida A. tarijense) siendo las demás especies del género fósil de mayor tamaño que el de la especie actual. Afortunadamente pudo estudiarse un ejemplar hembra adulto de la misma especie proveniente del Zoológico Municipal de La Plata (MLP-M 1.I.03.62) cuya masa fue establecida en 72 kg mediante una balanza por el personal de la institución cuando el animal se encontraba en buen estado sanitario (C. Coronel, com. pers.) y de esta manera contamos con un indicio sobre cuales ecuaciones estimaban con más exactitud la masa de la especie.
Adicionalmente y con el fin de contar con otro control de los resultados para las especies fósiles se estimó por comparación con el tamaño del esqueleto de otros osos y carnívoros de gran tamaño el rango posible de cada una de las cinco especies de Arctotherium. A tal fin se comparó la mayor cantidad de elementos esqueletarios disponibles y se estimó la mayor masa posible para cada especie, o sea el extremo superior del rango de masas (A. wingei 150 kg; A. vetustum 300 kg; A. tarijense 400 kg; A. bonariense 500 kg; A. angustidens 1200 kg.) Posteriormente se calculó para las especies actuales cual era la diferencia (Dm) entre el valor mínimo (Mmi) y el máximo (Mma) del rango y que porcentaje (X) de la Mma representaba este.
En el caso de T. ornatus:
Mma (175 kg) - Mmi (60 kg)= Dm (115 kg)
Mma (175 kg) 100%
Dm (115 kg) X= 65,7%
Los resultados obtenidos fluctuaron entre el 60% (U. malayanus) y el 80% (U. maritimus) y 65,7% para T. ornatus.
Se utilizó el porcentaje obtenido para T. ornatus a fin de calcular el extremo inferior del rango de masas (Mmi) de las especies de Arctotherium debido a la cercanía filogenética entre los taxones y a que este valor es muy cercano a la media de los valores obtenidos para todas las especies actuales (67,2 %):
En el caso de A. angustidens:
100% Mma (1200 kg)
65,7% Dm (788 kg)
Finalmente: Mmi = Mma (1200 kg) - Dm (788 kg) = 412 kg
De esta manera, se consideran como rangos de masa posibles los siguientes: A. wingei 51-150 kg y A. vetustum 102-300 kg (tamaño pequeño a mediano); A. tarijense 135-400 kg y A. bonariense 171-500 kg (tamaño mediano a grande) y A. angustidens 412-1200 kg (tamaño grande a gigante).
Finalmente y habida cuenta de la similitud morfológica entre T. ornatus y Arctotherium, las masas obtenidas para las especies fósiles (a través de las ecuaciones alométricas) se testearon utilizando las siguientes relaciones descriptas por la morfología geométrica:
MX / MTo = (LxX / LxTo)3
De donde MX = (LxX / LxTo)3 * MTo
MX= masa de la especie problema. MTo= masa de T. ornatus. LxX= medida x en la especie problema. LxTo= medida x en T. ornatus. Estas relaciones se aplicaron a medidas del húmero, fémur, cráneo y m1 y los resultados de encuentran en la Tabla 2. Esta ecuación indica la masa potencial de un individuo problema (en este caso un fósil) por comparación geométrica con un individuo de morfología semejante y masa conocida (en este caso T. ornatus).
Tabla 2. Masas (kg) de Arctotherium obtenidas por relación geométrica con Tremarctos ornatus y a partir de medidas relevadas sobre el m1 (lm1); fémur (f1, f2, f4 y f5); cráneo (cr1 y cr2) y húmero (h1, h2 y h3) expresadas en cm (veáse Materiales y Métodos).

DISCUSIÓN y RESULTADOS
Los valores obtenidos a base de la similitud geométrica con T. ornatus (a partir de las medidas cr1, cr2, f4, f5 y h1) sustentan los rangos de masa potenciales para cada especie propuestos por simple comparación de tamaños con especies actuales (véase Materiales y Métodos) y por lo tanto estos rangos se emplearán en la discusión de los resultados.
Las masas obtenidas a partir de las ecuaciones alométricas utilizadas se muestran en las Tablas 3 a 8. Los resultados obtenidos corroboran que la utilidad de las ecuaciones como herramienta predictiva varía según el rango de tamaños de los ejemplares problema y de las especies utilizadas al construir las ecuaciones. Para las especies de Arctotherium las estimaciones más cercanas a lo que nosotros planteamos como posibles, fueron obtenidas para las especies pequeñas y medianas: A. wingei, A. vetustum y A. tarijense. Esto no es sorprendente pues la masa de los individuos de estas especies se encuentra dentro del rango de masas de las especies actuales utilizadas para construir las ecuaciones alométricas.
De las 5 ecuaciones (M1 a M5) que utilizan la longitud del m1 (lm1) como variable independiente la única que estimó correctamente la masa del ejemplar MLP-M 1.I.03.62 de T. ornatus fue M4 (propuesta para Ursidae, véanse Tablas 1 y 3). Las ecuaciones M3 (a base de toda la muestra considerada) y M4 predijeron la masa de la especie más pequeña (A. wingei), mientras que las ecuaciones M3 y M5 (desarrollada para especies de más de 100 kg) resultaron mejores para las especies medianas (A. vetustum, A. tarijense y A. bonariense); finalmente la masa de la especie de mayor tamaño (A. angustidens) fue subestimada por todas las ecuaciones utilizadas, obteniéndose valores más cercanos a lo que consideramos posible con la ecuación M1 (Tabla 3). Por lo tanto, ninguna de las ecuaciones que utilizan como variable independiente la longitud del m1 (lm1) debería ser aplicada a todas las especies de Arctotherium en conjunto.
Tabla 3. Masas (kg) de todas las especies de Arctotherium y Tremarctos ornatus estimadas a partir de la longitud del m1 (lm1) expresada en cm (véase Materiales y Métodos) y aplicando las ecuaciones M1 a M5 (véase Tabla 1).

Las estimaciones obtenidas a base de medidas craneales (Tabla 4) muestran un patrón similar al de otras estimaciones pues la masa de las especies pequeñas y medianas (A. wingei, A. vetustum y A. tarijense) es bien estimada por todas las ecuaciones, mientras que todas excepto una (CR4) subestiman considerablemente la masa de A. bonariense y sobre todo la de A. angustidens. La masa del ejemplar MLP-M 1.I.03.62 de T. ornatus fue correctamente estimada por CR5 (construida a base de especies de Ursidae) y en menor medida por la ecuación CR2 (fundada también en úrsidos).
Tabla 4. Masas (kg) de todas las especies de Arctotherium y Tremarctos ornatus estimadas aplicando las ecuaciones CR1 a CR6 (véase Tabla 1) y a partir de medidas del cráneo (cr1 y cr2) expresadas en cm (véase Materiales y Métodos).
Aunque las medidas craneales y dentarias son las más utilizadas en la literatura, por su mayor frecuencia en el registro fósil, las ecuaciones sobre medidas del estilopodio y zeugopodio son mejores estimadoras ya que su estructura se ve afectada por el stress derivado de soportar el peso del cuerpo (Legendre & Roth, 1988). Adicionalmente las medidas del fémur y húmero han sido postuladas como las más adecuadas para estimar la masa de los osos fósiles (véanse Anyonge, 1993; Viranta, 1994; Christiansen, 1999, Egi, 2002).
Nuestros resultados indican que las ecuaciones basadas en medidas de fémur (Tabla 5) y húmero (Tabla 6) estiman la masa de los individuos de todas las especies de Arctotherium de manera realista, a excepción de algunos ejemplares gigantescos (e.g. MACN 5132). Las ecuaciones F6, F7, F9 (basadas en la longitud del fémur) y F10 y F13 (basadas en la circunferencia del fémur) presentan bajos %PE y menor %SEE y han estimado correctamente la masa tanto de Arctotherium como de T. ornatus. Las ecuaciones F17 a F19 tienen muy bajos %SEE y dan resultados posibles para las especies fósiles pero no para T. ornatus. La ecuación F23 parece estimar correctamente la masa de A. angustidens y llamativamente los resultados no se ven afectados por el enorme tamaño de algunos individuos. Por otra parte la ecuación F22 ha sobreestimado en gran medida la masa de de todas las especies consideradas.
Tabla 5. Masas (kg) de las especies de Arctotherium angustidens, A. bonariense, A. tarijense, Arctotherium sp. y Tremarctos ornatus estimadas a partir de medidas relevadas sobre el fémur (f1, f2, f3, f4 y f5) expresadas en cm o cm2 según corresponda (véase Materiales y Métodos) y aplicando las ecuaciones F1 a F23 (véase Tabla 1).
Tabla 6. Masas (kg) de las especies de Arctotherium angustidens, Arctotherium sp. y Tremarctos ornatus estimadas aplicando las ecuaciones H1 a H16 (véase Tabla 1) y a partir de medidas del húmero (h1, h2, h3 y h4) expresadas en cm (véase Materiales y Métodos).
En cuanto a las ecuaciones que utilizan medidas del húmero, fueron aplicadas solo a ejemplares de A. angustidens pues no se conocen húmeros asignables a otras especies. Los valores más realistas fueron obtenidos utilizando las ecuaciones H1, H4, H7, H10, H12 y H14 (Tabla 6), es destacable que cada una de estas seis ecuaciones produce valores muy cercanos para el mismo individuo y dentro de la especie. Por su parte, las ecuaciones H1, H6, H10, H12 y H13 producen valores cercanos a la masa conocida del ejemplar MLP-M 1.I.03.62 de T. ornatus.
Las ecuaciones basadas en medidas de la tibia subestimaron tanto la masa de T. ornatus como la de Arctotherium. La ecuación T8 en particular sobreestimó la masa de Arctotherium y subestimó la de T. ornatus (Tabla 7).
Tabla 7. Masas (kg) de las especies de Arctotherium angustidens, A. bonariense, Arctotherium sp. y Tremarctos ornatus estimadas aplicando las ecuaciones T1 a T8 (véase Tabla 1) y a partir de medidas relevadas sobre la tibia (t1, t2, t3 y t4) expresadas en cm o cm2 según corresponda (véase Materiales y Métodos).

De las tres ecuaciones basadas en medidas del radio, la ecuación R2 brindó resultados realistas para las especies analizadas, mientras que la R3 devolvió resultados imposibles. Por su parte la ecuación R1 subestimó la masa de todos los individuos analizados (Tabla 8).
Tabla 8. Masas (kg) de las especies de Arctotherium angustidens, A. bonariense, Arctotherium sp. y Tremarctos ornatus estimadas aplicando las ecuaciones R1 a R3 (véase tabla 1) y a partir de medidas relevadas sobre el radio (r1, r2 y r3) expresadas en cm (véase Materiales y Métodos).

CONCLUSIONES
Las ecuaciones que mejor han estimado la masa de todas las especies de Arctotherium fueron CR4, F6, F7, F10, F12 y F13 y F17 a F19. Si bien no ha podido ser estimada la masa de las dos especies fósiles más pequeñas (A. vetustum y A. wingei) a base de medidas del fémur, las ecuaciones F10 y F13 han estimado correctamente la masa de T. ornatus y por lo tanto es válido creer que pueden hacerlo con la de las dos especies fósiles pequeñas.
Las ecuaciones M1, F23, H1, H4, H7, H10, H12, H14 y R2 valoran correctamente la masa de los individuos de mayor tamaño, por lo que se recomienda su aplicación a la estimación de masas de A. angustidens.
La masa de los individuos de tamaño mediano a grande (A. tarijense y A. bonariense) fue correctamente estimada por la ecuación M5.
Por su parte, la ecuación M3 estima la masa de los individuos de tamaño mediano a pequeño de manera aceptable por lo tanto se recomienda su uso al estimar la masa de A. tarijense, A. bonariense, A. vetustum y A. wingei.
Adicionalmente las ecuaciones M4, CR5 y CR 6 estimaron correctamente la masa de los individuos de tamaño más pequeño (A. vetustum y A. wingei).
Finalmente, la masa del ejemplar MLP-M 1.I.03.62 de T. ornatus fue estimada con suficiente exactitud por CR5, M4, F9, F13, H6, H10, y H13 y en menor medida por CR2, F10, H1, H12, y R2.
AGRADECIMIENTOS
Los autores deseamos agradecer a los curadores de las colecciones estudiadas: A. Kramarz (MACN); Olga Vaccaro (MACN zool.); A. Currant (NHM, Londres); C. de Muizzon y F. Pujos (MNHN, Paris); D. Bo (MMPH); J. Petrocelli (MCA); J. Cabañez (Liceo Young); I. Olivares y D. Verzi (MLP M); A. Dondas (MMMP); K. Aaris (UZM), M. Reguero y L. Pomi (MLP). A Alejo Scarano, Gustavo Bulus, Sergio Vizcaíno, Susana Bargo y Per Christiansen por su asesoramiento sobre cuestiones metodológicas. A los técnicos del Departamento Paleontología de Vertebrados (MLP): Juan José Molly y Javier Posik por la preparación de algunos ejemplares. A los técnicos de la División Zoología de Vertebrados (MLP) Luis Pagano y Lalo Etcheverry. A Carlos Coronel (Zoológico Municipal de La Plata). Este trabajo fue parcialmente financiado por CONICET, ANPCyT PICT 38171 y UNLP.
BIBLIOGRAFÍA
1. Anyonge, W. 1993. Body mass in large extant and extinct carnivores. J. Zool., Lond., 231: 339-350. [ Links ]
2. Christiansen, P. 1999. What size were Arctodus simus and Ursus spelaeus (Carnivora: Ursidae ?. Ann. Zool. Fenn. 36: 93-102. [ Links ]
3. Egi, N. 2001. Body mass estimates in extinct mammals from limb bones dimension: the case of North American hyaenodontids. The Palaeontological Association. Palaeontology. 44: (3) 497-528. [ Links ]
4. Fariña, R.A. 1996. Trophic relationships among Lujanian Mammals. Evol. Theory 11: 125-134. [ Links ]
5. Fariña, R.A., S.F. Vizcaíno & M.S. Bargo. 1998. Body mass estimations in Lujanian (Late Pleistocene- Early Holocene of South America) mammal megafauna. Mast. Neot. 5 (2): 87-108. [ Links ]
6. Gingerich, P.D. 1990. Prediction of Body Mass in Mammalian Species from Long Bone Lengths and Diameters. Mus. Pal., The Univ. of Michigan. 28(4):79-92. [ Links ]
7. Kurtén, B. 1967. Pleistocene bears of North America: 2 Genus Arctodus, short faced bears. A. Zool. Fenn. 117: 1-60. [ Links ]
8. Legendre, S. & Roth, C. 1988. Correlation of carnassial tooth size and body weight in Recent carnivores (Mammalia). Historical Biology 1, 85-98. [ Links ]
9. Nowak, R.M. 1999. Walker's Mammals of the World, John Hopkins University Press, Baltimore, Maryland. 1936 pp. [ Links ]
10. Ruff, C. 1990. Body mass and hindlimb bone crosssectional and articular dimensions in anthropoid primates. En: Damuth, J. & MacFadden, B.J. (Eds.) Body Size in Mammalian Paleobiology: Estimation and Biological Implications. pp. 119-150. Cambridge University Press. New York. [ Links ]
11. Soibelzon, L.H. 2002. Los Ursidae (Carnivora, Fissipedia) fósiles de la Republica Argentina. Aspectos sistemáticos y paleoecológicos. PhD Thesis, Universidad Nacional de La Plata (inédito). [ Links ]
12. Soibelzon, L.H. 2004. Revisión sistemática de los Tremarctinae (Carnivora, Ursidae) fósiles de América del Sur. Rev. Mus. Arg. Cien. Nat. 6: 107-133. [ Links ]
13. Soibelzon, L.H. & A. Rincón. 2007. The fossil record of the short-faced bears (Ursidae, Tremarctinae) from Venezuela. Systematic, biogeographic, and paleoecological implications. N. J. Geol. Pal. 245: 287- 298. [ Links ]
14. Soibelzon, L.H., E.P. Tonni & M. Bond. 2005. The fossil record of South American short-faced bears (Ursidae, Tremarctinae). J. South Am. E. Sci. 20: 105-113. [ Links ]
15. Soibelzon, L.H., M.R. Romero, D.Huziel Aguilar & V.B Tartarini. 2008. A new finding in the Blancan of the El Salvador indicates the probable dating of the Tremarctinae's (Ursidae, Carnivora) entrance to South America. N. J. Geol. Pal. 250: 1-8. [ Links ]
16. Tackeray J.F. & J.A. Kieser. 1992. Body mass and carnassial length in modern and fossil carnivores. Trans. Mus. 35 :(24) 337-341. [ Links ]
17. Van Valkenburgh, B. 1990. Skeletal and dental predictors of body mass in carnivores. En: Damuth, J. & MacFadden, B.J. (Eds.): Body Size in Mammalian Paleobiology: Estimation and Biological Implications. pp. 181-206. Cambridge University Press. New York. [ Links ]
18. Viranta, S. 1994. Limb bone proportions and body mass of cave bear (Ursus spelaeus). Hist. Biol. (7): 293-250. [ Links ]
19. Wood, G.L. 1981. The Guinness Book of animal records. 3rd edit. Guinness publishing 240 pp. [ Links ]
Recibido: 5-III-2009
Aceptado: 23-XI-2009

