










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
versión On-line ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.12 no.2 Ciudad Autónoma de Buenos Aires dic. 2010
PALEONTOLOGIA
A new hadrosauroid (Dinosauria: Ornithopoda) from the Allen Formation (Late Cretaceous) of Patagonia, Argentina
Rubén D. Juárez Valieri1, José A. Haro2, Lucas E. Fiorelli3 & Jorge O. Calvo4
1 Centro Paleontológico Lago Barreales (CePaLB). Universidad Nacional del Comahue. Ruta Provincial N° 51, km 65. (8300). Neuquén, Argentina. rubendjuarez@gmail.com.
2Centro de Investigaciones Paleobiológicas (CIPAL). Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Velez Sarfield 299 (5000). Córdoba, Argentina. augustoharo@yahoo.com.
3Centro Regional de Investigaciones Científica y Transferencia Tecnológica (CRILAR). Entre Ríos y Mendoza. (5301). Anillaco, La Rioja, Argentina. lfiorelli@crilar-conicet.com.ar.
4Centro Paleontológico Lago Barreales (CePaLB). Universidad Nacional del Comahue. Ruta Provincial N° 51, km 65. (8300). Neuquén, Argentina. jorgecalvo@proyectodino.com.ar
Abstract: A new hadrosauroid taxon from the Late Cretaceous Allen Formation is erected, on the basis of abundant cranial and postcranial material from the Salitral Moreno locality of the Rio Negro Province, Argentina. The new taxon is here named as Willinakaqe salitralensis gen. et sp. nov., which is characterized by having an autapomorphous premaxilla with a long and convex rostrolateral surface rostral to the narial fossa and associated postcrania developing an unique character combination, including: dorsal vertebrae with a shallow fossa upon the base of the transverse processes; sacral and proximal caudal neural spines more than three times the height of the centrum; distal region of the postacetabular process of ilium ventrally deflected, among others. This new hadrosauroid closely resembles Secernosaurus koerneri, the other valid hadrosauroid species from South America, based in several synapomorphies, as: angle greater than 115° between the lateral margin of the facet for scapular articulation and the glenoid in the coracoid; supra-acetabular process of the ilium asymmetrical, caudodorsal margin of ilium with a well-defined ridge continuous with the dorsal margin of the proximal region of the postacetabular process. Moreover, previous records of Lambeosaurinae from the Late Cretaceous of South America are referred to W. salitralensis, and thus, the presence of lambeosaurines in this continent is here rejected. Accordingly, Willinakaqe salitralensis gen. et sp. nov. is currently the only hadrosauroid species reported from the Allen Formation.
Key words: Hadrosauroidea; Morphology; Palaeobiogeography; Late Cretaceous; South America.
Resumen: Se erige un nuevo taxón de hadrosauroideo del Cretácico Superior de la Formación Allen, en base a abundante material craneal y postcraneal de la localidad de Salitral Moreno, en la Provincia de Río Negro, Argentina. El nuevo taxón es denominado Willinakaqe salitralensis gen. et sp. nov., y se caracteriza por tener un premaxilar autapomórfico con una superficie rostrolateral alargada y convexa rostral a la fosa narial y postcráneos asociados con una combinación de caracteres únicos, que incluye: vértebras dorsales con una fosa poco profunda en la base de los procesos transversos, espinas neurales sacras y caudales proximales más de tres veces más altas que el centro; región distal del proceso postacetabular del ilion desviada ventralmente, entre otros. Este nuevo hadrosauroideo es muy similar a Secernosaurus koerneri, la otra especie válida de hadrosauroideo de América del Sur, con el que comparte varias sinapomorfías, como: ángulo mayor que 115 ° entre el borde lateral de la faceta articular escapular y la superficie glenoidea en el coracoides, proceso supra-acetabular del ilion asimétrico, margen caudodorsal del ilion con una cresta bien definida continua con el margen dorsal de la región proximal del proceso postacetabular. Los registros asignados anteriormente a Lambeosaurinae del Cretácico Tardío de América del Sur son referidos a Willinakaqe salitralensis gen. et sp. nov., y por lo tanto, se rechaza la presencia de lambeosaurinos en este continente. En consecuencia, Willinakaqe salitralensis gen. et sp. nov. es actualmente la única especie de hadrosauroideo reportada en la Formación Allen.
Palabras clave: Hadrosauroidea; Morfología; Paleobiogeografía; Cretácico Superior; América del Sur.
INTRODUCTION
Hadrosauroids are known from the Southern Hemisphere ever since Casamiquela (1964) described the first materials. The first named hadrosauroid from South America was Secernosaurus koerneri, which is based on an incomplete postcranial skeleton recovered close to the Río Chico locality in the Chubut province, central Patagonia, Argentina (Brett-Surman, 1979). Since its original description, Secernosaurus was considered as a problematic taxon because some bones present a high degree of deformation; consequently, several anatomical characters previously employed to assign the species as a basal hadrosauroid are product of taphonomic processes (Brett-Surman, 1979; Bonaparte et al., 1984; Bonaparte, 1996). Additionally, the precise stratigraphical provenance of Secernosaurus is unknown, decreasing its biostratigraphical and biogeographical relevance (Bonaparte, 1996; Prieto-Márquez & Salinas, 2010). The second species of Hadrosauroidea proposed and described for Cretaceous beds in South America is Kritosaurus australis, coming from the Los Alamitos ranch, Río Negro province, northeastern Patagonia (Bonaparte et al., 1984). This taxon is represented by several specimens, comprising cranial and postcranial remains of partial skeletons, which were recovered in the middle levels of Los Alamitos Formation, which is late Campanian or early Maastrichtian in age. K. australis was the most complete hadrosauroid species hitherto described for South America, and its assignment to the North American genus Kritosaurus was considered as a strong landmark in order to determine the provenance and time of arrival of this group of dinosaurs to South America (Bonaparte & Rougier, 1987; Bonaparte, 1996). A third unnamed hadrosauroid taxon was thought to be present in South America (Powell, 1987), described on the basis of an incomplete postcranial skeleton from the lower section of the Allen Formation, Salitral Moreno area, northern Patagonia. This specimen was considered as belonging to the Lambeosaurinae, contrasting with the "hadrosaurine" assignment proposed for Secernosaurus koerneri and Kritosaurus australis, thus providing additional evidence for an inter-American dispersal event at Campanian times (Bonaparte, 1996).
However, posterior authors suggested that Kritosaurus australis may not belong in fact to Kritosaurus, and the species was considered as "K." australis (Horner et al., 2004). Recent revisions of the material assigned to "Kritosaurus" australis led to the proposal that this purported species corresponds to the genus Secernosaurus, i.e. S. australis (Wagner, 2001), and more recently, to the hypothesis of its synonymy with the type species, Secernosaurus koerneri (Prieto-Márquez & Salinas, 2010). In this way, Kritosaurus australis is here considered as a junior synonym of Secernosaurus koerneri, following the latter suggestion.
In the present work, we describe new hadrosauroid materials from the Salitral Moreno area, coming from the same locality where the remains of the purported lambeosaurine were recovered (Juárez Valieri et al., 2007). Based on these specimens, as well as Powell's (1987) "lambeosaurine", and further material previously reported from the Allen Formation (González Riga & Casadío, 2000), we erect a new hadrosauroid taxon.
MATERIALS AND METHODS
We follow the nomenclature and definition of the clades among Hadrosauroidea described by Prieto-Márquez (2010), and the suggestions of the second edition of the compendium "The Dinosauria" by Weishampel et al. (2004) regarding anatomical orientations.
Anatomical abbreviations
ac: acetabulum, aig: cranial intercondylar groove, as: alveolar sulci, cf: coracoid foramen, cp: coronoid process, d: dentary, dc: deltopectoral crest, dpp: dorsal process of predentary, ds: distal shaft, en: external naris, er: edentulous region, f: frontal, fo: foramina, gl: glenoid, ilr: iliac ramus, ip: iliac peduncle, is: ischial shaft, mdp: maxillary dorsal process, mf: maxillary foramen, mc: medial carina (=primary carina), nc: neural channel, nf: nutrient foramen, ns: neural spine, p: parietal, pa: papilla, pb: pubic bar, pd: predentary, pdf: predentary facet, po: postorbital, pop: postacetabular process, pp: prepubis, pr: prezigapophysis , pra: pubic ramus, prp: preacetabular process, pz: postzygapophysis, rc: radial condyle, sa: surangular, sb: scapular blade, sf: splenial facet, sp: supraacetabular process, sq: squamosal, stf: supratemporal fenestra, sy: mandibular symphysis, th: teeth, tp: transverse process, uc: ulnar condyle, vpc: ventral process of coracoid.
Institutional abbreviations
MPHN-Pv: Museo Provincial de Historia Natural, Santa Rosa, La Pampa, Argentina, Colección de Paleontología de Vertebrados.
MPCA-Pv SM: Museo Provincial Carlos Ameghino, Cipolletti, Río Negro, Argentina, Paleontología de Vertebrados, Colección Salitral Moreno.
SYSTEMATICS
Ornithopoda Marsh 1881
Ankylopollexia Sereno 1986
Hadrosauroidea Sereno 1986
Saurolophidae (Brown 1914)
Willinakaqe gen. nov.
Etymology. The generic name is formed by the combination of the following words of the Mapuche language: willi meaning South, iná meaning mimic and kaqe meaning duck; the name thus meaning "the duck-mimic of the South".
Type species. The type and only known species of the genus is Willinakaqe salitralensis sp. nov.
Diagnosis. As for the type and only species (monotypic genus).
Willinakaqe salitralensis sp. nov.
Etymology. The name salitralensis refers to the Salitral Moreno locality, where the first remains were recovered.
Holotype. MPCA-Pv SM 8, a right premaxilla.
Paratypes. MPCA-Pv SM 2, partial skeleton including: dorsal, sacral and caudal vertebrae, ossified ligaments, haemapophyses, ribs, left scapula, right ilium and ischium, femora, tibiae, fibulae, calcanea and distal tarsals, and left astragalus and foot; MPCA-Pv SM 12, left maxillae with teeth; MPCA-Pv SM 03, right lower jaw, including dentary, surangular, angular, and teeth; MPCA-Pv SM 27, left scapula; MPCA-Pv SM 29, right coracoid; MPCA-Pv SM 32, left sternum; MPCA-Pv SM 33, left humerus; MPCA-Pv SM 39, left ilium, MPCA-Pv SM 41, right incomplete pubis.
Referred materials. MPCA-Pv SM 10, left maxilla; MPCA-Pv SM 4 to 6, left and right dentaries; MPCA-Pv SM 7 and 9, isolated teeth; MPCA-Pv SM 11, 13 and 14, cervical vertebrae; MPCA-Pv SM 15 to 22, dorsal vertebrae; MPCA-Pv 23 to 25, caudal vertebrae; MPCA-Pv SM 26, dorsal rib; MPCA-Pv SM 28, scapula; MPCA-Pv SM 30 and 31, coracoids; MPCA-Pv SM 34 to 38, humeri; MPCA-Pv SM 40, ilium; MPCA-Pv SM 42 to 44, femora; MPCA-Pv SM 45 and 46, tibiae; MPCA-Pv SM 47, fibula; MPCA-PV 48 to 51, metatarsals; MPCA-PV 52 to 55, phalanges and unguals; MPHN-Pv 01, partial single skeleton including cervical, dorsal, sacral, and caudal vertebrae, dorsal ribs, left scapula and coracoid, right femur, and two pedal phalanges, one of which is an ungual.
Diagnosis. Willinakaqe salitralensis gen. nov. et sp. nov. is diagnosed by the following combination of characters among Hadrosauroidea: premaxilla with a long and convex rostrolateral surface rostral to the narial fossa (autapomorphy); narial fossa shallow, with presence of a premaxillary foramen; caudodorsal and caudolateral processes of the premaxilla widely divergent, premaxillary denticles not well developed; dorsal vertebrae with a shallow fossa located on the lateral surface of the neural arch, upon the base of the transverse process (autapomorphy); eight sacral vertebrae in adults, the first with a ventral keel; high neural spines in sacral and proximal caudals, more than three times the height of the centrum; caudal neural spines showing a progressive distal expansion; dorsal border of the proximal section of the scapula straight; distal region of the postacetabular process of ilium ventrally deflected (autapomorphy); femur without enclosed cranial intercondylar groove.
Occurrence. The materials corresponding to the holotype, paratypes and most referred specimens of Willinakaqe salitralensis gen. nov. et sp. nov., housed at the Museo Provincial Carlos Ameghino, come from the southeastern region of the Salitral Moreno locality, in the north of the Río Negro province, 20 km. south of the General Roca locality, Argentina (Fig.1). These remains were recovered in a single bone-bed together with sauropod and ankylosaurian remains (Salgado & Azpilicueta, 2000; Coria & Salgado, 2001). The hadrosauroid remains are the most abundant in the assemblage, representing individuals of different ontogenetic stages, with estimated total lengths ranging between one and 9 meters. The larger specimens, as MPCA-Pv SM 2, display completely fused centra and neural arches in dorsal vertebrae and here are thus considered as presumable adult individuals (Irmis, 2007; Dalla Vecchia, 2009). Most of the preserved cranial, axial and appppendicular bones are represented by multiple elements, including partially articulated and closely associated pieces. Because all of them correspond to a single morphotype, the contention that these belong to a single taxon in the site is supported. The referred material housed at the Museo Provincial de Historia Natural de La Pampa, came from the Islas Malvinas locality, southwest of La Pampa province (Fig. 1). This specimen, MPHN-Pv 01, is considered as belonging to Willinake salitralensis gen. et sp. nov. because it preserves the same morphological traits, including these considered here as of diagnostic value in all the elements comparable with those from the type locality (see below). Both localities, Salitral Moreno and Islas Malvinas, correspond to the lower section of the Allen Formation (Malargüe Group), of late Campanian - early Maastrichtian age (Andreis et al., 1974; Casadío, 1994).
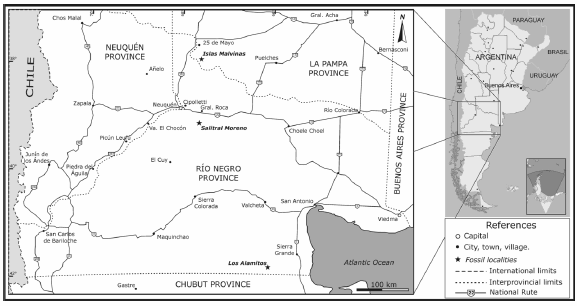
Fig. 1. Map indicating the northern Patagonian localities from which the hadrosauroid fossils here studied have been recovered (indicated as stars).
Description. In the present study we describe the elements that present useful morphological traits for the taxonomical and phylogenetic analysis of the species. Additional morphological traits of this taxon were codified by Prieto-Márquez (2010) for inclusion in their data matrix where it is informally named as "Salitral Moreno OTU" and are briefly treated in the Discussion section. A more detailed anatomical description of Willinakaqe salitralensis gen. nov. et sp. nov. will be published elsewhere.
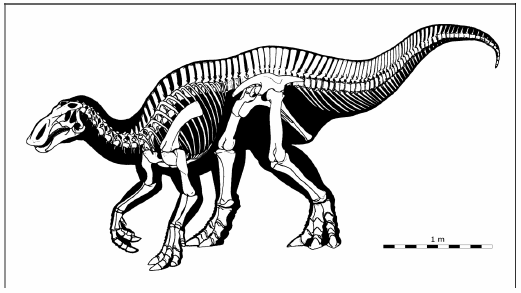
Fig. 2. Restoration of the skeleton of Willinakaqe salitralensis gen. et sp. nov. Preserved bones are white.
Cranial skeleton
Premaxilla. This bone (MPCA-Pv SM 8; Fig. 3A, B) is the most distinctive element available for Willinakaqe salitralensis gen. et sp. nov. It displays a suite of plesiomorphic and derived character states, whose combination is unique among Hadrosauroidea. Accordingly, this element was designed as the holotype of the new taxon. The premaxilla is almost complete, lacking the ventrolateral surface of the corner of the oral border and the caudal extremities of the caudodorsal and caudolateral processes. The denticles are poorly preserved in the available material and it is not possible to assert if they were as prominent as in basal hadrosauroids, such as Telmatosaurus transsylvanicus (Weishampel et al., 1993). Although a slight groove separates the denticle layer from the remaining premaxillary ventral surface, an inner palatal rim distinct from the denticle layer is not evident, as occurs in other saurolophids such as Prosaurolophus (Horner, 1992). In dorsal view, the oral and lateral margins are rounded and poorly transversely expanded. This morphology is also present in the primitive forms Eolambia caroljonesa, Bactrosaurus johnsoni, and Telmatosaurus transsylvanicus, and is not so transversely expanded as in the saurolophines, nor with the deflected rostroventral corner present in the lambeosaurines (Prieto Márquez et al., 2006). The area comprised between the oral margin and the rostroventral border of the circumnarial fossa is noticeably more extense than in basal forms like Eolambia, Telmatosaurus, or the lambeosaurines (Ostrom, 1961; Weishampel et al., 1993; Head, 2001). This area may be considered homologous with the area occupied by the external circumnarial fossa of the saurolophines, but Willinakaqe gen. nov. does not present the depression, nor the invaginated oral or lateral margins in this region, unlike the condition seen in Saurolophinae (Hadrosaurinae sensu Horner et al., 2004), and is considered as a possible autapomorphy of Willinakaqe salitralensis gen et sp. nov. (if not present also in near relatives such as Secernosaurus koerneri). A single foramen is present at the rostroventral end of the internal circumnarial fossa, which pierces the premaxillary body and opens at the ventral surface of the bone. This character is not currently known in any lambeosaurine, and is considered as a synapomorphy of the Saurolophinae (Horner et al., 2004; Prieto-Márquez, 2010). Two additional foramina are present on the rostral region of the dorsal surface of the premaxilla. At the caudal region of the premaxilla, the preserved portions of the caudodorsal and caudolateral processes do not present evidence of enclosing the narial passage, as occurs in lambeosaurines. Nevertheless, more complete skulls are necessary to confirm the latter assumption.

Fig. 3. Willinakaqe salitralensis gen. et sp. nov., cranial bones. A) and B) premaxilla of holotype specimen (MPCA-Pv SM 8) in A) dorsal and B) ventral view. C) to E) Maxilla of specimen (MPCA-Pv SM 12 in C) lateral, D) medial, and E) ventral views. Abbreviations in text.
Maxilla. Two almost complete maxillae were recovered from the Salitral Moreno locality (MPCA-Pv SM 10 and 12, Fig. 3C-E); the first corresponding possibly to an adult and the second to a subadult individual. This element is rostrocaudally elongated, with a rostrocaudally symmetrical morphology. The dorsal process of the maxilla is only slightly elevated, contrasting with the derived condition present in Lambeosaurinae, in which is dorsoventrally well-developed. In lateral view, a maxillary foramen is located below the base of the dorsal process and a second foramen is observed more caudally, with a ventral orientation. In medial view, the maxilla displays a continuous series of nutrient foramina. In general aspect, the maxilla of Willinakaqe gen. nov. is reminiscent to that described for Bactrosaurus johnsoni, Aralosaurus tuberiferus, and Pararhabdodon isonense (Godefroit et al., 1998, 2004; Prieto-Márquez et al., 2006).
Dentary. Two almost complete and two partially preserved dentaries are present among the Salitral Moreno material (MPCA-Pv SM 3 to 6; Fig. 4). This material includes elements of variable size. The dentary of Willinakaqe salitralensis gen. et sp. nov. displays a suite of characters proper of the derived hadrosauroids, but not apomorphic traits of Saurolophidae or any of their nested groups. The rostral region lacks an elongate symphysis, and shows a short diastema as present in basal hadrosauroids and some saurolophines and lambeosaurines (Horner et al., 2004). In lateral view, the ventral surface of dentary is weakly concave and subparallel with the straight dorsal margin at its mid-length. The predentary facet is almost vertical in lateral view. A series of foramina form a continuous line at mid-height on the lateral surface of the dentary. The coronoid process is subvertical, with a rostrocaudal expansion at its distal end, as usual in saurolophines and some lambeosaurines (Horner et al., 2004). In medial view, the dentary presents a series of subvertical parallel alveolar sulci. In all the preserved dentaries there are no more than thirty alveoli. The general morphology of the dentary of Willinakaqe salitralensis gen. et sp. nov. closely resembles that of Secernosaurus koerneri (Prieto-Márquez & Salinas, 2010).
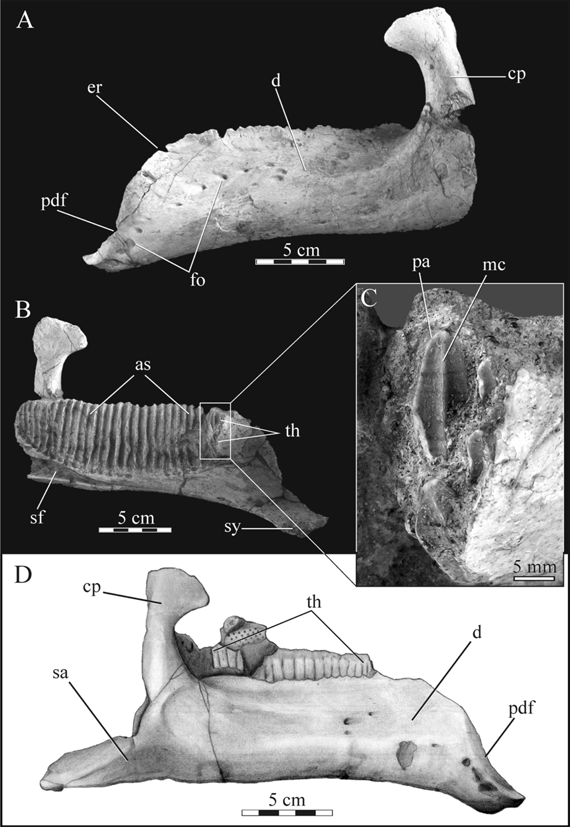
Fig. 4. Willinakaqe salitralensis gen. et sp. nov., lower jaw elements. A) lateral view of left dentary of specimen MPCA-Pv SM 6; B) medial view of the same element; C) closeup of B) to show medial surface of rostral tooth; C) lateral surface of right dentary of specimen MPCA-Pv SM 3. Abbreviations in text.
Teeth. Multiple teeth of Willinakaqe gen. nov. have been recovered adhered to some maxillae (MPCA-Pv 10 and 12) and dentaries (MPCA-Pv SM 3 and 4, Fig. 4B, C). Isolated dental elements are also present (MPCA-Pv SM 7 and 9), which are tentatively associated to Willinakaqe salitralensis gen. et sp. nov. because of their close association to the remaining cranial and postcranial material, and close similarity to the teeth preserved in articulation with bones. The dentary teeth are disposed in a dental battery with three pieces per alveolus. In the most rostral dentary teeth (MPCA-Pv SM 4, Fig. 4C), there is a complete absence of denticulations. These rostral dentary teeth present a single central carina and are mesiodistally symmetrical. In opposition, the teeth located in the middle and caudal section of the dental battery (MPCA-Pv SM 3) present small denticles on the distal borders. The morphology of these caudal dentary teeth is reminiscent to those found in the hadrosauroids from Antarctica described by Case et al. (2000) and some saurolophines (Horner et al., 2004).
Axial skeleton
Cervical vertebrae. The best preserved cervical vertebrae are those of MPHN-Pv 01, which have been previously described and illustrated by González Riga & Casadío (2000, Fig. 4A- C). Additional material came from the Salitral Moreno locality (MPCA-Pv SM 11, 13 and 14). The cervical vertebrae of Willinakaqe salitralensis gen. et sp. nov. displays the same basic morphology present in Telmatosaurus and more derived hadrosauroids. As originally noted by González Riga & Casadío (2000), the cervical vertebrae of this taxon can be distinguished from those of Secernosaurus on the basis of the angle of divergence between the postzygapophyses, which is greater in the latter taxon, and this represents one of the key characters allowing the association of the Salitral Moreno and the Islas Malvinas specimens in a single taxon.
Dorsal vertebrae. Isolated dorsal vertebrae comprising different regions of the column are present (MPCA-Pv SM 15 to 22, MPHN-Pv 01, Fig. 5A-H; González Riga & Casadío, 2000, Fig. 4D), as evidenced by the different angles between the transverse processes and neural spines and the size and orientation of the neural spines, which increases along the sequence and gets a more vertical orientation. Most of these vertebrae belong to juvenile individuals. As noted by González Riga & Casadío (2000) for the Islas Malvinas material, although their morphology is mostly usual for saurolophids, these vertebrae are different to those of Secernosaurus in the presence of a weak cavity located in the lateral face of the neural arc, upon the base of the transverse process (Fig. 5H). This trait is here considered as an autapomorphy of Willinakaqe salitralensis gen. et sp. nov.
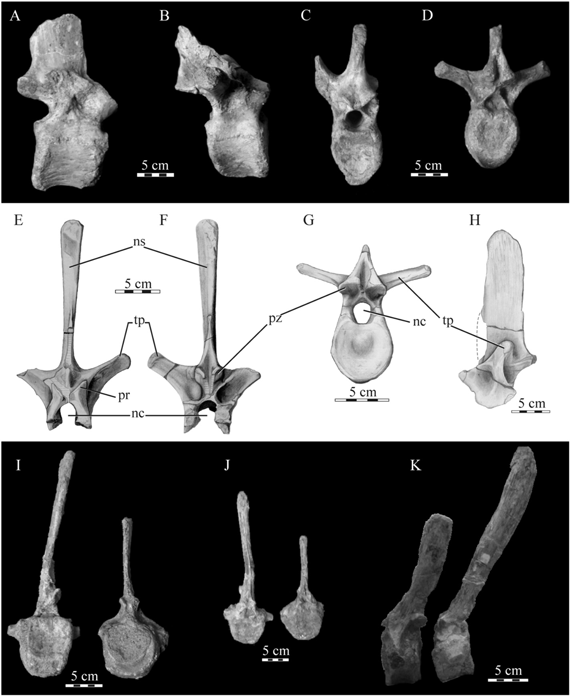
Fig. 5. Willinakaqe salitralensis gen. et sp. nov.; vertebrae. A-D) cranial dorsals, MPCA-Pv SM 15 and 16, in lateral (A and B) and caudal (C and D) views; E-H) caudal dorsals, MPCA-Pv SM 20-22, in cranial (E), caudal (F and G) and left lateral (H) views; I-K) proximal caudals, MPCA-Pv SM 23 and 24, in caudal (I), cranial (J), and left lateral (K) views. Abbreviations in text.
Sacral and caudal vertebrae. An articulated series of twenty five proximal and medial caudal vertebrae, continuing the complete sacral sequence, is present in the specimen MPCA-Pv SM 2 (Powell 1987, Fig. 2). Isolated caudals are also present in the Salitral Moreno (MPCA-Pv SM 23 to 25) and the Islas Malvinas material (MPHN-Pv 01, González Riga & Casadío 2000, Fig. 4F). The sacrum of an adult individual of Willinakaqe gen. nov., MPCN-Pv SM 2, is composed of eight fused vertebrae, as in Pararhabdodon isonense (Prieto-Márquez et al., 2006). This sacral number is low in comparison with most saurolophids, which present more than nine sacral vertebrae in adults (Horner et al., 2004). In Willinakaqe gen. nov., the first sacral vertebra presents a ventral keel, but it is not present in the following sacrals. All the sacral vertebrae, as well as the proximal caudal vertebrae, display dorsoventrally well-developed neural spines, with a height comprising 4.8 times the centrum height in the first caudal of the articulated sequence (Powell, 1987, Figs 2, 3). Additional isolated vertebrae from the Salitral Moreno locality, MPCA-Pv SM 23 and 24 (Fig. 5I-K), are considered here as proximal caudals due to the dorsal projection of their neural spines, and the similar ratio between the centrum and the neural spine heights. The caudal vertebrae recovered in the Islas Malvinas locality (González Riga & Casadío, 2000) preserve incomplete neural spines, but the morphology of the centrum and neural arch is identical to that of the other elements belonging to Willinakaqe gen. nov (see below). Another character also described by Powell (1987) and present in all the available elements is the progressive distal expansion of the neural spines, in both the proximodistal (along the axis of the tail) and transverse planes. A slight proximodistal (along the axis of the tail) constriction in the caudals at the base of the neural arch was observed by González Riga & Casadío (2000, Fig. 4F) in the Islas Malvinas material, and the same morphology is present in the caudal vertebrae from Salitral Moreno. These authors also noted the distinctive location of the postzygapophysis, situated dorsal to the prezygaphophysis, a condition also seen in all known specimens of Willinakaqe gen. nov. By contrast, Secernosaurus koerneri presents both zygaphophyses at almost the same level. The centra of the proximal caudal vertebrae are subcircular in contour in proximal view, with a convex ventral margin. Mid- and distal caudal vertebrae show a sagittal sulcus at their ventral surface.
Appendicular skeleton
Scapula. This bone is represented by four specimens, MPCA-Pv SM 2, 27, 28 and MPHN-Pv 01, the second of them being the most completely preserved (Fig. 6A, B). The entire morphology of the bone can be assessed on the basis of the currently available material. As mentioned by Powell (1987), the scapular blade is slightly distally expanded in adult and subadult individuals, as in Secernosaurus koerneri and non-saurolophid taxa, such as Bactrosaurus and Telmatosaurus (Godefroit et al., 1998; Weishampel et al., 1993). The distal border of the scapula is perpendicular to the shaft. In the proximal end, the scapula presents a large and prominent acromial bridge. A difference between the scapulae of Willinakaqe salitralensis gen. et sp. nov. and Secernosaurus koerneri is present in the dorsal border of the proximal region, which in the former is almost straight, as previously noted for the Islas Malvinas material by González Riga & Casadío (2000).

Fig. 6. Willinakaqe salitralensis gen. et sp. nov. A and B) scapula of specimen MPCA-Pv SM 2 in A) lateral and B) medial views. C and D) sternal plate of specimen MPCA-Pv SM in C) dorsal and D) ventral views.
Coracoid. Four coracoids are preserved: MPCA-Pv SM 29, which is completely preserved, MPCA-Pv SM 30 and 31, which preserve the entire bone except for the coracoidal hook, and MPHN-Pv 01, which comprises the cranial region of the bone. The morphology of the coracoid is closely similar to that found in Secernosaurus koerneri, with the coracoid foramen placed in the caudal region, completely surrounded by bone, and presenting a craniocaudally elongate oval shape. Bonaparte & Rougier (1987) considered this morp hology as diagnostic of "Kritosaurus" australis. The coracoid presents a convex craniodorsal corner. Ventral to this corner there is a fossa on the craniolateral border at mid-height of the bone. The coracoid is distally expanded and presents a strongly developed ventral coracoidal hook. This hook is similar in shape to that present in Secernosaurus, Telmatosaurus and Gryposaurus, more distally prominent than in Bactrosaurus, but not so developed as in Brachylophosaurus (Bonaparte & Rougier, 1987; Prieto-Márquez & Salinas 2010; Weishampel et al., 1993, 2004; Godefroit et al., 1998).
Sternum. An almost complete left sternal plate is preserved (MPCA-Pv SM 32, Fig. 6C, D). It presents a long and slender diaphysis, more elongated than that usually observed in saurolophids. At the proximal region, it expands up to the tip. The general shape is strongly reminiscent to that of Secernosaurus koerneri (Bonaparte & Rougier, 1987; Prieto-Márquez & Salinas, 2010).
Humerus. Six humeri of Willinakaqe salitralensis gen. et sp. nov. (MPCA-Pv SM 33 to 38) were recovered in the Salitral Moreno locality, representing different ontogenetic stages with sizes ranging between twelve and fifty five centimeters in proximodistal length. The most remarkable feature is the presence of a deltopectoral crest which is well-developed in proximodistal and mediolateral senses (Fig. 7), which resembles that seen in Tanius, Pararhabdodon,and the lambeosaurines, contrasting with the condition present in Telmatosaurus, Hadrosaurus, and the saurolophines. However, as observed by Prieto-Márquez et al. (2006) the presence of a prominent deltopectoral crest is a homoplastic character and not necessarily a lambeosaurine synapomorphy.
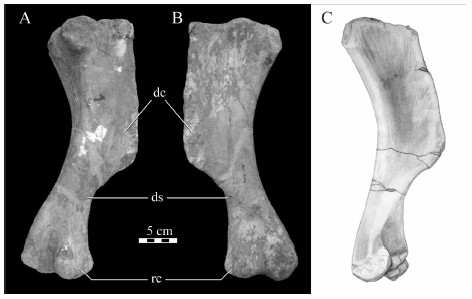
Fig. 7. Willinakaqe salitralensis gen. et sp. nov. Right humerus of specimen MPCA-Pv SM 33 in A) caudolateral and B) craniomedial views. C) Left humerus of specimen 38 in craniomedial view. Abbreviations in text.
Ilium. Three ilia have been recovered, MPCA-Pv SM 2, 39 and 40, of which the first two are almost complete. The general morphology of this bone (Fig. 8) is comparable to those of derived hadrosauroids, in the presence of a deflection on the dorsal surface and the occurrence of a well-developed postacetabular process. The supracetabular process is wide and prominent, as reported by Powell (1987) for MPCA-Pv SM 2, and the distal rim slightly bows ventrolaterally. This results in a dorsally convex and ventrally concave antitrochanter. The postacetabular process of the ilium is wide and presents a distinctive caudoventral curvature at the distal region. The overall morphology of the ilium of Willinakaqe salitralensis gen. et sp. nov. more closely resembles that of Secernosaurus koerneri than that of other hadrosauroids (see below).
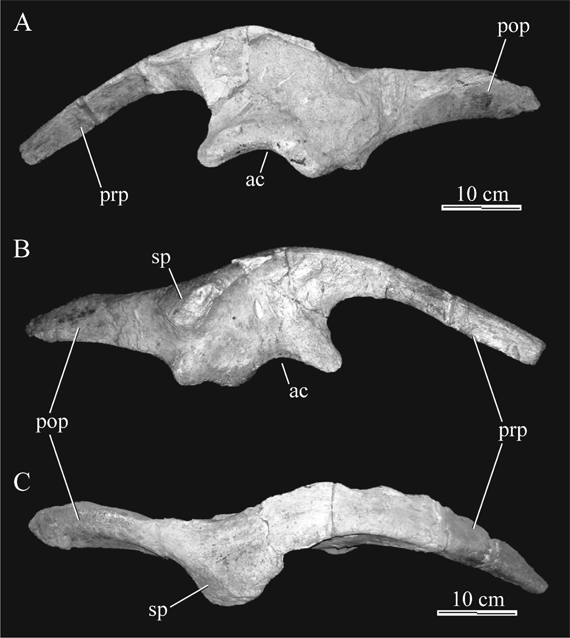
Fig. 8. Willinakaqe salitralensis gen. et sp. nov. Right ilium of specimen MPCA-Pv SM 39 in A) medial, B) lateral, and C) dorsal views. Abbreviations in text.
Ischium. A complete ischium is preserved in MPCA-Pv SM 2, which was briefly described by Powell (1987). The shaft is entirely straight, and the distal end does not present a foot-like expansion in this adult individual, exhibiting the same general morphology of Secernosaurus, but contrasting with the condition present in Pararhabdodon and several saurolophids, which show a well developed distal foot in adult individuals (Horner et al., 2004; Guenther, 2009).
Pubis. An incomplete pubis comprising most of the prepubic blade is preserved (MPCA-Pv SM 41, Fig. 9A, B). It bears a dorsoventrally convex lateral surface and a planar medial one. The blade presents its maximum dorsoventral expansion in the middle of the shaft. From this point, the dorsal and ventral borders converge cranially, resulting in a non-parallel pattern, unlike the condition present in Secernosurus and Gryposaurus (Prieto-Márquez, 2010: appendix).
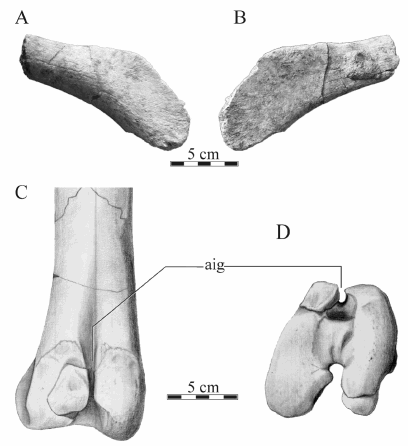
Fig. 9. Willinakaqe salitralensis gen. et sp. nov. Right prepubic process of specimen MPCA-Pv SM 41 in A) lateral and B) medial views. Left femur of specimen MPCA-Pv SM 42 in C) cranial and D) distal views. Abbreviations in text.
Femur. Five femora are preserved. Complete femora representing adult and subadult individuals are preserved in MPCA-Pv SM 2, 42 and 43, with sizes ranging from 35 centimeters to near a meter in proximodistal length. The overall femoral morphology is similar to that of other hadrosauroids. There is no complete enclosure of the cranial intercondylar groove by the projections of the condyles (Fig. 9C, D), contrasting with the condition present in Secernosaurus koerneri (Bonaparte & Rougier, 1987). The character does not appear to be ontogenetically variable in Willinakaqe gen. nov., because the open condition is present in all the subadult and adult specimens. This character is considered as homoplastic among hadrosauroids, but valid for specific differentiation (Godefroit et al., 1998).
DISCUSSION AND CONCLUSIONS
Willinakaqe salitralensis gen. et sp. nov. and Secernosaurus koerneri exhibit several anatomical similarities, mainly symplesiomorphies, but several of these allow differentiating these South American species from hadrosauroids less derived than Telmatosaurus transsylvanicus or Tanius sinensis, and from the different saurolophine and lambeosaurine subclades. Following to Prieto-Márquez (2010), Willinakaqe salitralensis gen. et sp. nov. can be included in the Saurolophidae and more inclusive clades among Hadrosauroidea on the basis of the following character states: location of the origin of the ventral deflection of the dentary near to the origin of the dental battery; bulging of the ventral lateral margin of the dentary straight or slightly bowed rostral to the coronoid process; thick and dorsoventrally elongated ridge on the medial side of the coronoid process absent; dentary lingual arching of the occlusal plane absent, with rostrocaudally straight occlusal plane; length of the ectopterygoid shelf relative to the total rostrocaudal length of the alveolar margin of the maxilla greater than 0.35; development of the postzygapophyseal proceses of cranial and middle cervical vertebrae three times or more longer than the breadth of the neural arch; length of the deltopectoral crest of the humerus developed greater than 55 percent of the proximodistal length of the humerus; lateroventral expansion of the deltopectoral crest of the humerus well expanded; ventral margin of the deltopectoral crest extending abruptly from the humeral shaft to give a distinct angular profile; ventral deflection of the preacetabular process of the ilium with an angle of less than 150°; supraacetabular process craniocaudally broad; morphology of the pubic peduncle of the ilium relatively short and triangular, with a proximal region that is craniocaudally much wider than the distal end; ischial peduncle of the ilium formed by two protrusions of similar size; postacetabular process of the ilium with a caudodorsally oriented dorsal margin, rising dorsally relative to the acetabular margin; distal articular surface of the iliac peduncle of the ischium cranially oriented; and acetabular and caudodorsal margins of the iliac peduncle of the ischium becoming slightly to greatly divergent near the proximal region of the peduncle. Willinakaqe salitralensis gen. et sp. nov. also displays the following synapomorphies of Saurolophinae: presence of a premaxillary foramen located rostrally and ventrolaterally to the rostral margin of the external naris; and pubic peduncle of ischium parallel with the ischial shaft. Willinakaqe gen. nov. displays the following characters supporting its inclusion into the unnamed clade comprising those taxa closer to Gryposaurus than to Brachyolophosaurus in the phylogeny published by Prieto-Márquez (2010, figure 5): Dorsoventral depth of the central plate of the ilium low; and ratio between the length of metatarsal III and its mediolateral breadth at midshaft lesser than 4.50. No unambiguous synapomorphies are present in all the resulting trees of this analysis supporting the association between the Gryphosaurus species and Willinakaqe salitralensis gen. et sp. nov. and their closer relatives, the unnamed Big Bend taxon of Prieto-Márquez (2010) and Secernosaurus koerneri. Four characters associate these latter three species: coracoid with slightly longer scapular facet respect to the lateral margin of the glenoid; presence of a brevis shelf at the base of the postacetabular process of the ilium (see discussion in Prieto-Márquez's character 244); iliac medioventral ridge on the medial side of the postacetabular process well-developed, oblique, and expanded into a flange forming the medial margin of an extensive brevis shelf that faces more ventrally than medially; and presence of a craniocaudally oriented median ridge on the dorsolateral surface of the postacetabular process of the ilium. Finally, Willinakaqe salitralensis gen. et sp. nov. is supported as the sister taxon of Secernosaurus koerneri by five character states in the Prieto-Márquez analysis: height of the neural spine greater than 2.10 times that of the centrum in the tallest caudal dorsal or sacral vertebrae; angle greater than 115° between the lateral margin of the facet for scapular articulation and the glenoid in the coracoid; supra-acetabular process of the ilium asymmetrical, with a caudally skewed lateral profile; caudodorsal margin of ilium with a well-defined ridge continuous with the dorsal margin of the proximal region of the postacetabular process; mid-shaft of the ischium with a very thick shaft.
Some characters indicating a derived placement for Willinakaqe gen. nov. and Secernosaurus in relation to Telmatosaurus and more basal taxa includes: diamond-shaped dentary teeth with only one straight medial keel and with small denticles; dentary with more than thirty alveoli and with more than two teeth position; strong inflection of the dentary dorsal border forming a low mandibular symphysis; coronoid process of the dentary bone vertically oriented, lacking a rostralward inclination; scapular blade slightly distally expanded; sternal caudoventral process well developed, relatively long, and thick relative to the sternal 'handle'; ilium with dorsal border displaying an inflexion between a convex cranial region and a concave caudal one upon the acetabulum; large and extensive supraacetabular process located dorsal to the acetabulum; prominent postacetabular process of the ilium; ischial shaft straight and caudoventrally oriented; ischium without foot-like expansion. Likewise, Willinakaqe gen. nov. and Secernosaurus present several characters which could be considered as plesiomorphic relative to Saurolophidae, such as a jugal with a ventral margin of the rostral process dorsal to level of lateral ridge of the ectopterygoid shelf, a maxilla-jugal contact restricted to a finger-like jugal process on the caudal margin of maxilla, less than thirty-five tooth positions in both dentary and maxillary dental batteries during ontogeny, no less than four foramina arranged in a sub-horizontal row along the lateral wall of the maxilla, and less than nine sacrals.
The new taxon Willinakaqe salitralensis represents the second hadrosauroid species recognized from South America, because it displays several differences with Secernosaurus koerneri, which contains the material previously assigned to "Kritosaurus" australis ( Prieto-Márquez & Salinas, 2010). These differences include the cervical vertebrae with a lower angle of divergence between the postzygapophyses; dorsal vertebrae with a shallow fossa positioned on the lateral surface of the neural arch, upon the base of the transverse process; tall neural spines in sacral and proximal caudals more than four times the centrum height; presence of a ventral keel in the proximal sacral vertebrae; the postzygapophysis of the proximal caudal vertebrae situated at a level dorsal to the prezygaphophysis; distal region of the postacetabular process of the ilium ventrally deflected; and femur with open cranial intercondylar groove, among others. Other characters diagnostic of Willinakaqe salitralensis gen. et sp. nov., as those present in the premaxilla, are not comparable with those of Secernosaurus koerneri because this element is unknown in the latter genus.
Based on the phylogenetic scheme of the hadrosauroids presented by Prieto-Márquez (2010), Willinakaqe salitralensis gen. et sp. nov. (their Salitral Moreno OTU) is found as the sister taxon to Secernosaurus koerneri within Saurolophinae. This analysis present decay index values of only one step, and bootstrap frequency values of less than fifty percent, for support of all the successive clades including the South American taxa with other saurolophines different of the "Big Bend Moreno" taxon. Furthermore, no less than nine reversal events are necessary along the hypothesized evolutionary history of the clade comprising Willinakaqe gen. nov., Secernosaurus and the Big Bend Moreno taxon to support their derived position within Saurolophinae. A more detailed study of the South American forms and additional specimens are necessary in order to confirm the position of both Secernosaurus and Willinakaqe gen. nov.
The presence of hadrosauroid remains in the Late Cretaceous of Patagonia is of particular interest for understanding the paleobiogeographic dispersal events occurring during this timespan, given that this group of dinosaurs is considered as being of Laurasian origin (Sues & Averianov, 2009). The original referral of the material from the Los Alamitos Formation to the genus Kritosaurus (Bonaparte et al., 1984) and the purported lambeosaurine condition of MPCA-Pv SM 2 advanced by Powell (1987) suggested than at least two different groups of hadrosauroids dispersed to South America during the Late Cretaceous. The phylogenetic placement of the South American hadrosauroids presented by Prieto-Márquez (2010) suggests only a dispersal event for the group followed by a local radiation. Whether or not these biogeographical events ocurred during the Campanian, and through the Isthmus of Panama or an alternative route, is still uncertain, pending of a better understanding of the Late Cretaceous terrestrial vertebrate faunas from South America.
To date, all the diagnostic hadrosauroid material recovered from the Allen Formation of the Neuquén Basin belong to a single taxon, Willinakaqe salitralensis gen. et sp. nov., suggesting a slight temporal difference between this stratigraphic unit and those containing the taxon Secernosaurus australis.
ACKNOWLEDGEMENTS
We express our gratitude to Alejandro Kramarz and Carlos Muñoz for permission to revise the collections under their care, to Guillermo Salinas, Juan Porfiri, Fernando Novas, and Andrea Cambiaso for comments and revision of the manuscript and provision of photographs, to Previterio Pacheco, Elías Gómez, Martín Berrondo, and Magalí Cárdenas for preparation of materials and to Martín Berrondo, for some of the illustrations. Finally, we want to acknowledge the useful corrections and comments of two anonymous reviewers. Fieldwork research project funded by the Universidad Nacional del Comahue (JOC) and Museum Carlos Ameghino of the city of Cipolletti.
BIBLIOGRAPHY
1. Andreis, R.R., A.M. Iñíguez Rodríguez, J.J. Lluch & D.A. Sabio. 1974. Estudio sedimentológico de las formaciones del Cretácico Superior del área del Lago Pellegrini (prov. de Río Negro, Rep. Argentina). Rev. Asoc. Geol. Arg., 29: 85-104. [ Links ]
2. Bonaparte, J.F. 1984. Nuevas pruebas de la conexión física entre Sudamérica y Norteamérica en el Cretácico Tardío (Campaniano). IIIº Congr. Arg. Paleont. y Bioestrat., Actas: 141-149. [ Links ]
3. Bonaparte, J.F. 1996. Cretaceous Tetrapods of Argentina. Münch. Geowiss. Abhandl. (A), 30: 73-130. [ Links ]
4. Bonaparte, J.F. & G.W. Rougier. 1987. The Late Cretaceous fauna of Los Alamitos, Patagonia, Argentina. Part VII - The hadrosaurs. Rev. Mus. Arg. Cienc. Nat. B. Rivadavia, 3(3): 155-161. [ Links ]
5. Bonaparte, J.F., M.R. Franchi, J.E. Powell & E.G. Sepúlveda. 1984. La Formación los Alamitos (Campaniano - Maastrichtiano) del sudeste de Río Negro, con descripción de Kritosaurus australis n. sp. (Hadrosauridae). Significado paleogeografico de los vertebrados. Rev. Asoc. Geol. Arg., 39(3-4): 284-289. [ Links ]
6. Brett-Surman, M.K. 1979. Phylogeny and palaeobiogeography of hadrosaurian dinosaurs. Nature, 277: 560-562. [ Links ]
7. Brown, B. 1914. Corythosaurus casuarius, a new crested dinosaur from the Belly River Cretaceous, with provisional classification of the family Trachodontidae. Bull. Amer. Mus. Nat. Hist., 33: 559-565. [ Links ]
8. Casadío, S.A. 1994. Estratigrafía y Paleontología del intervalo Maastrichtiano-Daniano en el occidente de la provincia de La Pampa, Argentina. Inedit Doctoral Thesis. Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Argentina. 420pp. [ Links ]
9. Casamiquela, R.M. 1964. Sobre un dinosaurio hadrosáurido de la Argentina. Ameghiniana, 3: 285-308. [ Links ]
10. Case, J.A., J.E. Martin, D.S. Chaney, M. Reguero, S.A. Marenssi, S.M. Santillana, & M.O. Woodburne. 2000. The first duck-billed dinosaur (Family Hadrosauridae) from Antarctica. J. Vert. Paleont., 20(3): 612-614. [ Links ]
11. Coria, R.A. & L. Salgado. 2001. South American Ankylosaurs. En: K. Carpenter (ed.), The Armored Dinosaurs, pp. 159-168, Indiana University Press, Bloomington. [ Links ]
12. Dalla Vecchia, F.M. 2009. Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy. J. Vert. Paleont., 29(4): 1100-1116. [ Links ]
13. Godefroit, P., Z-M. Dong, P. Bultynck, H. Li & L. Feng. 1998. Sino-Belgian Cooperative Program. Cretaceous Dinosaurs and Mammals from Inner Mongolia. 1. New Bactrosaurus (Dinosauria: Hadrosauroidea) material from Iren Dabasu (Inner Mongolia, P.R. China). Bull. Inst. Roy. Sci. Nat. Belg. Sci. Terre, 68(Suppl.): 1-70. [ Links ]
14. Godefroit, P., V. Alifanov & Y. Bolotsky. 2004. A re-appraisal of Aralosaurus tuberiferus (Dinosauria, Hadrosauridae) from the Late Cretaceous of Kazakhstan. Bull. Inst. Roy. Sci. Nat. Belg. Sci. Terre, 74(Suppl.): 139-154. [ Links ]
15. González Riga, B.J. & S.A. Casadío. 2000. Primer registro de Dinosauria (Ornithischia, Hadrosauridae) en la provincia de La Pampa (Argentina) y sus implicancias paleobiogeográficas. Ameghiniana, 37(3): 341-351. [ Links ]
16. Guenther, M.F. 2009. Influence of sequence heterochrony on hadrosaurid dinosaur postcranial development. Anat. Rec., 292: 1427-1441. [ Links ]
17. Head, J.J. 2001. A reanalysis of the phylogenetic position of Eolambia caroljonesa (Dinosauria, Iguanodontia). J. Vert. Paleont., 21(2): 392-396. [ Links ]
18. Horner, J.R. 1992. Cranial morphology of Prosaurolophus (Ornithischia: Hadrosauridae) with descriptions of two new hadrosaurid species and an evaluation of hadrosaurid phylogenetic relationships. Mus. Rockies Occ. Pap., 2: 1-119. [ Links ]
19. Horner, J.R., D.B. Weishampel & C.A. Forster. 2004. Hadrosauridae. En: D.B. Weishampel, P. Dodson & H. Osmólska (Eds.), The Dinosauria, Second Edition, pp. 438-463, University of California Press, Berkeley. [ Links ]
20. Irmis, R.B. 2007. Axial skeleton ontogeny in the Parasuchia (Archosauria: Pseudosuchia) and its implications for ontogenetic determination in archosaurs. J. Vert. Paleont., 27: 350-361. [ Links ]
21. Juárez Valieri, R.D., J.O. Calvo, J.C. Muñoz, G.C. Salinas & L. E. Fiorelli. 2007. New hadrosaur materials from Salitral Moreno, Allen Formation, Río Negro province, Argentina. XXIIIº Jor. Arg. Paleont. Vert., Resúmenes: 18. [ Links ]
22. Marsh, O.C. 1881. Principal characters of American Jurassic dinosaurs. Pt. IV. Spinal cord, pelvis, and limbs of Stegosaurus. Am. J. Sci. (ser. 3), 21: 167-170. [ Links ]
23. Ostrom, J.H. 1961. Cranial morphology of the hadrosaurian dinosaurs of North America. Bull. Am. Mus. Nat. Hist., 122: 33-186. [ Links ]
24. Powell, J.E. 1987. Hallazgo de un dinosaurio hadrosáurido (Ornithischia: Ornithopoda) en la Formación Allen (Cretácico Superior) de Salitral Moreno, Prov. de Río Negro, Argentina. Xº Congr. Geol. Arg., Actas,. 3: 149-152. [ Links ]
25. Prieto-Márquez, A. 2010. Global phylogeny of Hadrosauridae (Dinosauria: Ornithopoda) using parsimony and Bayesian methods. Zool. J. Linn. Soc., 159: 435-502. [ Links ]
26. Prieto-Márquez, A. & G. Salinas. 2010. A re-evaluation of Secernosaurus koerneri and Kritosaurus australis (Dinosauria, Hadrosauridae) from the Late Cretaceous of Argentina. J. Vert. Paleont., 30: 813-837. [ Links ]
27. Prieto-Márquez, A., R. Gaete, G. Rivas, A. Galobart & M. Boada. 2006. Hadrosauroid dinosaurs from the Late Cretaceous of Spain: Pararhabdodon isonensis revisited and Koutalisaurus kohlerorum, gen. et. sp. nov. J. Vert. Paleont., 26(4): 929-943. [ Links ]
28. Salgado, L. & C. Azpilicueta. 2000. Un nuevo saltasaurino (Sauropoda, Titanosauridae) de la provincia de Río Negro (Formacion Allen, Cretácico superior), Patagonia, Argentina. Ameghiniana, 37(3): 259-264. [ Links ]
29. Sereno, P.C. 1986. Phylogeny of the bird-hipped dinosaurs (order Ornithischia). Nat. Geogr. Res., 2(2): 234-256. [ Links ]
30. Sues, H.-D. & A. Averianov. 2009. A new basal hadrosauroid dinosaur from the Late Cretaceous of Uzbekistan and the early radiation of duck-billed dinosaurs. Proc. R. Soc. B, 276: 2549-2555. [ Links ]
31. Wagner, J.R. 2001. The hadrosaurian dinosaurs (Ornithischia: Hadrosauria) of Big Bend National Park, Brewster County, Texas, with implications for Late Cretaceous paleozoogeography. Inedit Master's Thesis. Texas Tech University, EEUU. 417pp. [ Links ]
32. Weishampel, D.B., D.B. Norman & D. Grigorescu. 1993. Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: the most basal hadrosaurid dinosaur. Palaeontology, 36(2): 361-385. [ Links ]
33. Weishampel, D.B., P. Dodson & H. Osmolska. 2004. Introduction. En: D.B. Weishampel, P. Dodson & H. Osmólska (Eds.), The Dinosauria, Second Edition, pp. 1-3, University of California Press, Berkeley. [ Links ]
Recibido: 19-II-2010
Aceptado: 18-XI-2010

