










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
versión On-line ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.13 no.2 Ciudad Autónoma de Buenos Aires jul./dic. 2011
PALEONTOLOGÍA
El registro de maderas fósiles de Leguminosae de Sudamérica
Roberto R. Pujana1, Leandro C. A. Martínez1 & Mariana Brea2
1 Museo Argentino de Ciencias Naturales. Av. A. Gallardo 470. D1405DJR Buenos Aires, Argentina. rpujana@macn.gov.ar.
2 CICYTTP-CONICET. Matteri y España s/n. E3105BWA Diamante, Entre Ríos, Argentina.
Resumen: La familia Leguminosae posee un extenso registro de maderas fósiles en todo el mundo. En Sudamérica se reconocen 56 registros afines a la familia. Se enlistan todos los registros conocidos y se incluyen algunos datos inéditos. La familia Leguminosae es la de mayor diversidad paleoxilológica de Sudamérica. La misma posee un extenso registro paleoxilológico temporal, desde el Paleoceno hasta el Pleistoceno, y geográfico, desde Patagonia hasta el norte de Colombia.
Palabras clave: Leguminosae; Fabaceae; Maderas fósiles; Revisión; Sudamérica.
Abstract: The fossil wood record of Leguminosae from South America. The Leguminosae has an extensive fossil wood record worldwide. In South America 56 records with affinity to Leguminosae were counted. Each record is analyzed and unpublished data is also included. The Leguminosae is the most palaeoxylological diverse family of South America. It has an extensive temporal record, from the Palaeocene to the Pleistocene, and a broad geographical span, from Patagonia to north Colombia.
Key words: Leguminosae; Fabaceae; Fossil wood; Review; South America.
INTRODUCCIÓN
La familia Leguminosae Jussieu, nom. cons. et nom. alt. o Fabaceae Lindley, nom. cons. se distribuye actualmente en todos los continentes a excepción de la Antártida. Comprende aproximadamente 720 géneros y con unas 18000 especies es la tercer familia de angiospermas en cuanto a su número de representantes (Lewis et al., 2005). Su registro fósil es extenso, con hallazgos de leños, hojas, polen, estructuras repoductivas y frutos, con cientos de morfoespecies asignadas a sus tres subfamilias: Mimosoideae, Caesalpinioideae y Papilionoideae (Herendeen, 1992).
Haciendo referencia a su registro paleoxilológico, en todos los continentes, excepto Antártida y Oceanía se conocen numerosas maderas fósiles asignadas a las tres subfamilias (Müller Stoll & Mädel, 1967; Gros, 1992; Wheeler & Bass, 1992; Gregory et al., 2009). El registro fósil de maderas de esta familia en Sudamérica es relativamente extenso. La anatomía del leño es muy diagnóstica y se caracteriza por tener elementos de vaso relativamente grandes, puntuaciones intervasculares alternas, placas de perforación simples y parénquima axial.
El objetivo de esta revisión es reunir los datos paleoxilológicos de las Leguminosae en Sudamérica, debido a que algunos de ellos son inéditos o de difícil acceso. Finalmente se obtienen conclusiones acerca de la historia y diversidad de esta familia en América del Sur.
MATERIALES Y MÉTODOS
La metodología utilizada en este trabajo se basó en la búsqueda y revisión de la bibliografía donde se citan y referencian, en base a su anatomía, maderas fósiles afines a las Leguminosae de Sudamérica incluyendo: resúmenes y actas de congresos, simposios y reuniones, tesis y algunos datos inéditos de los autores de este trabajo. Además, se consultó el catálogo de Gregory et al. (2009), Inside Wood (2004) y otras revisiones sobre leños fósiles de Leguminosae como las realizadas por Gros (1992) y Müller Stoll & Mädel (1967), adoptando los criterios propuestos por dichos autores en la nomenclatura de las morfoespecies y morfogéneros.
En la Tabla 1, se encuentra la totalidad de los registros con su nomenclatura actualizada. Cada registro constituye un xilotipo para una unidad estratigráfica determinada. A su vez cada xilotipo se corresponde con una unidad taxonómica, haya sido o no nombrada o válidamente publicada. Se considera como un mismo registro la presencia de un xilotipo en dos o más localidades donde aflora la misma Formación.
Tabla 1. Listado de los xilotipos de Leguminosae de Sudamérica.
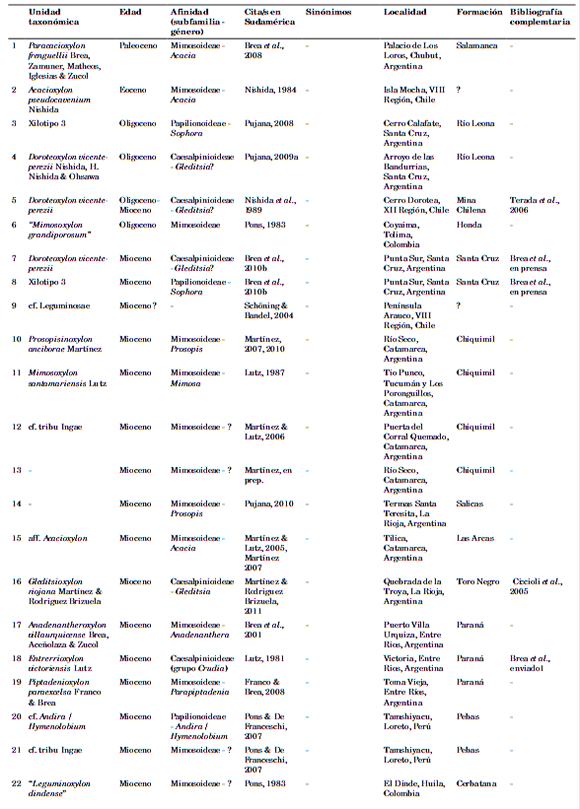
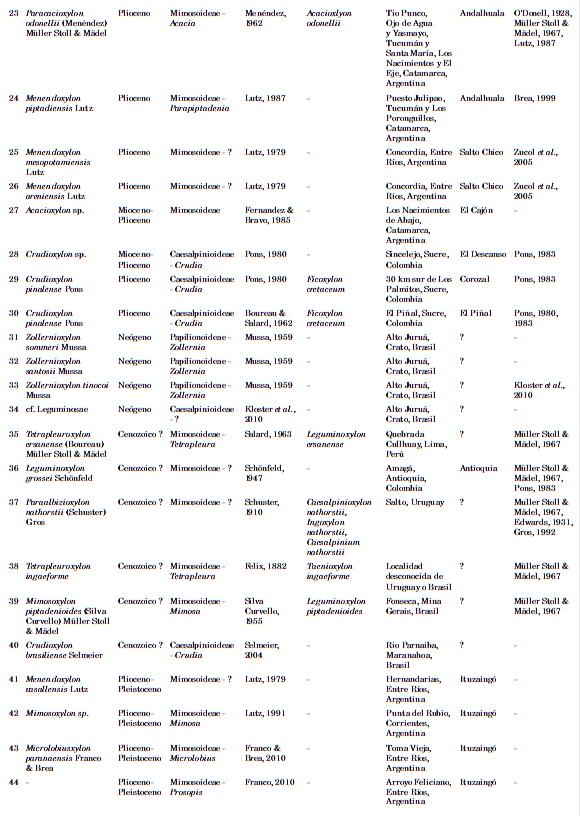

REGISTRO FÓSIL
La familia Leguminosae tiene aún un origen geográfico incierto, si bien se cuentan con evidencias desde el Cretácico Superior basadas principalmente en datos palinológicos (Raven & Polhill, 1981). No obstante, los registros más antiguos de leños certeramente asignados a esta familia proceden del Paleoceno (Wheeler & Bass, 1992; Brea et al., 2008).
A continuación se mencionan las citas sobre maderas de leguminosas en América del Sur en un orden cronológico.
Paleógeno
Entre los registros más antiguos de Leguminosae, se halla Paracacioxylon frenguellii Brea et al., (2008), el cual fue recolectado en sedimentos de la Formación Salamanca, proveniente del Paleoceno inferior (Daniano) de la provincia del Chubut, Argentina. Esta madera afín a Acacia Miller posee porosidad semi-circular a circular, radios multiseriados de hasta 6 células de espesor y abundante parénquima axial.
Del Eoceno de Sudamérica únicamente se describió Acacioxylon pseudocavenium Nishida (1984) de la Isla Mocha, de la VIII región de Chile. La misma se caracteriza por poseer vasos solitarios o en series radiales, radios 1-3 seriados y abundante parénquima axial (Nishida, 1984).
Provenientes del Oligoceno patagónico, se describieron dos xilotipos. El primero, Doroteoxylonvicente-perezii Nishida et al. (1989) fue descripto por primera vez para la Patagonia chilena a partir de un ejemplar recolectado en la localidad Cerro Dorotea, proveniente de sedimentos de la Formación Mina Chilena, con una edad estimada oligocena superior-miocena inferior; posteriormente Terada et al. (2006) mencionan nuevos ejemplares de la misma localidad. Estos autores le asignan una afinidad probable a las Proteaceae debido a los radios anchos y las supuestas bandas de vasos en el leño temprano, sin embargo consecutivamente Pujana (2009a) describe un ejemplar de una localidad argentina cercana y de edad oligocena con excelente estado de preservación y lo reasigna a las Leguminosae, probablemente a la subfamilia Caesalpinoideae. De los mismos sedimentos oligocenos, pertenecientes a la Formacion Río Leona, Pujana (2008) describe el "xilotipo 3" afín a la sección Edwardsia Seem. del género Sophora L., caracterizado por radios 5-6 seriados, y vasos pequeños dispuestos en bandas tangenciales a irregulares dendríticas y parénquima radial disyunto. "Mimosoxylon grandispososum" Pons (1983) recolectada en sedimentos oligocenos de la Formación Honda, en Coyaima, Colombia es la única madera paleógena de Leguminosae hallada fuera de Patagonia e islas adyacentes. Se caracteriza por poseer vasos grandes, radios normalmente triseriados y parénquima cristalífero. Sin embargo su publicación no es efectiva debido a que su descripción se encuentra en una tesis inédita (Art. 29 ICNB, Mcneill et al., 2006).
Neógeno
Las maderas fósiles de Leguminosae son mucho más abundantes en el Neógeno respecto al Paleógeno, particularmente en el Noroeste Argentino y Mesopotamia Argentina.
En Patagonia, recientemente Brea et al. (2010b; en presa) citan la presencia de Doroteoxylon vicente-perezii y el "xilotipo 3" provenientes de la Formación Santa Cruz (Mioceno inferior). En el centro de Chile, Schöning & Bandel (2004) describen brevemente una posible madera de Leguminosae con caracteres típicos de la familia, placas de perforación simples, vasos relativamente grandes y parénquima paratraqueal.
Provenientes de sedimentos del Mioceno, en el Grupo Santa María (noroeste de Argentina), se han descripto numerosas maderas afines a las Leguminosae. Mimosoxylon santamariensis Lutz (1987), procedente de las Formaciones San José (Mioceno medio) y Chiquimil (Mioceno superior), se caracteriza por sus vasos generalmente solitarios y radios homogéneos predominantemente biseriados siendo afín al género Mimosa L. según la autora. Martínez & Lutz (2006) describen de la Formación Chiquimil un leño con afinidad a la tribu Ingae y correspondiente probablemente a un nuevo morfogénero con vasos solitarios, radios 1-2 seriados y parénquima escaso. De la misma formación fue recolectado Prosopisinoxylon anciborae Martínez (2010), caracterizado por poseer vasos de dos tamaños distintos y radios anchos (Martínez, 2007, 2010). Otros restos asignados a este morfogénero son los recientemente descriptos por Pujana (2010) de la Formación Salicas (Mioceno Superior) en la provincia de La Rioja. Martínez (en prep.) actualmente se encuentra describiendo materiales afines a Mimosoideae provenientes de la misma localidad donde fue recolectado Prosopisinoxylon anciborae. De la Formación Las Arcas (Mioceno Medio) aflorante en la provincia de Catamarca, Martínez & Lutz (2005) y Martínez (2007) describieron ejemplares probablemente afines a Acacia (Mimosoideae) con radios 4-10 seriados y parénquima cristalífero con grandes cristales.
La Mesopotamia argentina también porta sedimentos de edad miocena, tal es el caso de la Formación Paraná (Mioceno Superior), de la cual proceden Entrerrioxylon victoriensis Lutz (1981), Anadenantheroxylon villaurquisense Brea et al. (2001) y Piptadenioxylon paraexcelsa Franco & Brea (2008). Entrerrioxylon victoriensis fue establecida como Papilionoidea por Lutz (1981) debido a determinados caracteres como la disposición de los vasos en patrón dendrítico, elementos de vasos de dos tamaños, presencia de traqueidas vasculares, parénquima axial escaso y estratificación parcial. Brea et al. (enviado 1) le asignan una afinidad a las Caesalpinoideae y dentro de esta a la tribu Detarieae s.l. por la presencia de canales axiales difusos. Anadenantheroxylon villaurquisense, afín al género Anadenanthera Speg. se caracteriza por poseer vasos relativamente pequeños, parénquima axial bandeado y radios homogéneos generalmente uniseriados (Brea et al., 2001). Piptadenioxylon paraexcelsa posee porosidad difusa a semi-circular, vasos predominantemente solitarios y radios homocelulares 1-2 seriados.
Pons (1983) describe, del departamento de Huila de Colombia, de edad miocena, la morfoespecie "Leguminoxylon dindense" no efectivamente publicada (Art. 29 ICNB, Mcneill et al., 2006). Esta última posee porosidad difusa, parénquima vasicéntrico a bandeado y radios 1-3 seriados.
Del Mioceno de Perú, Pons & De Franceschi (2007) describen brevemente dos ejemplares. Uno de ellos con afinidad a Andira o Hymenolobium con abundante parénquima axial, radios uni-triseriado y parénquima cristalífero. El otro ejemplar, afín a la tribu Ingae de las Mimosoideae, posee parénquima aliforme confluente, cristales y radios generalmente uniseriados. Estos autores mencionan también dos maderas afines a la subfamilia Mimosoideae pero no realizan descripciones de las mismas (Table 1, Pons & De Franceschi, 2007).
Provenientes de sedimentos del Mioceno Superior - Plioceno (Formación El Cajón), Fernández & Bravo (1985) describen brevemente un espécimen afín a Acacioxylon de la provincia de Catamarca (Argentina). En la provincia de La Rioja (Argentina) aflora la Formación Toro Negro (Mioceno Superior - Plioceno), la cual posee una rica asociación de fósiles entre los que se encuentra la madera Gleditsioxylon riojana Martínez & Rodríguez Brizuela (2011). Esta morfoespecie presenta caracteres diagnósticos tales como engrosamientos helicoidales en vasos y radios altos que la alían con las Caesalpinoideae.
Los registros del Plioceno son aún más numerosos, y es de este período donde provienen los primeros hallazgos de maderas fósiles del norte de Argentina, con el trabajo de O 'Donell (1938) quien describe muy brevemente maderas fósiles de dicotiledóneas. Posteriormente, Menéndez (1962) estudia ejemplares similares de la misma zona y describe la morfoespecie Acacioxylon odonellii Menéndez. Müller Stoll & Mädel (1967) combinan esta morfoespecie a Paracacioxylon Müller Stoll & Mädel, creado por ellos para estaúnica morfoespecie, debido a que Acacioxylon, según la diagnosis de Felix & Nathorst (1899), presenta fibras no septadas, mientras que los ejemplares de Menéndez (1962) poseen fibras septadas. Lutz (1987) describe varios ejemplares de P. odonellii provenientes de distintas localidades donde aflora la Formación Andalhuala (Plioceno). Paracacioxylon odonnellii (Menéndez) Müller Stoll & Mädel se caracteriza por tener los anillos marcados, vasos generalmente solitarios, parénquima axial vasicéntrico escaso, radios normalmente 5 seriados y fibras septadas. De la Formación Andalhuala también proceden los leños asignados a Menendoxylon piptadiensis Lutz (1987). Esta madera fósil posee porosidad difusa a circular, radios uniseriados bajos y parénquima axial vasicéntrico, correspondiéndose con la subfamilia Mimosoideae y afín al género actual Piptadenia Benth.
En sedimentos cenozoicos de la República Oriental del Uruguay, probablemente de la Formación Salto (Plioceno), fue descripta Caesalpinium nathorstii Schuster (1910), que posteriormente Edwards (1931) combina a Caesalpinioxylon nathorstii (Schuster) Edwards y luego Müller Stoll & Mädel (1967) a Ingoxylon nathorstii (Schuster) Müller Stoll & Mädel. Finalmente, Gros (1992) combina la morfoespecie a Paralbizioxylon nathorstii (Schuster) Gros por problemas nomenclaturales con el morfogénero Ingoxylon Müller Stoll & Mädel. Esta madera posee vasos solitarios o en series radiales hasta 3 vasos, radios uniseriados a veces biseriados y fibras septadas.
El registro más septentrional de maderas fósiles de Leguminosae de Sudamérica es del Plioceno del departamento de Sucre en Colombia (Boureau & Salard, 1962; Pons, 1980). Pons (1980) describe el nuevo morfogénero Crudioxylon Pons caracterizado por el parénquima bandeado de poco espesor, poca densidad de vasos y radios 1-3 seriados y al cual le asigna la especie Crudioxylon pinalense Pons, y otro ejemplar de una localidad cercana y sedimentos un poco más antiguos (Formación El Descanso) lo asigna a Crudioxylon sp. Previamente Boureau & Salard (1962) describieron unas maderas fósiles asignándolas a Ficoxylon cretaceum Schenk. Pons (1980) sinonimiza esta última morfoespecie con Crudioxylon pinalense.
Las primeras maderas de la Mesopotamia argentina asignadas a Leguminosae fueron descriptas por Lutz (1979) y proceden de las Formaciones Salto Chico e Ituzaingó (Plioceno-Pleistoceno), quien menciona tres morfoespecies asignadas al género Menendoxylon Lutz: Menendoxylon vasallensis Lutz, Menendoxylon mesopotamiensis Lutz y Menendoxylon areniensis Lutz. Este morfogénero se caracteriza por tener porosidad difusa, vasos solitarios o en series radiales cortas y radios 1-4 seriados (Lutz, 1979). Lutz (1991) describe otro ejemplar de la Formación Ituzaingó y lo asigna a Mimosoxylon sp. El mismo posee vasos grandes, anillos poco marcados y radios normalmente 3-5 seriados. Posteriormente, Franco & Brea (2010) definen al morfogénero Microlobiusxylon Franco & Brea de la misma Formación, con afinidad al género actual Microlobius C. Presl. y caracterizado por poseer parénquima axial muy abundante, parénquima cristalífero y radios normalmente 1-2 seriados. Recientemente, Franco (2010) describe una nueva madera afín al género Prosopis L. proveniente de la Formación Ituzaingó, con vasos pequeños a medianos, y vasos en series radiales o agrupados.
Mussa (1959) describe del Neógeno el morfogénero Zollernioxylon Mussa y le asigna tres nuevas morfoespecies. Zollernioxlyon se caracteriza por poseer parénquima bandeado, estratificación en parénquima y radios, parénquima cristalífero y radios 1-3 seriados. Las tres morfoespecies descriptas por la autora presentan diferencias cuantitativas entre ellas. Recientemente de la mismaárea, Kloster et al. (2010) describen una madera con afinidad a las Caesalpinoideae procedente de la Formación Solimões caracterizada por poseer vasos mayoritariamente solitarios, parénquima unilateral a confluente y radios 1-2 seriados.
Del terciario de Colombia, Schönfeld (1947) describe Leguminoxylon grossei Schönfeld caracterizada por parénquima paratraqueal bandeado, radios 2-3 seriados. Sin embargo, Müller Stoll & Mädel (1967) dudan sobre la afinidad de esta última a la familia Leguminosae. En años recientes, Selmeir (2004) describe Crudioxylon brasilense Selmeier recolectada en sedimentos cenozoicos del río Parnaiba en el estado de Maranho en Brasil, sin establecer una mayor precisión en cuanto a la edad. Esta última posee un característico bandeado parenquimático, vasos solitarios o en series radiales cortas y radios homo o heterocelulares uniseriados. Silva Curvello (1955) describe de Minas Gerais en Brasil una madera de edad terciaria incierta, Leguminoxylon piptadenoides Silva Curvello, con parénquima vasicéntrico y radios biseriados heterogéneos. Sin embargo, según la autora, la presencia de placas escalariformes pone en duda la afinidad de estos ejemplares a las Leguminosae. Posteriormente Leguminoxylon piptadenoides fue combinada por Müller Stoll & Mädel (1967) a Mimosoxylon piptadenoides (Silva Curvello) Müller Stoll & Mädel.
Felix (1882) describe por primera vez una madera afín a las Leguminosae para Sudamérica, y la nombra Taenioxylon ingaeforme proveniente de una localidad desconocida de Brasil o Uruguay de edad terciaria incierta. Se caracteriza por presentar parénquima aliforme a confluente, parénquima terminal radios homogeneos o heterocelulares, radios 1-5 seriados (Müller Stoll & Mädel 1967). Posteriormente Müller Stoll & Mädel (1967) la combinan al morfogénero Tetrapleuroxylon.
Salard (1963) cita la presencia de Tetrapleuroxylon ersanense (Boureau) Müller Stoll & Mädel, como Leguminoxylon ersanense Boreau de sedimentos presuntamente terciraios de Perú. Esta madera posee porosidad semianular, radios heterogéneos 2-3 seriados, y parénquima axial aliforme y terminal.
Del Pleistoceno, varios de los registros de maderas de Leguminosae provienen de la Formación El Palmar donde Brea (1999) cita la presencia de Menendoxylon piptadiensis y luego Zucol et al. (2005) la presencia de Menendoxylon areniensis y Menendoxylon mesopotamiensis. Posteriormente Brea et al. (2010a) describen 4 nuevas morfoespecies. La primera Amburanoxylon tortorellii Brea, Zucol & Patterer afín al género actual Amburana Schwacke & Taub. caracterizada por la presencia de vasos principalmente solitarios, múltiples radiales de 2-5 elementos y agrupados, menos de 5 vasos por mm2, radios homocelulares y parénquima axial vasicéntrico y confluente. La segunda Holocalyxlyon cozzoii Brea, Zucol & Patterer afín a Holocalyx Micheli. presenta vasos solitarios, múltiples radiales cortos y agrupados, radios homocelulares principalmente biseriados y estratificados, parénquima axial apotraqueal en bandas marginales, aliforme y confluente y cristales prismáticos en células del parénquima axial. La tercera es Prosopisinoxylon castroae Brea, Zucol & Patterer que se diferencia de P. anciborae (Martínez, 2010) por la presencia de porosidad difusa, la densidad de vasos, el ancho de los radios y el arreglo del parénquima axial. Por último, la cuarta morfoespecie es Mimosoxylon caccavariae Brea, Zucol & Patterer que se caracteriza por la presencia de vasos solitarios, múltiples radiales de 2-4 elementos con tendencia a un patrón dendrítico, numerosos y con radios 1-4 seriados. Del Pleistoceno de São Paulo, Brasil, Suguio (1971) menciona la presencia de tres maderas con afinidad a los géneros actuales Myrocarpus, Piptadenia y Centrolobium. Luego Suguio & Mussa (1978) describen de una localidad cercana a la de Suguio (1971) dos nuevas morfoespecies: Piptadenioxylon chimeloi Suguio & Mussa de vasos pequeños a medianos, radios estratificados y 2-3 seriados y Myrocarpoxylon sanpaulense Suguio & Mussa de vasos pequeños a medianos y estratificación en radios y fibras.
Comentarios sobre la preservación de las maderas fósiles de Leguminosae
La mayoría de los fósiles se encuentran en buen estado de preservación anatómica, por lo que es posible establecer una afinidad a nivel morfoespecífico en prácticamente la totalidad de los registros. Solo se registran dos casos de biodeterioro en maderas fósiles afines a Leguminosae. El primer caso fue documentado para la Formación Salicas, donde Tauber & Mazzoni (2003) describen galerías causadas por larvas de coleópteros de la familia Buprestidae en maderas de supuesta afinidad a Prosopis. El otro caso es en Gleditsioxylon riojana (Formación Toro Negro), en la cual se observan hifas y probables cuerpos fructíferos dentro de los vasos (Martínez & Rodríguez Brizuela, 2011).
CONCLUSIONES
El registro de maderas fósiles de Leguminosae resulta ser tanto extenso como así también el más diverso de Sudamérica, con 56 registros contabilizados (Tabla 1; Fig. 1). Otras familias con abundantes xilotipos en América del Sur son las Nothofagaceae y las Anacardiaceae (Poole, 2002; Herbst et al., 2007; Franco, 2009; Pons & de Franceschi, 2007; Pujana, 2009a, 2009b; Martínez & Pujana, 2010). Su registro temporal es también extenso abarcando desde el Paleoceno (Daniano) hasta el Pleistoceno, no obstante la mayor concentración se halló en sedimentos neógenos.
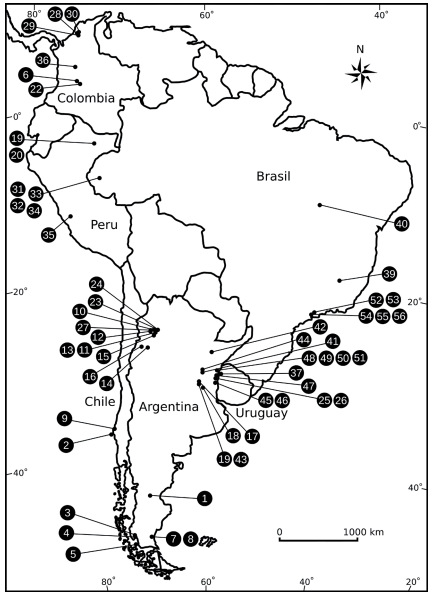
Fig. 1. Mapa de Sudamérica con la ubicación de las localidades fosilíferas. En los xilotipos que se encontraron en más de una localidad fosilífera de una misma unidad estratigráfica, figura la localidad del holotipo. El registro no. 38, no incluido en la figura, proviene de una localidad desconocida de Uruguay o Brasil.

Fig. 2. Anatomía de algunos xilotipos de Leguminosae. A, sección transversal de Doroteoxylon vicente-perezii, Caesalpinoideae. Escala: 50 μm. B, sección transversal del "xilotipo 3" de Pujana (2008), Papilionoideae. Escala: 500 μm. C, sección longitudinal tangencial del "xilotipo 3" de Pujana (2008), Papilionoideae. Escala: 250 μm. D. sección transversal de Gleditsioxylon riojana. Escala: 500 μm. E, Sección longitudinal tangencial de una Mimosoideae de la Formación Chiquimil (Martínez, en prep.). Escala: 20 μm. F, imagen de microscopio electrónico de barrido de Gleditsioxylon riojana. Escala: 100 μm. G, sección transversal de Amburanoxylon tortorellii, Papilionoideae. Escala: 200 μm. H, sección transversal de Paracacioxylon odonnelli, Mimosoideae. Escala: 500μm. I, sección longitudinal tangencial de Holocalyxylon cozzoi, Papilionoideae. Escala: 200 μm.
Dentro de Argentina se encuentra el 55 % de los registros de Sudamérica y las localidades fosilíferas se pueden agrupar en 3 zonas bien delimitadas, centro-noroeste (Neógeno), Mesopotamia (Neógeno) y Patagonia (Paleógeno y Neógeno). Cada xilotipo descripto se encuentra únicamente en una unidad estratigráfica, a excepción de Doroteoxylon vicente-perezii, el "xilotipo 3" y algunas especies de Menendoxylon que fueron encontrados en formaciones cercanas, tanto geográfica como temporalmente. Esta particularidad sugeriría una gran diversidad de Leguminosae en el pasado de Sudamérica como también se observa en el presente.
Si bien es frecuente que la mala preservación en maderas fósiles de angiospermas en general dificulte la asignación a nivel específico, las Leguminosae estudiadas para Sudamérica pudieron ser casi en su totalidad asignadas a un género o especie actual afín. Sin embargo, algunas (i.e. Doroteoxylon vicente-perezii) no poseen una afinidad genérica actual precisa pero este se debe a otros factores y no a la preservación. Los factores podrían ser: extinción del linaje, su evolución anatómica o la ausencia de la descripción anatómica de las maderas actuales de sus descendientes.
En base a los registros consultados se observa que existen trabajos los cuales no se limitan solo a la descripción taxonómica, sino que también aportan a partir de los estudios anatómicos, inferencias paleoambientales y paleobiogreográficas. Por ejemplo, es frecuente que la anatomía presente estructuras adaptadas a condiciones xerofíticas, es decir, estructuras anatómicas especializadas para tolerar el estrés hídrico como por ejemplo la presencia de vasos de dos tamaños distintos (i.e. Brea et al., 2010a; Franco, 2010; Martínez, 2010; Pujana, 2010) mientras que también la presencia de determinados taxones indica las condiciones paleoecológicas y/o regiones fitogeográficas donde se habrían desarrollado las plantas en el pasado (i.e. Franco & Brea, 2010). Asimismo, algunos taxones pueden contribuir a la historia biogeográfica de determinado grupo (i.e. Pujana, 2008; Martínez & Rodríguez Brizuela, 2011).
Los 56 registros de maderas fósiles de Leguminosae de Sudamérica proceden en gran parte de trabajos recientes, prácticamente la mitad es de los últimos 10 años (i.e. Pujana, 2009a; Franco & Brea, 2010; Brea et al., 2010a; Martínez, 2010), lo que sugiere que las investigaciones sobre esta temática se irán incrementando en los años siguientes.
AGRADECIMIENTOS
Este trabajo surgió gracias a la Dra. Maria A. Castro que propuso realizar esta revisión para la V Conferencia Internacional de Leguminosas, que se llevó a cabo en Buenos Aires, Argentina en agosto de 2010. Los autores agradecen a Luis Palazzesi y a Sergio Archangelsky por su colaboración con la bibliografía.
BIBLIOGRAFÍA
1.Boureau, E. & M. Salard. 1962. Sur un bois fossile du département de Bolivar (Colombie). Bol. Geo. Univ. Ind. Santander (Bucaramanga) 11: 35-44. [ Links ]
2.Brea M. 1999. Leños fósiles de Anacardiaceae y Mimosaceae de la Formación El Palmar (Pleistoceno superior), departamento de Concordia, provincia de Entre Ríos, Argentina. Ameghiniana 36: 63-69. [ Links ]
3. Brea M., P. G. Aceñolaza & A.F. Zucol. 2001. Estudio paleoxilológico en la Formación Paraná, Entre Ríos, Argentina. XIº Simposio Argentino de Paleobotánica y Palinología, Publicación Especial no. 8, pp. 7-17, Asociación Paleontológica Argentina. [ Links ]
4. Brea M, A.B. Zamuner, S.D. Matheos, A. Iglesias & A.F. Zucol. 2008. Fossil wood of the Mimosoideae from the early Paleocene of Patagonia, Argentina. Alcheringa 32: 427-441. [ Links ]
5. Brea, M., A.F. Zucol & N. Patterer. 2010a. Fossil woods from late Pleistocene sediments from El Palmar Formation, Uruguay Basin, Eastern Argentina. Rev. Palaeobot. Palyno. 163: 35-51. [ Links ]
6. Brea, M., A.F. Zucol & M.S. Bargo. 2010b. Estudios paleobotánicos en la Formación Santa Cruz (Mioceno), Patagonia, Argentina. Xº Congreso Argentino de Paleontología y Bioestratigrafía y VIIº Congreso Latinoamericano de Paleontología, Resúmenes, p. 140. [ Links ]
7. Brea, M., A.F. Zucol & A. Iglesias. En prensa. Fossil plants studies from late early Miocene in Santa Cruz Formation: Paleoecology and paleoclimatology at the passive margin of Patagonia, Argentina. En: S.F. Vizcaíno, R.F. Kay & M.S. Bargo (Eds.), Paleobiology in Patagonia. Reconstructing a High-Latitude Paleocommunity in the Early Miocene Climatic Optimum. Cambridge University Press. [ Links ]
8. Brea, M., M.J. Franco & A.I. Lutz. Enviado 1. Redescription and re-assignation of Entrerrioxylon victoriensis from the Middle Miocene, Paraná Formation, South America. Rev. Palaeobot. Palyno. [ Links ]
9. Ciccioli, P.L., C.O. Limarino & S.A. Marenssi. 2005. Nuevas edades radimétricas para la Formación Toro Negro en la Sierra de los Colorados, Sierras Pampeanas Noroccidentales, provincia de La Rioja. Rev. Asoc. Geol. Arg. 60: 251-254. [ Links ]
10. Edwards, W.N. 1931. Dicotyledones (Ligna). Fossilium Catalogus, II, Plantae 17. Berlin, 96 pp. [ Links ]
11. Felix, J. 1882. Studien über fossile Hölzer. Leipzig, 84 pp. [ Links ]
12. Felix, J. & A. Nathorst. 1899. Versteinerungen aus dem mexicanischen Staat Oaxaca 3. Fossile Hölzer von Tlacolula. En: J. Felix & H. Lenk (Eds.), Beiträge zur Geologie und Paläontologia der Republik Mexiko, 2: 46-51, Leipzig. [ Links ]
13. Fernández, R.I. & E.V. Bravo. 1985. La presencia del género Acacioxylon (Leguminosae) en sedimentos terciarios del área de los Nacimientos de Abajo (departamento de Belén - provincia de Catamarca) República Argentina. Vº Simposio Argentino de Paleobotanica y Palinologia, San Miguel de Tucumán, p. 11. [ Links ]
14. Franco, M.J. 2009. Leños fósiles de Anacardiaceae en la Formación Ituzaingó (Plioceno-Pleistoceno), Toma Vieja, Paraná, Entre Ríos, Argentina. Ameghiniana 46: 587-604. [ Links ]
15. Franco, M.J. 2010. Paleoecología de leños de Anacardiaceae y Fabaceae en la Formación Ituzaingó (Plioceno-Pleistoceno), Entre Ríos, Argentina. Xº Congreso Argentino de Paleontología y Bioestratigrafía y VIIº Congreso Latinoamericano de Paleontología, Resúmenes, pp. 102-103. [ Links ]
16. Franco, M.J. & M. Brea. 2008. Leños fósiles de la Formación Paraná (Mioceno Medio), Toma Vieja, Paraná, Entre Ríos, Argentina: registro de bosques estacionales mixtos. Ameghiniana 45: 699-718. [ Links ]
17. Franco M.J. & M. Brea. 2010. Microlobiusxylon paranaensis gen. et sp. nov. (Fabaceae, Mimosoideae) from the Pliocene-Pleistocene of Ituzaingó Formation, Paraná Basin, Argentina. Rev. Bras. Paleont. 13: 103-114. [ Links ]
18. Gregory M., I. Poole & E.A. Wheeler. 2009. Fossil dicot wood names. IAWA Supplement 6. IAWA, 220 pp. [ Links ]
19. Gros J.P. 1992. A synopsis of the fossil record of Mimosoid legume wood. En: P. Herendeen & D.L. Dilcher (Eds.), Advances in legume systematics, 4. The fossil record, pp. 69-83, The Kew Royal Botanic Gardens. [ Links ]
20. Herbst, R., M. Brea, A. Crisafulli, S. Gnaedinger, A.I. Lutz, & L.C.A. Martínez. 2007. La paleoxilología en la Argentina. Historia y desarrollo. En: Publicación Especial no. 11, Ameghiniana 50º aniversario, pp. 57-71, Asociación Paleontológica Argentina. [ Links ]
21. Herendeen, P.S. 1992. The fossil history of the Leguminosae from the Eocene of southeastern North America. En: P. Herendeen & D.L. Dilcher (Eds.), Advances in legume systematics, 4. The fossil record, pp. 85-160, The Kew Royal Botanic Gardens. [ Links ]
22. Inside Wood. 2004. http://insidewood.lib.ncsu.edu/search [acceso abril de 2011]. [ Links ]
23. Kloster, A., S. Gnaedinger & K. Adami Rodrigues. 2010. Xilotafoflora da Formação Solimões, Neógeno, Alto Juruá, Acre, Amazônia Ocidental, Brasil. Xº Congreso Argentino de Paleontología y Bioestratigrafía y VIIº Congreso Latinoamericano de Paleontología, Resúmenes, p. 175. [ Links ]
24. Lewis, G., B. Schrire, B. Mackinder & M. Lock. 2005. Legumes of the World. Royal Botanic Garden, Kew, 592 pp. [ Links ]
25. Lutz, A.I. 1979. Maderas de angiospermas (Anacardiaceae y Leguminosae) del Plioceno de la provincia de Entre Ríos, Argentina. FACENA 3: 39-63. [ Links ]
26. Lutz A.I. 1981. Entrerrioxylon victoriensis nov. gen. et sp. (Leguminosae) del Mioceno Superior (Fm. Paraná) de la provincia de Entre Ríos, Argentina. FACENA, 4: 21-29. [ Links ]
27. Lutz A.I. 1987. Estudio anatómico de maderas terciarias del valle de Santa María (Catamarca-Tucuman), Argentina. FACENA 7: 125-143. [ Links ]
28. Lutz, A.I. 1991. Descripción anatómica de Mimosoxylon sp. del Plioceno del Plioceno (Formación Ituzaingó) de la provincia de Corrientes, Argentina. Rev. Asoc. Cs. Nat. Litoral 22: 3-10. [ Links ]
29. Martínez, L.C.A. 2007. Maderas de Prosopis L. del Mioceno del Noroeste de Argentina. 31º Jornadas Argentinas de Botánica. Resúmenes Boletín de la Sociedad Argentina de Botánica 42 Suplemento, p. 119-120. [ Links ]
30. Martínez, L.C.A. 2010. Prosopisinoxylon anciborae nov. gen. et sp. (Leguminosae, Mimosoideae) from the Late Miocene Chiquimil Formation (Santa María Group), Catamarca, Argentina. Rev. Palaebot. Palyno. 158: 262-271. [ Links ]
31. Martínez, L.C.A. & A.I. Lutz. 2005. Primera cita de una leguminosa fósil, Formación Las Arcas, Grupo Santa María, Catamarca, Argentina. Ameghiniana 42 Suplemento: 34R-35R. [ Links ]
32. Martínez, L.C.A. & A.I. Lutz. 2006. Un nuevo género de Fabaceae fósil de la Formación Chiquimil (Mioceno Superior), Catamarca, Argentina. IXº Congreso Argentino de Paleontología y Bioestratigrafía, Resúmenes p. 44. [ Links ]
33. Martínez, L.C.A. & R.R. Pujana. 2010. Sobre la presencia de Resinaxylon schinusoides Pujana en la Formación San Julián (Oligoceno), Santa Cruz, Patagonia Argentina. Ameghiniana 47: 535-539. [ Links ]
34. Martínez, L.C.A. & R. Rodriguez Brizuela. 2011. Gleditsioxylon riojana nov. sp. en el Neógeno de Argentina. Sus relaciones paleoambientales y biogeográficas. Geobios 44: 461-472. [ Links ]
35. Mcneill, J., F.R. Barrie, H.M. Burdet, V. Demoulin, D.L. Hawksworth, K. Marhold, D.H. Nicolson, J. Prado, P.C. Silva, J.E. Skog, J.H. Wiersema & N.J. Turland. 2006. International code of botanical nomenclature (Vienna Code) adopted by the 17th International Botanical Congress, Vienna, Austria, July 2005 (versión electrónica). International Association for Plant Taxonomy, acceso 04-2011. [ Links ]
36. Menéndez, C.A. 1962. Leño petrificado de una leguminosa del Terciario de Tiopunco, provincia de Tucumán. Ameghiniana 2: 121-126. [ Links ]
37. Müller Stoll W.R. & E. Mädel. 1967. Die fossilen Leguminosen-Hölzer. Palaeontographica B 119: 95-174. [ Links ]
38. Mussa D. 1959. Contribuiçao à paleoanatomia vegetal. II. Madeiras fósseis do territorio do Acre (Alto Juruá), Brasil. Bol. Div. Geo. Mineral. 195: 1-54. [ Links ]
39. Nishida M. 1984 The anatomy and affinities of the petrified plants from the Tertiary of Chile. III. Petrified woods from Mocha island, Central Chile. En: M. Nishida (ed.), Contributions to the botany in the Andes I, Vol I., pp. 96-110, Academia Scientific Book Inc. [ Links ]
40. Nishida M., H. Nishida & T. Ohsawa. 1989. Comparison of the petrified woods from the Cretaceous and Tertiary of Antarctica and Patagonia. NIPR Symposium on Polar Biolology, Proceedings 2: 198-212. [ Links ]
41. O'Donell, C. 1938. Troncos y ramas fósiles de dicotiledóneas en el Araucariano de Tiopunco (Dto. Tafí, Prov. Tucumán). Cuad. Mineral. Geol. UNT 1: 26-29. [ Links ]
42. Pons, D. 1980. Les bois fossile du Tertiaire Superieur de la region de Toluviejo-Corozal (Dep. de Sucre, Colombie). Comptes Rendus 105º Congrès national des Sociétés savantes, pp. 163-182. [ Links ]
43. Pons, D. 1983. Contribution à l'étude paléobotanique du Mésozoïque et du Cénozoïque de Colombie. Tesis doctoral, Université Pierre et Marie Curie, Paris, 655 pp. [ Links ]
44. Pons, D. & D. De Franceschi. 2007. Neogene woods from western Peruvian Amazon and palaeoenvironmental interpretation. B. Geosci. 82: 343-354. [ Links ]
45. Poole, I. 2002. Systematics of Cretaceous and Tertiary Nothofagoxylon: implications for southern hemisphere biogeography and evolution of the Nothofagaceae. Aust. Syst. Bot. 15: 247-276. [ Links ]
46. Pujana, R.R. 2008. Estudio paleoxilológico del Paleógeno de Patagonia austral (Formaciones Río Leona, Río Guillermo y Río Turbio) y Antártida (Formación La Meseta). Tesis doctoral Universidad de Buenos Aires, Buenos Aires, 182 pp. [ Links ]
47. Pujana, R.R. 2009a. Fossil woods from the Oligocene of southwestern Patagonia (Río Leona Formation). Atherospermataceae, Myrtaceae, Leguminosae and Anacardiaceae. Ameghiniana 46: 523-535. [ Links ]
48. Pujana, R.R. 2009b. Fossil woods from the Oligocene of southwestern Patagonia (Río Leona Formation). Rosaceae and Nothofagaceae. Ameghinana 46: 621-636. [ Links ]
49. Pujana, R.R. 2010. Una nueva Mimosoideae (Leguminosae) de la Formación Salicas (Mioceno), provincia de La Rioja, Argentina. Xº Congreso Argentino de Paleontología y Bioestratigrafía y VIIº Congreso Latinoamericano de Paleontología, Resúmenes p. 203. [ Links ]
50. Raven, P.H. & R.M. Polhill. 1981. Biogeography of Leguminosae. En: R.M. Polhill & P.H. Raven (Eds.), Advances in Legume Systematics, Part 1, pp. 27-34, Royal Botanic Gardens, Kew. [ Links ]
51. Salard, M. 1963. Sur un bois tertiaire du Pérou. Comptes Rendus 88º Congrès national des Sociétés savantes, pp. 483-496. [ Links ]
52. Schönfled, G. 1947. Hölzer uas dem Tertiär von Kolumbien. Abh. Senckenberg. Naturf. Ges. 475: 1-53. [ Links ]
53. Schöning M. & K. Bandel. 2004. A diverse assemblage of fossil hardwood from the Upper Tertiary (Miocene?) of the Arauco Peninsula, Chile. J. S. Am. Earth Sci. 17: 59-71. [ Links ]
54. Schuster, J. 1910. Ueber Nicolien und Nicolien ähnliche Hölzer. Kungl. Svenska Vet. Handl. 45: 1-18. [ Links ]
55. Selmeier A. 2004. First record of Matisia (Bombacaceae) and Crudia (Caesalpiniaceae) wood from the Tertiary of Rio Paranaiba, Brazil. Zitteliana 44: 113-124. [ Links ]
56. Silva Curvello, W. 1955. Sôbre um vegetal do Linhito de Fonseca, Minas Gerais. An. Acad. Bras. Ciênc. 27: 293-296. [ Links ]
57. Suguio K. 1971. Estudo dos troncos de árvores linhitificados dos aluvioés antigos do Rio Pinheiros (São Paulo). Anais XXVº Congreso Brasileiro de Geología Vol. 1., pp. 63-69. [ Links ]
58. Suguio K. & D. Mussa. 1978. Madeiras fosseis dos aluvioes antigos do Rio Tiete, Sao Paulo. Bol. IG- Univ. Sao Paulo, Inst. Geocienc. 9: 25-45. [ Links ]
59. Tauber, A.A. & A.F. Mazzoni. 2003. Trazas de insectos xilófagos de la Formación Salicas (Mioceno tardío) de la provincia de Catamarca, Argentina. Ameghiniana 40 Suplemento: 94R. [ Links ]
60. Terada K., T. O. Asakawa & H. Nishida. 2006. Fossil wood assemblage from Cerro Dorotea, Última Esperanza, Magallanes (XII) region, Chile. En: H. Nishida (ed.), Post-Cretaceous floristic changes in Southern Patagonia, Chile. Faculty of Science and Engineering, Chuo University, pp. 67-90. [ Links ]
61. Wheeler, E.A. & P. Bass. 1992. Fossil wood of the Leguminosae: a case study in xylem evolution and ecological anatomy. En: P. Herendeen & D.L. Dilcher (Eds.), Advances in legume systematics, 4. The fossil record, pp. 281-301. [ Links ]
62. Zucol, A.F., M. Brea & A. Scopel. 2005. First record of fossil wood and phytolith assemblages of the Late Pleistocene in El Palmar National Park (Argentina). J.S. Am. Earth Sci. 20: 33-43. [ Links ]
Recibido: 13-VI-2011
Aceptado: 4-XI-2011

