










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
versión On-line ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.16 no.1 Ciudad Autónoma de Buenos Aires jun. 2014
PALEONTOLOGÍA
Estudios palinológicos del Grupo Baqueró (Cretácico Inferior), provincia de Santa Cruz, Argentina. X. Polen de Gymnospermae y Apéndice Final
Liliana Villar de Seoane & Sergio Archangelsky
CONICET- División Paleobotánica, Museo Argentino de Ciencias Naturales "B. Rivadavia", Av. Ángel Gallardo 470, (1405) Buenos Aires, Argentina. E-mail: lvillar@macn.gov.ar; sarcang@fibertel.com.ar
Resumen
Con el presente trabajo se completa el estudio palinológico de las gimnospermas halladas en la Formación Anfiteatro de Ticó, que comprende los afloramientos de la localidad homónina, y los de la Estancia Bajo Tigre. Esta formación se encuentra en la base del Grupo Baqueró, perteneciente al Aptiano de la provincia de Santa Cruz. El estudio incluye 9 géneros distribuidos en 11 especies, de los cuales 1 es género nuevo (Ticoites), 3 son nuevas especies (Araucariacites spinosus, Ephedripites acutus y Ticoites gamerroi) y 1 especie se cita por primera vez para el Cretácico Inferior de Argentina (Dilwynites granulatus Harris). Los resultados finales de esta serie confirman la existencia de una gran variedad de polen de gimnospermas, grupo que en la actualidad está casi extinto, debido a los grandes cambios climáticos ocurridos en Patagonia desde el Aptiano hasta nuestros días.
Palabras clave: Gymnospermae; Granos de polen; Formación Anfiteatro de Ticó; Aptiano; Provincia de Santa Cruz; Argentina.
Abstract
Palynological studies from the Baqueró Group (Lower Cretaceous), Santa Cruz Province, Argentina. X. Pollen of Gymnospermae and final Appendix. In the present paper, the palynological study of Gymnosperm pollen from the Anfiteatro de Ticó Formation is completed. The material was collected in Anfiteatro de Ticó and Estancia Bajo Tigre localities from the Santa Cruz Province. The Anfiteatro de Ticó Formation, the basal unit of the Baqueró Group, has an Aptian age. The study includes 9 genera and 11 species. One new genus (Ticoites), and three new species (Araucariacites spinosus, Ephedripites acutus and Ticoites gamerroi) are described. Also, Dilwynites granulatus Harris is cited for the first time in the Lower Cretaceous of Argentina. This paper, the final contribution of the series, indicates the presence of a wide variety of gymnosperms, a group that nowadays is almost extinct in Patagonia due to large recurrent climatic changes that have occurred since the Aptian.
Key words: Gymnospermae; Pollen grains; Anfiteatro de Ticó Formation; Aptian; Santa Cruz Province; Argentina.
INTRODUCCIÓN
A lo largo de las últimas décadas, se han publicado diversos trabajos sobre la palinología de la Formación Anfiteatro de Ticó (Archangelsky, 1994; Archangelsky & Gamerro, 1965, 1966 a, b, c, 1967; Archangelsky & Villar de Seoane, 1989, 1990, 1991, 1992, 1994, 1996, 1998, 2005; Del Fueyo & Archangelsky, 2005; Limarino et al., 2012; Pérez Loinaze et al., 2013), Esta formación es la unidad basal del Grupo Baqueró, y es la más rica y diversa tanto mega como microscópicamente en relación a Bajo Tigre y Punta del Barco, las otras dos formaciones del Grupo.
Si bien, a nivel polínico las pteridófitas presentan mayor diversidad específica, las gimnospermas son un componente muy abundante dentro de las comunidades vegetales de la Formación Anfiteatro de Ticó, en especial las Coniferales. En este orden se destacan las Araucariaceae, Podocarpaceae y Cheirolepidiaceae y en menor medida las Taxodioideae.
Las Cheirolepidiaceae se extinguieron en el comienzo del Cenozoico (Pocock et al., 1990). Respecto a los grupos actuales, ya no existen Taxodioideae en Patagonia y la mayoría de los taxones de las Araucariaceae y Podocarpaceae se hallan en Australia y Nueva Zelanda (Krüssmann, 1985), quedando sólo algunas especies nativas en el sur argentino (e.g.: Araucaria araucana (Mol.) K. Koch, Podocarpus nubigenus Lindley, y Saxegothaea conspicua Lindley).
En la Formación Anfiteatro de Ticó, las Podocarpaceae están representadas fundamentalmente por granos bi y trisacados. Los granos bisacados pertenecen a los géneros Gamerroites Archangelsky y Podocarpidites Cookson, éste último muy abundante y diversificado (Archangelsky & Villar de Seoane, 2005). Entre los granos trisacados, se destacan los géneros Microcachryidites Cookson ex Couper y Trisaccites (Cookson) Menéndez y Callialasporites trilobatus (Balme) Sukh Dev.
Las Araucariaceae están representadas por Araucariacites Cookson ex Couper, Dilwynites Harris, Cyclusphaera Elsik y Balmeiopsis Archangelsky, siendo los dos últimos dominantes en los diferentes niveles analizados. Taylor et al. (1989) realizaron un estudio detallado de la exina de Cyclusphaera psilata Volkheimer & Sepúlveda, y Archangelsky (1994) estudió comparativamente las ultraestructuras de Callialasporites, Balmeiopsis y Araucariacites. Dettmann & Jarzen (2000) analizaron la ultraestructura de Dilwynites y la compararon con la del género actual Wollemia. Del Fueyo & Archangelsky (2005) estudiaron la ultraestructura de granos de Cyclusphaera encontrados in situ dentro de un cono microesporangiado.
Las Cycadophyta, si bien presentan una gran diversidad foliar, especialmente las Bennettitales, ello no se corresponde con su escasa representación polínica, con una única especie (Cycadopites nitidus (Balme) de Jersey). En cambio, las Caytoniales (Vitreisporites Leschik) son muy abundantes y de amplia distribución.
Las Gnetales que fueron cosmopolitas durante el Cretácico Medio, aparecen pobremente representadas por dos especies de distribución muy restringida (Ephedripites acutus sp. nov. y Ephedripites sp. A).
En la presente contribución, damos a conocer los taxones gimnospérmicos que completan el conocimiento de este grupo de plantas de la Formación Anfiteatro de Ticó. El estudio incluye 9 géneros, uno de ellos nuevo (Ticoites) y 11 especies, de las cuales 3 son nuevas (Araucariacites spinosus, Ephedripites acutus y Ticoites gamerroi), mientras que otra se cita por primera vez en estratos del Cretácico Inferior de Argentina (Dilwynites granulatus Harris).
MATERIALES Y MÉTODOS
El material estudiado fue colectado en diferentes niveles fosilíferos de la Formación Anfiteatro de Ticó, los mismos proceden de tres afloramientos ubicados en el Anfiteatro de Ticó, y en la Estancia Bajo Tigre. El área de estudio se halla ubicada en la base del Grupo Baqueró, a 48º30'33" S y 69º14'11" O en la provincia de Santa Cruz, Argentina. Su ubicación geográfica y el detalle de los diferentes perfiles pueden ser consultados en Cladera et al. (2002). La Formación Anfiteatro de Ticó está datada isotópicamente en el Aptiano (Corbella, 2001; Limarino et al., 2012; Pérez Loinaze et al., 2013).
El estudio fue realizado con un microscopio Leitz Diaplan de la División Paleobotánica del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" y las coordenadas de los ejemplares corresponden al citado microscopio. Las fotografías fueron obtenidas con una cámara digital Leica DFC 280 y las mediciones fueron efectuadas con el programa Leica QWin.
El material analizado se encuentra en preparados microscópicos que fueron utilizados en contribuciones anteriores (Archangelsky & Villar de Seoane, 1992, 1994, 1996, 1998, 2005) y que se hallan depositados en la Colección Palinológica del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" bajo la sigla BA Pal.
El número que aparece entre paréntesis en el ítem Medidas de cada una de las especies descriptas, corresponde a la media.
PALEONTOLOGÍA SISTEMÁTICA
Turma SACCITES Erdtman 1947
Subturma MONOSACCITES Chitaley emend.
Potonié & Kremp 1954
Género Ticoites nov. gen.
Especie tipo. Ticoites gamerroi nov. sp.
Ticoites gamerroi nov. sp.
(Fig. 1, A-E)
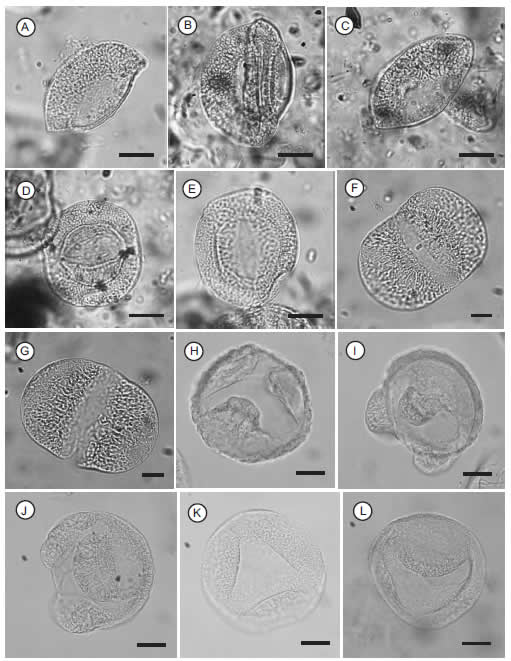
Fig. 1. A-E, Ticoites gamerroi nov. gen. et sp.; F-G, Vitreisporites pallidus (Reissinger) Nilsson 1958; H-I, Microcachryidites castellanosii Menéndez 1968; J, Microcachryidites antarcticus Cookson 1947; K-L, Trisaccites microsaccatum (Couper) Couper 1960.
Escalas gráficas = 10 µm/ Scale bars = 10 µm.
1977. Spec. Indet., Harris, Plate 4, fig. 21.
1983. Monosaccites sp. 2. Kotova, pág. 900, Plate 9, figs. 11 y 12.
2002. Accinctisporites sp., Prámparo & Volkheimer, pág. 399, Figura 3 H.
Derivatio nominis. El epíteto genérico alude a su presencia en la Formación Anfiteatro de Ticó. El nombre específico alude al Lic. Juan Carlos Gamerro, destacado palinólogo argentino quien contribuyó inicialmente con el estudio de la palinología de la Formación Anfiteatro de Ticó.
Diagnosis (genérico-específica). Grano de polen monosacado de contorno circular a levemente elíptico en vista polar y lenticular en vista ecuatorial. Saco continuo de borde liso, insertado Fig. 1. A-E, Ticoites gamerroi nov. gen. et sp.; F-G, Vitreisporites pallidus (Reissinger) Nilsson 1958; H-I, Microcachryidites castellanosii Menéndez 1968; J, Microcachryidites antarcticus Cookson 1947; K-L, Trisaccites microsaccatum (Couper) Couper 1960.
Escalas gráficas = 10 µm/ Scale bars = 10 µm. en la cara distal dejando libre la cara proximal. Saco con retículo uniforme de aréolas poligonales. Colpo de bordes nítidos a difusos (leptoma). Exina tectada y lisa.
Diagnosis. Monosaccate pollen grain with circular to lightly elliptic amb in polar view and lenticular in equatorial view. A continuous saccus with smooth outline only inserted on the distal face. Saccus with polygonal areolae in the reticulum. Colpus with visible to few visible outline (leptoma). Exine tectate and smooth.
Material tipo. Holotipo. N.F. Williamsonia, BA Pal. 5010, coord. 36,7/ 102,3.
Paratipo. Estancia Bajo Tigre, N.F. Pique Carbón, BA Pal. 5052, coord. 49,5/105,5.
Distribución. Fm. Anfiteatro de Ticó, N.F. Williamsonia, BA Pal. 5010; Estancia Bajo Tigre, N.F. Pique Carbón, BA Pal. 5050, 5051, 5052, 5053, 5064, 5065, 5066 y 5067.
Procedencia geográfica. Estancia Bajo Tigre, provincia de Santa Cruz, Argentina.
Procedencia estratigráfica. Grupo Baqueró, Formación Anfiteatro de Ticó, Aptiano.
Descripción. Grano de polen monosacado de contorno circular a levemente elíptico en vista polar (Fig. 1A-C) y lenticular en vista ecuatorial (Fig.1D-E). Saco continuo de borde liso, insertado en la cara distal rodeando un leptoma colpado (Fig. 1D-E), dejando libre la cara proximal. El saco presenta un retículo uniforme con aréolas poligonales, las de mayor diámetro ubicadas cercanas al cuerpo y disminuyendo su tamaño hacia el borde (Fig. 1A). El colpo presenta bordes nítidos a difusos, en este último caso se asemeja a un leptoma (Fig. 1D). Exina tectada, externamente lisa (Fig. 1A).
Medidas. (52 ejemplares) Ancho total: 29,4 (37,4) 44,8 µm; Largo total: 28,7 (36,3) 39,2 µm; Alto total: 16,8 (21,8) 28,7 µm; Ancho del cuerpo: 16,8 (20,5) 26,1 µm; Largo del cuerpo: 20,6 (26,3) 35,3 µm; Alto del cuerpo: 9,8 (14,5) 20,6 µm; Ancho mayor del saco: 5,6 (10,6) 15,4 µm; Ancho menor del saco: 5,6 (9,3) 11,8 µm; Diámetro de las areolas: 1,4 (2,3) 2,9; Largo del colpo: 10,3 (18,4) 20,6 µm; Espesor exina: 1,1 (1,6) 2,2 µm.
Comentarios. El polen monosacado no es un tipo frecuente en las asociaciones palinológicas, tanto en las actuales como en las fósiles, menos aún en las cretácicas, aunque a veces dominan en algunas asociaciones del paleozoico superior. En la Tabla 1 se presentan los géneros de polen monosacado fósil reconocidos hasta el presente para el Mesozoico y se definen sus características esenciales. En su comparación con Ticoites se observan como diferencias importantes, la exina lisa y el tamaño del grano sensiblemente menor a los otros géneros (Accinctisporites, Succinctisporites, Patinasporites, Walchiites). A ello se suman las diferencias cronológicas (Triásico Superior y Jurásico Inferior) y las procedencias geográficas extragondwánicas (Suiza, Siberia, Alemania) de los otros géneros. Ottone et al. (1992, 2005) describieron un nuevo género de monosacado (Variapollenites) para el Triásico Medio y Superior de Argentina, pero como en los casos anteriores, Ticoites se diferencia por su exina lisa y su tamaño menor.
Este tipo de polen ya había sido ilustrado como 'monosaccate grain' para la Fm. Anfiteatro de Ticó por Archangelsky & Gamerro (1966, Lám. II, fig D). Por otra parte, Prámparo & Volkheimer (2002) citan a Accinctisporites sp. para el Cretácico temprano de la Formación La Amarga (Cuenca Neuquina sudoriental) que por todas sus características pertenece al nuevo género Ticoites y por ello se lo ha incluido en sinonimia. Otra forma sinonimizada es la citada como Monosaccites sp. 2, para la perforación 511-57-2 perteneciente al Jurásico Superior y Cretácico Inferior de la Plataforma Continental de las Islas Malvinas (Kotova, 1983). Por otra parte, también Harris (1977, Lám. 4, fig. 21) ilustró un ejemplar como "spec. indet." para el Cretácico Temprano de la perforación 330 del Deep Sea Drilling Project, ubicada al este de las Islas Malvinas y al sur de la Cuenca Argentina en el océano Atlántico, y que también asignamos a nuestra especie. Tanto el tamaño como los caracteres morfológicos, edad y ubicación geográfica de los anteriores permiten su nueva asignación.
Turma ALETES Ibrahim 1933
Subturma AZONALETES Luber emend. Potonié & Kremp 1954
Género Araucariacites Cookson ex Couper 1953
Especie tipo. Araucariacites australis Cookson 1947
Araucariacites spinosus sp. nov.
(Fig. 2 C-E)
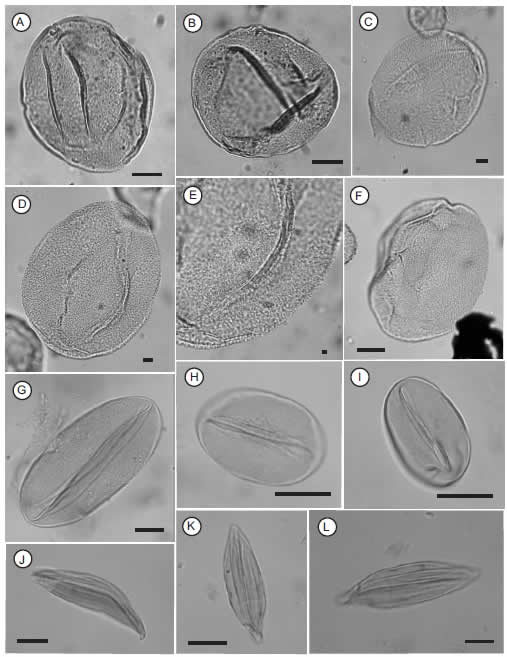
Fig. 2. A-B, Araucariacites australis Cookson 1947; C-E, Araucariacites spinosus sp. nov.; F, Dilwynites granulatus Harris 1965; G-I, Cycadopites nitidus (Balme) de Jersey 1964; J, Ephedripites sp. A; K-L, Ephedripites acutus sp. nov.
Escalas gráficas = 10 µm/ Scale bars = 10 µm.
Derivatio nominis. Del latín spinosus = con espinas.
Diagnosis. Grano de polen inaperturado, de contorno ecuatorial circular a subcircular. Exina gruesa y ornamentada de manera uniforme y densa por diminutas espinas de hasta 1,1 µm de alto.
Diagnosis. Inaperturate pollen grain with a Fig. 2. A-B, Araucariacites australis Cookson 1947; C-E, Araucariacites spinosus sp. nov.; F, Dilwynites granulatus Harris 1965; G-I, Cycadopites nitidus (Balme) de Jersey 1964; J, Ephedripites sp. A; K-L, Ephedripites acutus sp. nov.
Escalas gráficas = 10 µm/ Scale bars = 10 µm.
circular to subcircular equatorial outline. Exine thick bearing compact and minutes spines up to 1.1 µm high.
Material tipo. Holotipo. Estancia Bajo Tigre, N.F. Ginkgoites, BA Pal. 5025, coord. 28/103,2.
Paratipo. Estancia Bajo Tigre, N.F. Ginkgoites, BA Pal. 5025, coord. 32,1/98,2.
Material adicional. Estancia Bajo Tigre, N.F. Ginkgoites, BA Pal. 5025, coord. 48,1/102,8; 45,2/94,8; 44,8/103,2; 42,8/95; 40,8/94; 38,9/95,6; 33,1/110,1.
Procedencia geográfica. Estancia Bajo Tigre, provincia de Santa Cruz, Argentina.
Procedencia estratigráfica. Grupo Baqueró, Formación Anfiteatro de Ticó.
Descripción. Grano de polen grande e inaperturado, de contorno ecuatorial circular a subcircular (Fig. 2D). Exina gruesa y ornamentada por pequeñas espinas de diferentes tamaños (de 0,6 hasta 1,1 µm de alto), densamente distribuidas en toda la superficie (Fig. 2 C-D), y separadas entre sí por una distancia menor a 0,25 µm. La exina está constituida por dos capas del mismo espesor (Fig. 2E).
Medidas. (28 ejemplares): Diámetro ecuatorial: 89,6 (123,2) 148,4 µm; espesor exina: 2,8 (2,9) 5,6 µm; espinas: diámetro: 0,3 µm, altura: 0,6 (0,75) 1,1 µm.
Comparaciones. Araucariacites spinosus sp. nov. se diferencia de la especie tipo, Araucariacites australis Cookson, por poseer un tamaño mayor, una exina más gruesa y con una ornamentación compacta y microespinosa. Araucariacites pergranulatus Volkheimer del Jurásico Medio de la cuenca Neuquina, difiere por su menor tamaño y su ornamentación verrugosa (Volkheimer, 1968).
Afinidad botánica. Los granos de Araucariacites spinosus son semejantes a los granos de polen actuales de la familia Araucariaceae, de amplia distribución en el Hemisferio Sur.
Género Dilwynites Harris 1965
Especie tipo. Dilwynites granulatus Harris 1965
Dilwynites granulatus Harris 1965
(Fig. 2F)
2013. Dilwynites granulatus, Macphail et al., pág. 3, Fig. 2 G-H.
Descripción. Grano de polen inaperturado, de contorno ecuatorial circular a subcircular. Exina delgada, hialina y densamente ornamentada por gránulos. Entre los mismos se observan, especialmente sobre los bordes del grano, escasas clavas y gemas.
Material estudiado. Estancia Bajo Tigre, N.F. Ginkgoites, BA Pal. 5025, coord. 48/110,2; 47,9/106,8; 45,5/99,4; 42,5/100,8; 40,2/108,9; 34,8/102,9; 31/105,9.
Medidas. (13 ejemplares): Diámetro ecuatorial: 45,1 (50) 55 µm; espesor exina: 1,1 µm; diámetro gránulos: 0,3-0,5 µm; diámetro gemas: 0,6 - 1,1 µm; clavas: diámetro: 0,6 µm, altura: 1,1 µm.
Comparaciones. Nuestros ejemplares son muy semejantes a los descriptos por Dettmann y Jarzen (2000) para el Terciario Temprano de Victoria, Australia y para el Cretácico Tardío de las Islas James Ross y Vega de la Península Antártica. También se asemejan a los ejemplares de la Fig. 2 G-H ilustrados por Macphail et al. (2013) para el Eoceno y Mioceno de Australia.
Tabla 1. Comparaciones entre el nuevo género Ticoites y los otros géneros monosacados mesozoicos.

En cambio, difieren de Dilwynites sp. cf. D. tuberculatus Harris descripta para la Formación Ligorio Márquez (Paleoceno Tardío-Eoceno Temprano) de la localidad Río Zeballos, provincia de Santa Cruz, en que ésta posee ornamentación tuberculada y espaciada (Macphail et al., 2013, Fig. 2 A-F). Esta mención sería la primera del género Dilwynites para Argentina y América del Sur.
Afinidad botánica. El polen de Dilwynites granulatus es semejante a los representantes actuales de Wollemia W.G. Jones, K.D. Hill et J.M. Allen (Jones et al., 1995), género monotípico de la familia Araucariaceae que crece al oeste de la ciudad de Sydney, Australia (Dettmann & Jarzen, 2000).
Comentarios. Esta es la primera mención de la especie para el Cretácico Temprano de Argentina, y probablemente la de mayor antigüedad a nivel global.
Turma PLICATES Naumova 1939 emend. Potonié 1960
Subturma COSTATES Potonié 1970
Género Ephedripites Bolkhovitina 1953
ex Potonié 1958
Especie tipo. Ephedripites mediolobatus Bolkhovitina 1953 ex Potonié 1958
Ephedripites acutus sp. nov.
(Fig. 2 K-L)
Derivatio nominis. Del latín acutus = terminado en punta sin disminución gradual.
Diagnosis. Granos de polen perprolados, con extremos aguzados y engrosados. Exina delgada con 4 a 8 carenas longitudinales, ocasionalmente 12.
Diagnosis. Perprolate pollen grain with acute and thickened ends. Exine thin bearing 4-8, occasionally 12 longitudinal ridges.
Material tipo. Holotipo. Fm. Anfiteatro de Ticó, N.F. Williamsonia, BA Pal. 5020, coord. 28/103,6.
Paratipo. Fm. Anfiteatro de Ticó, N.F. Williamsonia, BA Pal. 5021, coord. 32,3/97,3.
Distribución. Fm. Anfiteatro de Ticó, N.F. Williamsonia, BA Pal. 5008, 5020 y 5021; N.F. Basal, BA Pal. 5076 y 5077.
Procedencia geográfica. Provincia de Santa Cruz, Argentina.
Procedencia estratigráfica. Grupo Baqueró, Formación Anfiteatro de Ticó.
Descripción. Los granos de polen son fusiformes y tienen extremos aguzados y engrosados (Fig. 2K). La relación largo/ancho de los granos es de 3,5/1 aproximadamente. La exina posee carenas longitudinales, lisas y de bordes nítidos que culminan en los polos (Fig. 2K-L). El ancho de las carenas es variado, aproximadamente 2,5 µm en los granos de 4 carenas, y de 0,5 µm de espesor en los granos de 8 y ocasionalmente 12 carenas (Fig. 2L).
Medidas. (24 ejemplares) Cuerpo: largo: 31,9 (51,1) 61,6 µm; ancho: 11,0 (14,6) 26,4 µm; espesor exina: 0,6-1,0 µm.
Comparaciones. Ephedripites menendezii Volkheimer descripto para el Jurásico de Neuquén, se diferencia de Ephedripites acutus sp. nov. por tener un cuerpo más angosto, menor número de costillas y poseer exina punteada (Volkheimer, 1972).
Ephedripites sp. A
(Fig. 2J)
Descripción. Granos de polen perprolados, fusiformes, con extremos romos y exina delgada. Carenas longitudinales, delgadas (0,5 µm de ancho), lisas y de bordes nítidos, que se tornan helicoidales hacia los polos. Se presentan en número de 20 a 22, aproximadamente. Es característico el agrupamiento de las carenas en dos series compactas que se destacan a ambos lados de la apertura.
Material estudiado. Estancia Bajo Tigre, N.F. Ginkgoites, BA Pal. 5026, coord. 46/105; 38,8/112.
Medidas. (2 ejemplares) Cuerpo: largo: 50,6 µm; ancho: 20,9-26,4 µm; espesor exina: 0,6-1,0 µm.
Comparaciones. Ephedripites sp. A se diferencia de Ephedripites acutus sp. nov. y de Ephedripites menendezii Volkheimer 1972 por tener un cuerpo más ancho y mayor número de carenas.
Afinidad botánica. Las especies descriptas para el género Ephedripites se atribuyen a la familia Ephedraceae y son semejantes, entre otros, a los granos de polen actuales de Ephedra helvetica (Kurmann & Zavada, 1994).
Otros taxones ilustrados
Otros taxones adicionales de gimnospermas no han sido descriptos en este texto, aunque sí ilustrados (ver Apéndice). Corresponden a especies de una amplia distribución estratigráfica y geográfica, que ya han sido estudiados en muchas oportunidades por diferentes autores. Constatamos su presencia, con el único objeto de incluirlos en el análisis general de la composición florística de las asociaciones palinológicas de los niveles que fueron analizados a lo largo de la serie que culmina con esta contribución.
Por otra parte, en las asociaciones polínicas de la Formación Anfiteatro de Ticó fueron hallados escasos granos de polen de angiospermas publicados recientemente (Archangelsky & Archangelsky, 2013).
Distribución de briófitas, pteridófitas y gimnospermas halladas en los niveles fosilíferos de la Formación Anfiteatro de Ticó
El análisis polínico de los 8 niveles fosilíferos estudiados (Archangelsky & Gamerro, 1965, 1966 a, b, c; Archangelsky & Villar de Seoane, 1992, 1994, 1996, 1998, 2005) ha permitido determinar que las esporas de pteridófitas dominan en los niveles Brachyphyllum, Williamsonia, Basal y Ticoa harrisii. Las gimnospermas, en cambio, son mayoría en los niveles BTO, Ginkgoites y Athrotaxis. El nivel Pique Carbón muestra la misma proporción de esporas que de granos de polen. Las briófitas sólo se encuentran presentes en los niveles Basal, Williamsonia y Athrotaxis. Por otra parte, el nivel Williamsonia es el más rico en esporas y granos de polen, no sólo por su variedad sino también por su abundancia.
Estos resultados permiten inferir que los niveles Brachyphyllum, Williamsonia, Basal y Ticoa harrisii corresponderían a ambientes boscosos, húmedos y umbrófilos con un importante desarrollo del sotobosque. El nivel Pique Carbón mostraría espacios más abiertos con amplia distribución de árboles, arbustos y sotobosque. Los niveles BTO, Ginkgoites y Athrotaxis tendrían praderas soleadas con buen desarrollo de árboles y arbustos pero sin formación de densos bosques por lo que el sotobosque sería escaso. Los niveles Basal, Williamsonia y Athrotaxis estarían indicando la presencia de ambientes muy húmedos y espejos de agua someros por el desarrollo de briófitas.
Una mención particular merecen las cheirolepidiáceas y ephedráceas, ya que ambos grupos son buenos indicadores de climas semiáridos y áridos (Taylor & Alvin, 1984; Pant & Verma, 1974). Las cheirolepidiáceas están presentes en todos los niveles, mientras que las ephedráceas se hallan restringidas a los niveles Williamsonia y Ginkgoites. Ambos grupos están asociados con elevados porcentajes de pteridófitas, lo cual estaría indicando la marcada estacionalidad (seca/húmeda) existente en la región durante el Aptiano.
Finalmente, cabe señalar que en todos los niveles fosilíferos estudiados en la Formación Anfiteatro de Ticó no fueron hallados elementos de origen marino (quistes de dinoflagelados o radiolarios, entre otros). Ello confirma que las comunidades vegetales analizadas, habitaron en ambientes exclusivamente continentales (Limarino et al., 2012). Entre ellos podemos destacar tres tipos diferentes de ambientes, de acuerdo a los restos vegetales encontrados: a) los existentes al borde de ríos, arroyos y/o espejos de aguas tranquilas; b) bosques con una variable penetración de luz solar; y c) planicies abiertas con exposición solar directa.
CONCLUSIONES
En ésta, la décima y última contribución de la serie, se completa el registro del polen gimnospérmico ya iniciado en trabajos previos (ver Apéndice).
Las gimnospermas constituyeron uno de los principales componentes en las asociaciones palinológicas de la Formación Anfiteatro de Ticó, en la mayor parte de los niveles fosilíferos estudiados.
Se confirma que durante el Aptiano, las cheirolepidiáceas, araucariáceas y podocarpáceas fueron un elemento dominante de los bosques patagónicos, con una marcada diversidad polínica, confirmada por numerosos restos megascópicos, que incluyen conos masculinos en los que se encontraron granos de polen que fueron reconocidos también en estado disperso, en conexión orgánica con ramas foliares. Tales son los casos de las podocarpáceas: Apterocladus lanceolatus Archangelsky (1966) con polen de Callialasporites (Balme) Srivastava (Gamerro, 1965 b), y Trisacocladus tigrensis (Archangelsky 1966) con polen de Trisaccites microsaccatum [(Couper) Couper 1960) (Gamerro 1965)]; las araucariáceas: Alkastrobus peltatus con polen de Cyclusphaera (Del Fueyo & Archangelsky, 2005), y Brachyphyllum irregulare (Archangelsky 1966) con polen de Balmeiopsis (Archangelsky 1966); y la cheirolepidiácea Tomaxellia biforme (Archangelsky 1966) con polen de Classopollis (Archangelsky y Gamerro 1966).
Las Araucariaceae (Araucariacites, Dilwynites) sólo se encuentran en los niveles Williamsonia y Ginkgoites, mientras que Balmeiopsis y Cyclusphaera son más abundantes y aparecen en todos los niveles.
Las Podocarpaceae están representadas por dos tipos de granos. Los bisacados, que son muy abundantes y se registran en todos los ni41 Villar de Seoane & Archangelsky: Palinología de Baqueróveles (Archangelsky & Villar de Seoane, 2005), mientras que, los trisacados tienen una distribución restringida a los niveles Williamsonia, Ginkgoites, BTO, Athrotaxis, Basal y Pique Carbón.
Se ha encontrado una única especie monosacada en los niveles Williamsonia y Pique Carbón, posiblemente también relacionada con las Podocarpaceae.
Las Cheirolepidiaceae, representadas por el género Classopollis, se extinguieron a principios del Paleoceno, aunque fueron dominantes y variadas en especies en todos los niveles fosilíferos de la Formación Anfiteatro de Ticó (Villar de Seoane, en prensa).
Las Ginkgoales, con variedad de cutículas foliares y fructificaciones femeninas (Karkenia incurva Archangelsky), no fueron reconocidas a nivel polínico. En el caso de las Cycadophyta que habitaban los espacios abiertos en las planicies patagónicas, estarían representadas por Cycadopites nitidus muy abundante en el nivel Williamsonia.
Dentro de las Pteridospermales, se destacan las Caytoniales con una importante presencia de Vitreisporites pallidus en el nivel Ginkgoites.
Las Gnetales que fueron cosmopolitas durante el Cretácico temprano, tienen un bajo porcentaje del polen Ephedripites acutus sp. nov. y Ephedripites sp. A, especies restringidas a los niveles Ginkgoites y Williamsonia.
El otro componente importante dentro de las comunidades vegetales de la cuenca, fue el de los helechos y licofitas, los cuales desarrollaron un denso y variado sotobosque. Dominaron en los niveles fosilíferos Williamsonia, Brachyphyllum, Basal y Ticoa harrisii.
Lista de especies identificadas
Ticoites gamerroi nov. gen. et sp. Figuras 1A-E
Vitreisporites pallidus (Reissinger) Nilsson 1958. Figuras 1F-G
Microcachryidites antarcticus Cookson 1947. Figura 1J
Microcachryidites castellanosii Menéndez 1968. Figuras 1H-I
Trisaccites microsaccatum (Couper) Couper 1960. (Utilizamos este género en lugar de Trichotomosulcites por haber sido este último mal determinado originalmente por Couper, 1953) Figuras 1K-L
Araucariacites australis Cookson 1947. Figuras 2A-B
Araucariacites spinosus sp. nov. Figuras 2C-E
Dilwynites granulatus Harris 1965. Figura 2F
Cycadopites nitidus (Balme) de Jersey 1964. Figuras 2G-I
Ephedripites acutus sp. nov. Figuras 2K-L
Ephedripites sp. A. Figura 2J
Apéndice Final
Listado de todas las especies de esporas y granos de polen hallados y descriptos en la Formación Anfiteatro de Ticó por Archangelsky, 1994 (1); Archangelsky & Archangelsky, 2013 (2); Archangelsky & Gamerro, 1965 (3); Archangelsky & Gamerro, 1966 (4); Archangelsky & Gamerro, 1966a (5); Archangelsky & Gamerro, 1966b (6); Archangelsky & Gamerro, 1966c (7); Archangelsky & Gamerro, 1967 (8); Archangelsky & Villar de Seoane 1989 (9); Archangelsky & Villar de Seoane 1990 (10); Archangelsky & Villar de Seoane, 1991 (11); Archangelsky & Villar de Seoane, 1992 (12); Archangelsky & Villar de Seoane, 1994 (13); Archangelsky & Villar de Seoane, 1996 (14); Archangelsky & Villar de Seoane, 1998 (15); Archangelsky & Villar de Seoane, 2005 (16); Del Fueyo & Archangelsky, 2005 (17); Gamerro, 1965a (18); Gamerro, 1965b (19); Taylor & Taylor (20); Villar de Seoane (21).
DIVISION HEPATOPHYTA
Aequitriradites baculatus Döring (3)
Aequitriradites spinulosus (Cookson & Dettmann) Cookson & Dettmann (3)
Aequitriradites verrucosus (Cookson & Dettmann) Cookson & Dettmann (3)
Coptospora foveolata Archangelsky & Villar de Seoane (14)
Naiaditaspora gemmata Archangelsky & Villar de Seoane (14)
Rouseisporites reticulatus Pocock (6)
Staplinisporites caminus (Balme) Pocock (14)
Taurocusporites segmentatus Stover (6)
DIVISION ANTHOCERATOPHYTA
Chomotriletes minor (Kedves) Pocock (14)
Foraminisporis asymmetricus (Cookson & Dettmann) Dettmann (14)
Foraminisporis dailyi (Cookson & Dettmann) Dettmann (5)
Stoverisporites lunaris (Cookson & Dettmann) Norvick & Burger (14)
DIVISION LYCOPHYTA
Orden LYCOPODIALES
Auritulinasporites sp. A Archangelsky & Villar de Seoane (15)
Camarazonosporites microalveolatus Archangelsky & Gamerro (6)
Concavisporites cf. laticrassus Volkheimer (15)
Concavisporites sp. A Archangelsky & Villar de Seoane (15)
Densoisporites corrugatus Archangelsky & Gamerro (3)
Obtusisporis obtusangulus (Krutzsch) Pocock (15)
Obtusisporites sp. A Archangelsky & Villar de Seoane (15)
Reticulatisporites ornatus Archangelsky & Villar de Seoane (15)
Retitriletes austroclavatidites (Cookson) Döring et al. (15)
Retitriletes baqueroense Archangelsky & Villar de Seoane (15)
Retitriletes sp. A Archangelsky & Villar de Seoane (15)
Sestrosporites pseudoalveolatus (Couper) Dettmann (6)
Orden SELAGINELLALES
Antulsporites baculatus Archangelsky & Gamerro (5)
Bacutriletes guttula Archangelsky & Villar de Seoane (11)
Bacutriletes triangulatus Taylor & Taylor (20)
Bacutriletes sp. A Taylor & Taylor (20)
Cabochonicus carbunculus (Dijkstra) Batten & Ferguson (11)
Ceratosporites equalis Cookson & Dettmann (13)
Ceratosporites setosus Archangelsky & Villar de Seoane (13)
Horstisporites iridodea Taylor & Taylor (20)
Orden ISOETALES
Auriculozonospora spongiosa Archangelsky & Villar de Seoane (10)
Hughesisporites patagonicus Archangelsky (10) (20)
Hughesisporites rugulatus Archangelsky & Villar de Seoane (11)
Minerisporites elegans Archangelsky & Villar de Seoane (9)
Minerisporites laceratus Archangelsky & Villar de Seoane (10)
Minerisporites patagonicus Archangelsky & Villar de Seoane (9)
Verrutriletes tomtaylori Archangelsky & Villar de Seoane (10)Revista del Museo Argentino de Ciencias Naturales, n. s. 16(1), 2014 44
DIVISIÓN SPHENOPHYTA
Familia EQUISETACEAE
Calamospora mesozoica Couper (13)
DIVISION PTERIDOPHYTA
Familia OSMUNDACEAE
Baculatisporites comaumensis (Cookson) Potonié (13)
Osmundacidites diazii Volkheimer (13)
Osmundacidites sp. A Archangelsky & Villar de Seoane (13)
Rugulatisporites neuquenensis Volkheimer (13)
Rugulatisporites sp. A Archangelsky & Villar de Seoane (13)
Rugulatisporites sp. B Archangelsky & Villar de Seoane (13)
Familia SCHIZAEACEAE
Cicatricosisporites annulatus Archangelsky & Gamerro (7)
Cicatricosisporites australiensis (Cookson) Potonié (7)
Cicatricosispòrites baqueroensis Archangelsky & Gamerro (7)
Cicatricosisporites giganteus Archangelsky & Gamerro (7)
Cicatricosisporites hughesii Dettmann (7)
Cicatricosisporites ticoensis Archangelsky & Gamerro (7)
Contignisporites cooksonii (Balme) Dettmann (5)
Microfoveolatosporis sp. A Archangelsky & Villar de Seoane (15)
Trilobosporites apiverrucatus Couper (6)
Trilobosporites purverulentus (Verbitskaya) Dettmann (6)
Trilobosporites trioreticulosus Couper (6)
Vallizonosporites spinosus Archangelsky & Villar de Seoane (15)
Familia GLEICHENIACEAE
Clavifera sp. A Archangelsky & Villar de Seoane (12)
Gleicheniidites senonicus Ross (12)
Gleicheniidites sp. A Archangelsky & Villar de Seoane (12)
Gleicheniidites sp. B Archangelsky & Villar de Seoane (12)
Laevigatosporites belfordii Burger (15)
Laevigatosporites ovatus Wilson & Webster (15)
Ornamentifera tuberculata (Grigorjeva) Bolchovitina (12)
Ornamentifera sp. A Archangelsky & Villar de Seoane (12)
Familia DICKSONIACEAE
Cibotiidites auriculatus Archangelsky & Villar de Seoane (15)
Familia LOPHOSORIACEAE
Cyatheacidites tectifera Archangelsky & Gamerro (3)
Familia CYATHEACEAE
Cyathidites australis Couper (13)
Cyathidites minor Couper (13)
Cyathidites sp. A Archangelsky & Villar de Seoane (13)
Cyathidites sp. B Archangelsky & Villar de Seoane (13)
Deltoidospora minor (Couper)Pocock (13)
Leiotriletes regularis (Pflug) Krutzsch (13)
Familia PTERIDACEAE
Biretisporites sp. A Archangelsky & Villar de Seoane (13)
Familia DENNSTAEDTIACEAE
Leptolepidites major Couper (12)
Neoraistrickia sp. A Archangelsky & Villar de Seoane (13)
Verrucosisporites sp. A Archangelsky & Villar de Seoane (13)
Familia DRYOPTERIDACEAE
Tuberculatosporites sp. A Archangelsky & Villar de Seoane (15)
Incertae Sedis
Polypodiaceoisporites elegans Archangelsky & Gamerro (5)
Polypodiidites sp. A Archangelsky & Villar de Seoane (15)
Polypodiidites sp. B Archangelsky & Villar de Seoane (15)
Rotverrusporites labratus Archangelsky & Villar de Seoane (15)
DIVISION CONIFEROPHYTA
Familia ARAUCARIACEAE
Araucariacites australis Cookson (1)
Balmeiopsis limbatus (Balme) Archangelsky (1)
Cyclusphaera psilata Volkheimer & Sepúlveda (17)
Familia CHEIROLEPIDIACEAE
Classopollis asper Villar de Seoane (21)
Classopollis grandis Villar de Seoane (21)
Classopollis bussoni Reyre (21)
Classopollis chateaunovii Reyre (21)
Classopollis kieseri Reyre (21)
Classopollis noeli Reyre (21)
Classopollis pujoli Reyre (21)
Classopollis quezeli Reyre (21)
Classopollis rarus Reyre (21)
Classopollis triangulus Reyre (21)
Classopollis sp. A Villar de Seoane (21)
Familia PODOCARPACEAE
Callialasporites dampieri (Balme) Sukh Dev (1) (19)
Callialasporites trilobatus (Balme) Sukh Dev (19)
Gamerroites volkheimeri Archangelsky (16)
Gamerroites sp. A Archangelsky & Villar de Seoane (16)
Podocarpidites auriculatus Archangelsky & Villar de Seoane (16)
Podocarpidites ellipticus Cookson (16)
Podocarpidites futa Archangelsky & Villar de Seoane (16)
Podocarpidites marwickii Couper (16)
Podocarpidites parviauriculatus Archangelsky & Villar de Seoane (16)
Podocarpidites vestitus Archangelsky & Villar de Seoane (16)
Podocarpidites sp. A Archangelsky & Villar de Seoane (16)
Trisaccites microsaccatum (Couper) Couper (18)
División MAGNOLIOPHYTA
Clavatipollenites sp. 1 Archangelsky & Archangelsky (2)
Clavatipollenites sp. 2 Archangelsky & Archangelsky (2)
Clavatipollenites sp. 3 Archangelsky & Archangelsky (2)
Clavatipollenites sp. 4 Archangelsky & Archangelsky (2)
Retimonocolpites sp. Archangelsky & Archangelsky (2)
Tipo Anacostia Archangelsky & Archangelsky (2)
AGRADECIMIENTOS
Los autores desean expresar su agradecimiento a la Directora de la Revista, Dra. Silvia Césari, y los revisores, Dres. Eduardo Ottone y Mirta Quattrocchio por las valiosas observaciones realizadas en el manuscrito; y al Sr. Orlando Cárdenas por el procesamiento de las muestras palinológicas. Este estudio fue realizado con los subsidios PICT 433 - 2007 y PICT 528-2012 otorgados por la Agencia Nacional de Promoción Científica y Tecnológica; y el PIP 679 - 2009 financiado por el CONICET.
BIBLIOGRAFIA
1. Archangelsky, S. 1966. New gymnosperms from the Ticó flora, Santa Cruz Province, Argentina. Bull. Brit. Mus. (Nat. Hist.) Geol. 13: 261-295. [ Links ]
2. Archangelsky, S. 1994. Comparative ultrastructure of three Early Cretaceous gymnosperm pollen grains: Araucariacites, Balmeiopsis and Callialasporites. Rev. Palaeobot. Palynol. 83: 185-198. [ Links ]
3. Archangelsky, S. & A. Archangelsky. 2013. Aptian angiosperm pollen from the Ticó Flora, Patagonia, Argentina. Int. J. Plant Sci. 174: 559-571. [ Links ]
4. Archangelsky, S. & J.C. Gamerro. 1965. Estudio palinológico de la Formación Baqueró (Cretácico), provincia de Santa Cruz. I. Ameghiniana 4: 159-170. [ Links ]
5. Archangelsky, S. & J.C. Gamerro. 1966 a. Estudio palinológico de la Formación Baqueró (Cretácico), provincia de Santa Cruz. II. Ameghiniana 4: 201-209. [ Links ]
6. Archangelsky, S. & J.C. Gamerro. 1966 b. Estudio palinológico de la Formación Baqueró (Cretácico), provincia de Santa Cruz. III. Ameghiniana 4: 229-236. [ Links ]
7. Archangelsky, S. & J.C. Gamerro. 1966 c. Estudio palinológico de la Formación Baqueró (Cretácico), provincia de Santa Cruz. IV. Ameghiniana 4: 363-372. [ Links ]
8. Archangelsky, S. & J.C. Gamerro. 1967. Spore and pollen types of the Lower Cretaceous in Patagonia (Argentina). Rev. Palaeobot. Palynol. 1: 211-217. [ Links ]
9. Archangelsky, S. & L. Villar de Seoane. 1989. Ultraestructura de dos nuevas megasporas cretácicas de Santa Cruz, Argentina. Bol. Asoc. Latinoam. Paleobot. Palinolog. 12: 1325. Buenos Aires. [ Links ]
10. Archangelsky, S. & L. Villar de Seoane. 1990. Morfología y estructura de megasporas cretácicas de Patagonia, República Argentina. Rev. Española Micropal. 22 (3): 419450. Madrid. [ Links ]
11. Archangelsky, S. & L. Villar de Seoane. 1991. Notas sobre la flora fósil de la Zona de Ticó, Provincia de Santa Cruz. XI. Morfología y estructura de tres megasporas. Ameghiniana 28 (34): 353364. Buenos Aires. [ Links ]
12. Archangelsky, S. & L. Villar de Seoane. 1992. Estudios palinológicos de la Formación Baqueró (Cretácico), Provincia de Santa Cruz, Argentina. V. Asoc. Paleontol. Argent., Publ. Espec. Nº 2. VIII Simp. Argentino Paleob. Palin. 2327. Buenos Aires. [ Links ]
13. Archangelsky, S. & L. Villar de Seoane. 1994. Estudios palinológicos de la Formación Baqueró (Cretácico), Provincia de Santa Cruz, Argentina. VI. Ameghiniana 31: 4153. Buenos Aires. [ Links ]
14. Archangelsky, S. & L. Villar de Seoane. 1996. Estudios palinológicos de la Formación Baqueró (Cretácico), Provincia de Santa Cruz, Argentina. VII. Ameghiniana 33: 307-314. Buenos Aires. [ Links ]
15. Archangelsky, S. & L. Villar de Seoane. 1998. Estudios palinológicos de la Formación Baqueró (Cretácico), Provincia de Santa Cruz, Argentina. VIII. Ameghiniana 35: 7-19. Buenos Aires. [ Links ]
16. Archangelsky, S. & L. Villar de Seoane. 2005. Estudios palinológicos del Grupo Baqueró (Cretácico Inferior), Provincia de Santa Cruz, Argentina. IX. Polen bisacado de Podocarpaceae. Revista Española de Paleontología 20: 37-56. [ Links ]
17. Bolkhovitina, N.A. 1953. Spore-pollen assemblages of the Cretaceous of the Central part of the U.S.S.R. Trud. Akad. Nauk. S.S.S.R., s. Geol. 61: 1-183. (en ruso). [ Links ]
18. Cladera, G., R. Andreis, S. Archangelsky & R. Cúneo. 2002. Estratigrafía del Grupo Baqueró, Patagonia (provincia de Santa Cruz, Argentina). Ameghiniana 39: 3-20. [ Links ]
19. Cookson, I.C. 1947. Plant microfossils from the lignites of the Kerguelen Archipelago. British, Australian, New Zealand Antarctic Research Expedition (1929-31) Rep. A., 2: 129-142. [ Links ]
20. Corbella, H. 2001. Tuffs of the Baqueró Group and the Mid-Cretaceous frame, Extraandean Patagonia, Argentina. XI Congreso Latinoamericano de Geología y III Congreso Uruguayo de Geología. Trabajo 190 pp. (En CD). [ Links ]
21. Couper, R.A. 1953. Upper Mesozoic and Cainozoic spores and pollen grains from New Zealand. N. Zeal. Geol. Surv., Paleont. Bull. 22: 77 pp. [ Links ]
22. Couper, R.A. 1960. New Zealand Mesozoic and Cainozoic plant microfossils. N. Zeal. Geol. Surv., Paleont. Bull. 32: 1-82. [ Links ]
23. Del Fueyo, G.M. & S. Archangelsky. 2005. A new araucarian pollen cone with in situ Cyclusphaera Elsik from the Aptian of Patagonia, Argentina. Cret. Res. 26:757-768. [ Links ]
24. Dettmann, M.E. & D.M. Jarzen. 2000. Pollen of extant Wollemia (Wollemi Pine) and comparisons with pollen of other extant and fossil Araucariaceae. En: M.M. Harley, C.M. Morton and S. Blackmore (Eds.). Pollen and Spores: Morphology and Biology p.p. 187-203. Royal Botanic Gardens, Kew. [ Links ]
25. Erdtman, G. 1947. Suggestions for the classification of fossil and recent pollen grains and spores. Svensk Bot. Tidskr. 41: 104-114. [ Links ]
26. Gamerro, J.C. 1965 a. Morfología del polen de la conífera Trisacocladus tigrensis Archang. de la Formación Baqueró, Provincia de Santa Cruz. Ameghiniana 4: 31-38. [ Links ]
27. Gamerro, J.C. 1965b. Morfología del polen de Apterocladus lanceolatus Archang. (Coniferae) de la Formación Baqueró, Provincia de Santa Cruz. Ameghiniana 4:133-138. [ Links ]
28. Harris, W.K. 1965. Basal Tertiary microfloras from the Princetown area, Victoria, Australia. Palaeontographica B 115: 75-106. [ Links ]
29. Jones, W.G., K.D. Hill & J.M. Allen. 1995. Wollemia nobilis, a new living Australian genus and species in the Araucariaceae. Telopea 6:173-176. [ Links ]
30. Kotova, I.Z. 1983. Palynological study of Upper Jurassic and Lower Cretaceous sediments, site 511, deep sea drilling Project Leg 71 (Falkland Plateau), Initial Reports of the Deep Sea Drilling Project. Leg. LXXI (2), 32: 879-906. [ Links ]
31. Krüssmann, G. 1985. Manual de cultivated Conifers. H.D. Warda (ed.) Timber Press, Portland, Oregon, 361 p.p. [ Links ]
32. Kurmann, M.H. & M.S. Zavada. 1994. Pollen morphological diversity in extant and fossil gymnosperms. En: M.H. Kurmann and J.A. Doyle (eds.). Ultrastructure of fossil spores and pollen, pp. 123-137, Royal Botanic Gardens, Kew. [ Links ]
33. Limarino, C.O., M.G. Passalia, M. Llorens, E.I. Vera, V. Perez Loinaze & S. Césari. 2012. Depositional environments and vegetation of Aptian sequences affected by volcanism in Patagonia. Palaeogeog., Palaeoclimat., Palaeoecol. 323-325: 22-41. [ Links ]
34. Macphail, M., R.J. Carpenter, A. Iglesias & P. Wilf. 2013. First evidence for Wollemi Pine-type pollen (Dilwynites: Araucariaceae) in South America. PLoS ONE, 8(7): e69281. doi:10.1371/journal.pone.0069281. [ Links ]
35. Naumova, S.N. 1939. Spores and pollen of coals of the U.S.S.R. Rep. Int. Geol. Congr., 17 th Session U.S.S.R. 1: 353-364. [ Links ]
36. Ottone, E.G., P.P. Alvarez & S.V. Benoit. 1992. Late Triassic plant microfossils from the Rancho de Lata Formation, Main Cordillera, Argentina. Micropaleontology 38: 261-278. [ Links ]
37. Ottone, E.G., A.C. Mancuso & M. Resano. 2005. Miospores and chlorococcalean algae from the Los Rastros Formation, Middle to Upper Triassic of central-western Argentina. Ameghiniana 42: 347-362. [ Links ]
38. Pant, D.D. & B.K. Verma. 1974. Taxonomy of the genus Ephedra. Significance of stem and leaf epidermis and cuticle. Bot. J. Linnean Soc. 69: 287-308. [ Links ]
39. Pérez Loinaze, V.S., E.I. Vera, M.G. Passalia, M. Llorens, R. Friedman, C.O. Limarino. & S.N. Césari. 2013. High-precision Ue-Pb zircon age from the Anfiteatro de Ticó Formation: Implications for the timing of the early angiosperm diversification in Patagonia. J. South Am. Earth Sci. 48: 97-105. [ Links ]
40. Pocock, S.A.J., G. Vasanthy & B.S. Venkatachala. 1990. Pollen of Circumpolles - An enigma or morphotrends showing evolutionary adaptation. Rev. Palaeobot. Palynol. 65: 179-193. [ Links ]
41. Potonié, R. & G. Kremp. 1954. Die Gattungen der paläozoischen Sporae dispersae und ihre Stratigraphie. Geol. Jahrb. 69: 111-194. [ Links ]
42. Potonié, R. 1958. Synopsis der Gattungen der Sporae dispersae. II Beih. Geol. Jb. 31: 1-114. [ Links ]
43. Potonié, R. 1960. Synopsis der Gattungen der Sporae dispersae. III Beih. Geol. Jb. 39: 1-189. [ Links ]
44. Potonié, R. 1970. Synopsis der Gattungen der Sporae dispersae. V. Ibidem, 87: 1-156. [ Links ]
45. Prámparo, M.B. & W. Volkheimer. 2002. Nuevos hallazgos de palinomorfos en la Formación La Amarga, Miembro Bañados de Caichigue: cuenca Neuquina sudoriental, Argentina. Ameghiniana 39: 395-404. [ Links ]
46. Taylor, T.N. & K.L. Alvin. 1984. Ultrastructure and development of Mesozoic Pollen: Classopollis. Am. J. Bot. 71: 575-587. [ Links ]
47. Taylor, T. N. & W. Taylor. 1988. Ultrastructural análisis of selected Cretaceous megaspores from Argentina. J. Micropal. 7: 73-87. [ Links ]
48. Taylor,T.N., M.S. Zavada & S. Archangelsky. 1989. The ultrastructure of Cyclusphaera psilata from the Cretaceous of Argentina. Grana 26: 74-80. [ Links ]
49. Villar de Seoane, L. (en prensa) Estudio morfológico del género Classopollis (Pflug) Pocock y Jansonius (Cretácico, Argentina). Rev. Bras. Paleont. [ Links ]
50. Volkheimer, W. 1968. Esporas y granos de polen del Jurásico de Neuquén (República Argentina). I. Descripciones sistemáticas. Ameghiniana 5: 333-370. [ Links ]
51. Volkheimer, W. 1972. Estudio palinológico de un carbón caloviano de Neuquén y consideraciones sobre los paleoclimas jurásicos de la Argentina. Rev. Mus. La Plata (N.S.) 6: 101-157. [ Links ]
Recibido: 27-11-2013
Aceptado: 27-03-2014
