










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
versión On-line ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.16 no.2 Ciudad Autónoma de Buenos Aires dic. 2014
ECOLOGÍA
Máximo térmico crítico en peces argentinos de agua dulce, Sudamérica
Sergio E. Gómez
Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" (CONICET), Av. Ángel Gallardo 470 (DJR1405) Ciudad Autónoma de Buenos Aires, Argentina, gomezsergioe@yahoo.com
Resumen
Utilizando la técnica de máximo térmico critico, con calentamiento de 18° C/hora y partiendo de muy diferentes temperaturas de aclimatación, se determinaron los valores de temperatura de pérdida de equilibrio y temperaturas letales en ocho especies de peces pampásicos. Los resultados muestran una relación fisiológica con los distintos "tipos ecológicos" en relación a la temperatura. Los peces de aguas libres o litorales muestran la mayor tolerancia y los peces de fondo la menor, de acuerdo a la estabilidad del ambiente.
Palabras clave: Máximo térmico critico; Peces pampásicos; Tipos ecológicos; Sudamérica.
Abstract
Critical thermal maximum in Argentine freshwater fish, South America. Using the critical thermal maximum technique with heating at 18°C/hour and from very different temperatures acclimation, the lethal temperature values and loss of equilibrium temperatures in eight pampasic species were identified. The results show a physiological relationship with the various "ecological types" in relation to temperature. The open-water fish and shore fish show greater tolerance than lower bottom fish, according to the stability of the environment.
Key words: Critical thermal maximum; Pampasic fish; Ecological types; South America.
INTRODUCCIÓN
El interés por cuantificar la tolerancia de los peces a las altas temperaturas proviene desde principios del siglo XX (Hathaway, 1927). Es un tema dificultoso con abundante bibliografía donde se desarrollan diversas técnicas, las que fueron compiladas y aplicadas por Fry (1971); en todos los casos la temperatura de aclimatación es la variable más importante. Los peces tienen la particularidad de variar sus límites de letalidad en función de la temperatura de aclimatación, ésta es la temperatura a la que han estado expuestos por períodos prolongados. Se ha determinado que la velocidad de aclimatación es de 1° C/ día (Brett, 1946), pudiendo ser mucho más rápida (Gómez, 1996).
Entre otras, la técnica de "máximo critico" es útil ya que el calentamiento rápido, usualmente de 18° C/hora (= 3° C/minuto), no permite la aclimatación de los peces a temperaturas intermedias durante la experiencia (Becker & Genoway, 1979; Paladino et al., 1980; Elliot, 1981). Por lo tanto, al no ser posible la aclimatación los resultados entre especies son comparables. Otros autores han usado distintas velocidades de calentamiento, Arrueda Freire & Prodocimo (2001) documentan experimentos realizados a 1° C/min; velocidades mayores no permiten una correcta observación de los peces.
Este método fue objetado en relación a otros como, "concentración letal para el 50%" y "tiempo de resistencia" (Lutterschimdt & Hutchinson, 1997). Por el contrario, Kilgour & McCauley (1986) demostraron que las técnicas de "concentración letal para el 50%" y "máximo térmico critico" proporcionan resultados equivalentes e interconvertibles.
Cuando la temperatura aumenta muy rápido experimentalmente, los peces incrementan su movilidad y ritmo respiratorio, posteriormente sobreviene la "perdida de equilibrio" caracterizada por arritmia respiratoria y falta de coordinación motriz. A los pocos grados mueren, este punto se reconoce por el cese total de movimientos operculares. Cualquiera de los dos puntos, perdida de equilibrio o muerte, puede tomarse como "máximo térmico critico".
En la Argentina el conocimiento sobre temperaturas letales es muy limitado, para la zona patagónica se encuentran los trabajos de Gómez (1990) y Ortubay et al. (1997) y para la zona parano-platense de Dioni & Reartes (1975) y Gómez (1996).
En ictiogeografía argentina la temperatura es fundamental para delimitar tres zonas: Parano-platense, Patagónica y Andino-Cuyana (Cussac et al., 2009). En la zona Parano-platense se reconoce la "pampasia" o ecorregión de las pampas que incluye las llanuras del este de la Argentina entre los 33° y 39° de latitud sur. La temperatura media anual es 13° a 17° Ccon medias mínimas y máximas de 9° y 25° Crespectivamente. Es una región muy extensa con varios ríos y cientos de lagunas (Canevari et at., 1998).
De acuerdo al hábitat, morfología, alimentación y modo de vida los peces pueden agruparse en distintos "tipos ecológicos". Este concepto fue desarrollado por Ringuelet, (1975) y ampliado por Menni (2004), quien distinguió distintos tipos ecológicos de peces; tres de ellos son pertinentes en este trabajo. El criterio fue muy utilizado, (López et al., 2006), a veces con nombres similares. Muñoz (2012) reconoce cinco "categorías ecológicas" en el estuario del Río de La Platamientras que Guzmán & Lyons (2002) agrupan los peces en ocho "grupos ecológicos". Los estudios se concentran en los atributos indicados, pero no se avanzó en la ecofisiología de los distintos tipos ecológicos.
En este trabajo se determina el máximo térmico critico en ocho especies Parano-platenses, relacionando los resultados con los tipos ecológicos representados (sensu Menni, 2004), analizando posibles relaciones ecofisiológicas.
MATERIALES Y MÉTODOS
Se desarrollaron 27 experimentos en ocho especies autóctonas, capturadas en el campo, con un total de 116 ejemplares. Los peces fueron aclimatados, usualmente en grupos de cinco individuos, durante 10 días a una temperatura constante (temperatura de aclimatación= TA) antes de la experiencia, entre 9,43° y 32,13° C. Se utilizó una tasa de calentamiento promedio de 18,00° C/hora (rango= 16,2° a 19,4° C). La temperatura se controló con termómetros de mercurio intercalibrados a 0,05° Cy para el calentamiento se utilizaron dos resistencias eléctricas y un regulador de la intensidad de corriente. La experiencia se desarrolla con la introducción de los ejemplares en una cámara de vidrio de 20 litros de capacidad y a la misma temperatura que la de aclimatación. En la cámara la temperatura se mantiene homogénea y con aireación suficiente mediante una bomba de 300 litros/hora de capacidad.
Las especies utilizadas fueron Cnesterodon decemmaculatus, Jenynsia multidentata, Odontesthes bonariensis, Australoheros facetus, Gymnogeophagus meridionalis, Corydoras paleatus, capturados en laguna Chascomús (35° 34' LS; 58° 01' LW). Los ejemplares de Callichthys callichthys y Rhinodoras d'orbigny, fueron capturados en el delta del paraná bonaerense.
Durante la experiencia, cuando la temperatura aumenta a niveles altos, los peces primeramente pierden el ritmo respiratorio y luego la capacidad de natación coordinada; el promedio aritmético de los valores individuales del grupo se denomina "temperatura de pérdida de equilibrio" (TPE). La muerte se caracteriza por el cese total de movimientos operculares, el promedio aritmético de los valores individuales se denomina "temperatura de muerte" (TM) del grupo. Para cada caso se calculó TPE, TM, se midió la longitud estándar individual y se registró el valor medio (LE) del grupo.
RESULTADOS
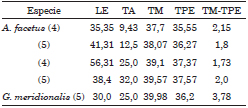
Para cada grupo experimental se tabularon los resultados indicando LE, TA, TM, TPE y la diferencia TM-TPE. De acuerdo a su modo de vida y morfología se identificaron tres "tipos ecológicos" distintos de peces pampásicos, de acuerdo a Menni (2004). Peces de superficie: Cnesterodon decemmaculatus, Jenynsia multidentata y Odontesthes bonariensis (Tabla 1) peces de fondo: Corydoras paleatus, Callichthys callichthys y Rhinodoras d'orbigny (Tabla 2) y peces litorales de aguas vegetadas: Australoheros facetus y Gymnogeophagus meridionalis (Tabla 3).
Tabla 1. Peces de superficie. Para cada grupo de 3 especies se indica numero de ejemplares (entre paréntesis), longitud estándar media del grupo (LE), temperatura de aclimatación (TA), temperatura de muerte (TM), temperatura de perdida de equilibrio (TPE) y diferencia entre TM y TP. LE en mm, temperaturas en ° C, total 55 individuos.

Tabla 2. Peces de fondo. Abreviaturas como en Tabla 1, 10 grupos, total 43 individuos.

Tabla 3. Peces litorales y de aguas vegetadas. Abreviaturas como en Tabla 1, 5 grupos, total 23 individuos.
Los datos, para cada tipo ecológico, se graficaron utilizando la temperatura de aclimatación como variable independiente y la temperatura de muerte o temperatura de pérdida de equilibrio, como dependiente. En cada caso se ajustó una relación logarítmica (Figs. 1, 2 y 3) utilizando técnicas de regresión y correlación. En el examen de los gráficos de dispersión en ningún caso se detectó relación de TM o TPE con el tamaño corporal.

Fig. 1. Relación entre temperatura de aclimatación y temperatura de perdida de equilibrio (TPE) en peces de superficie (círculos; r= 0,7792 n.s.) y peces de fondo (cuadrados; r= 0,9283 s. p<0,05). R2 es el coeficiente de determinación de cada curva.

Fig. 2. Relación entre temperatura de aclimatación y temperatura letal (TM) en peces pampásicos de superficie (cuadrados; r= 0,8234 s. p<0,05) y temperatura de pérdida de equilibrio (TPE); (círculos; r= 0,7792 n.s.). R2 es el coeficiente de determinación de cada curva.
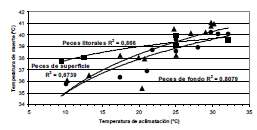
Fig. 3. Relación entre temperatura de aclimatación y temperatura letal en peces pampásicos. Peces de superficie (triángulos, r= 0,8209 s. p<0,05), peces litorales de aguas vegetadas (cuadrados, r= 0,9306 s. p<0,05) y peces de fondo (círculos, r= 0,8988 s. p<0,05). R2 es el coeficiente de determinación de cada curva.
La correlación entre temperatura de aclimatación (TA) y temperatura de muerte (TM) fue significativa en los tres grupos (p<0,05). Las curvas de regresión (Fig. 3) mostraron coeficientes de determinación (R2) elevados.
DISCUSIÓN Y CONCLUSIONES
El agua es el único elemento en la tierra, que por efecto de la temperatura ambiente, puede existir en forma sólida, líquida y gaseosa. La temperatura condiciona o determina fuertemente los modos de vida de los organismos acuáticos y es considerado un factor primario de supervivencia. Al aumentar la temperatura los peces sufren estrés térmico que es fácilmente detectable con la técnica de máximo crítico (Elliot, 1981). El estrés térmico produce múltiples efectos, se destaca el importante cambio en los parámetros hematológicos (Docan et al., 2013). Los valores de resistencia muestran una gran variedad interespecífica. El caso mas extremo conocido de resistencia es Cyprinus diabolis, que vive en aguas termales de Norteamérica a 33,9° Cy su límite letal es de 43° C.
Los peces mantienen características ecofisiológicas de su lugar de origen. Caverth et al. (2006) trasplantaron ejemplares criados en aguas templadas a otros ambientes de aguas mas frías. Los ejemplares de aguas templadas siguieron manteniendo un límite de resistencia más alto que ejemplares co-específicos de aguas más frías.
Los resultados experimentales aquí presentados fueron obtenidos en condiciones de laboratorio y pueden no representar la real plasticidad de las especies en la naturaleza.
El aumento de la temperatura de aclimatación produce un aumento de la temperatura de pérdida de equilibrio y de la temperatura de muerte. Este aumento no es lineal y poco evidente con altas temperaturas de aclimatación, que en los tipos ecológicos de peces pampásicos estudiados es de 30° Caproximadamente. Los peces de superficie tienen una TPE mayor que los peces de fondo, la diferencia promedio (TM-TPE) es de 1,3° y 1,6° Crespectivamente (Fig. 1). Un caso singular es Jenynsia multidentata donde TPE y TM son coincidentes (Tabla 1), no hay una explicación clara para este caso de muerte sin síntomas previos.
La diferencia entre temperatura de muerte y temperatura de pérdida de equilibrio (TM-TPE), que usualmente es como máximo de 2° o 3° C, es una variable poco estudiada. Gómez (1990) en Hatcheria macraei, un siluriforme patagónico de abolengo brasílico, encontró una diferencia mucho mayor que fue calificada como relicto fisiológico.
Peces de superficie mostraron una TPE promedio de 37,2° y una TM promedio de 38,55° C, en ambos casos la relación con TA no es lineal (Fig. 2). Cuanto mayor es la TA la diferencia entre TM y TPE es menor. Resultados análogos se obtienen con los otros grupos ecológicos. Se deben diseñar experiencias de recuperación para determinar en que punto el proceso de muerte térmica es reversible.
Cuando se analiza la TM de los tres grupos en función de la TA (Fig. 3), se observa que hasta una aclimatación de 25° C, los tres grupos se distinguen claramente: los peces de superficie y litorales son los más resistentes con alta variación, dependiendo de la TA, los peces de fondo son los menos resistentes. Frente a situaciones climáticas adversas, los peces de superficie pueden cambiar su posición en la columna de agua buscando estratos más favorables.
Con temperaturas mayores a 25° Clos tres tipos ecológicos se mezclan con TM entre 39° y 41° Cdonde la TA tiene poca influencia. Es destacable que 25° Ces la temperatura ambiente media máxima en lagunas pampásicas. Esto puede interpretarse como una adaptación óptima a los ambientes lagunares.
La tolerancia a la temperatura esta relacionada con la estabilidad y fluctuaciones del ambiente. En peces marinos los que están estrictamente asociados a los arrecifes de coral muestran la menor zona de tolerancia térmica, mientras que las especies residentes o transitorias de la zona intermareal tienen una mayor tolerancia térmica (Eme & Bennet, 2009).
Si se aplica el criterio de Arrueda Freire & Prodocimo (2001) para distinguir peces dulceacuícolas tropicales de peces de aguas templadas; las especies aquí estudiadas pertenecen a aguas templadas. El hábitat parano-platense siempre se reconoció por su inestabilidad, además de los efectos del cambio ambiental (Gómez et al., 2008), en estos ambientes las especies mas euritópicas y resistentes son dominantes. Este trabajo muestra que a mayor inestabilidad del hábitat mayor tolerancia térmica. Donde los peces de superficie y de zonas litorales vegetadas, muy expuestos a variables climáticas inestables son los más tolerantes, y los peces de fondo, con un hábitat muy estable, son los menos tolerantes.
BIBLIOGRAFIA
1. Arrueda Freire, C. & V. Prodocimo. 2001. Critical termal maxima and minima of the platyfish Xiphophorus maculatus Günther (Poecillidae, Cyprinodontiformes) – a tropical species of ornamental freshwater fish. Revista Brasileira de Zoologia (supl. 1): 97-106.
2. Becker, C.D., & R.G. Genoway. 1979. Evaluation of the critical thermal maximum for determining thermal tolerance of freshwater fish. Environmental Biology of Fishes, Dordrech 4(3): 245-246. [ Links ]
3. Brett, J.R. 1946. Rate of gain of heat-tolerance in goldfish (Carassius auratus). Univ. of Toronto Studies Biological Series. 53: 1-28. [ Links ]
4. Brown, J.H. & C.R. Feldmeth. 1971. Evolution in constant and flutuating environments: thelmal tolerance ofdesert pupfish (Cyprinodon). Evolution 25: 390-398. [ Links ]
5. Canevari, P., D.E. Blanco, E. Bucher, G. Castro & I. Davison (eds.). 1998. Los humedales de la Argentina. Clasificación, situación actual, conservación y legislación. Wetlands International. Publicación N°46, 208 pp. [ Links ]
6. Carveth, J.C., A.M. Widmer & S.A. Bonar. 2006 Comparison of upper thermal tolerances of native and nonnative fish species in Arizona. Transactions of the American Fisheries Society 135:1433–1440, 2006
7. Cussac, V.E., D.A. Fernández, S.E. Gómez & H.L. López. 2009. Fishes of southern South America: a story driven by temperature. Fish Physiology and Biochemistry. (Heidelberg, Germany) 35 (1):29-42. [ Links ]
8. Dioni, W. & J.L. Reartes. 1975. Susceptibilidad de algunos peces del Paraná Medio expuestos a temperaturas extremas en condiciones de campo y laboratorio. Physis, Bs.As., sec. B 34 (89): 129-137. [ Links ]
9. Docan, A., V. Cristea & L. Dediu. 2013. Influence of thermal stress on the hematological profile of Oncorhynchus mykiss held in different stocking densities in recirculating aquaculture systems. Lucrari Sttinfice, Seria Zootehnie 55: 262-272. [ Links ]
10. Elliot, J.M. 1981. Some aspects of thermal stress on freshwater teleosts. In: Pickering, AD(ed.). Stress and fish. Academic Press, London. UK. [ Links ]
11. Eme, J. & W. Bennet. 2009. Critical thermal tolerance polygons of tropical marine fish from Indonesia. Journal of Thermal Biology 34: 220-225. [ Links ]
12. Fry, F.E.J. 1971. Effects of environmental factors on the physiology of fish. In: Hoar, W.S. & D.J. Randall (eds.). Fish physiology; vol 6. Academic Press, New York. USA. [ Links ]
13. Gómez, S.E. 1990. Some thermal ecophysiological observations on the catfish Hatcheria macraei (Girard 1855) (Siluriformes, Trichomycteridae). Biota, Osorno, Chile, Vol. 6: 89-95. [ Links ]
14. Gómez, S.E. 1996. Resistenza alla temperatura e alla salinitá in pesci della provincia di Buenos Aires (Argentina), con implicatiozini zoogeografiche. En: Atti Congressuali, 4º Convegno Nazionale. Associazione Italiana di Ittiologia di Acque Dolci (A.I.I.A.D.), Trento, Italia: 171-192. [ Links ]
15. Gómez, S.E., M.J. Gonzalez Naya & L.P. Arribas. 2008. Cambio ambiental en América Austral: pasado, presente y perspectivas futuras. En: "Efecto de los cambios globales sobre biodiversidad". Red 406RT0285 "Efecto de los cambios globales sobre los humedales de Iberoamérica". Eds. L. Fernández Reyes y A. V. Volpedo. Programa CYTED- Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo: Buenos Aires: 47-58. [ Links ]
16. Guzmán, A., & J. Lyons. 2003. Los peces de las aguas continentales del estado de Jalisco, México. Análisis preliminar. - Gnosis [en linea]: Disponible en: http://www.redalyc.org/articulo.oa?id=73000112 [ Links ]
17. Hathaway, E.S. 1927. Quantitative study of the changes produced by aclimatisation in the tolerance of high temperatures by fishes and amphibians. Bulletin of United States Bureau Fisheries 43(2): 169-192. [ Links ]
18. Kilgour, D.M., & R.W McCauley. 1986. Reconciling the two methods of measuring upper lethal temperatures in fishes. Environmental Biology of Fishes, Dordrech 17(4): 281-290. [ Links ]
19. López, H.L., R.C. Menni, R. Ferriz, J. Ponte Gómez & M. Cuello. 2006. Bibliografía de los peces continentales de la Argentina. ProBiota, FCNM, UNLP, Serie Técnico Didáctica 9: 1-165 (ISSN1617-3204). [ Links ]
20. Lutterschimdt, W.I. & V.H. Hutchinson. 1997. The critical thermal maximum: history and critique. Canadian Journal of Zoology 75(10): 1561 – 1574.
21. Menni, R. 2004. Peces y ambientes en la Argentina continental. Monografías del Museo Argentino de Ciencias Naturales No 5, Buenos Aires, 316 pp. [ Links ]
22. Muñoz, N. 2012. Categorías ecológicas de peces en el estuario del Río de La Plata. Tesis de Licenciatura, Univ. Nac. De la Rep. de Uruguay, 75pp. [ Links ]
23. Ortubay, S.G., S.E Gómez & V.E. Cussac. 1997. Lethal temperatures of a Neotropical fish relict in Patagonia, the scale-less characinid Gymnocharacinus bergi Steindachner 1903. Environmental Biology of Fishes, Dordrech, 49 :341-350. [ Links ]
24. Paladino, F.V., J.R. Spotila, J.P. Schubauer & K.T. Kowalski. 1980. The critical thermal maximum: a technique used to elucidate physiological stress and adaptation in fishes. Reviews of Canadian Biology 39: 115-122. [ Links ]
25. Ringuelet, R.A. 1975. Zoogeografía y ecología de los peces de aguas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América del Sur. Ecosur 2(3): 1-122. [ Links ]
Recibido: 29-IV-2014
Aceptado: 7-VIII-2014

