










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
On-line version ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.16 no.2 Ciudad Autónoma de Buenos Aires Dec. 2014
PALEONTOLOGÍA
Selección y uso de los recursos madereros en cazadores-recolectores de la transición Pleistoceno-Holoceno y Holoceno medio, sierras de Tandilia oriental, Argentina
Mariana Brea1, Diana Mazzanti2 & Gustavo A. Martínez2,3
1Laboratorio de Paleobotánica, Centro de Investigaciones Científicas y Transferencia de Tecnología a la Producción (CICyTTP-CONICET), Dr. Materi y España SN, E3105BWA-Diamante, Entre Ríos, Argentina; cidmbrea@gmail.com.
2Laboratorio de Arqueología, Universidad Nacional de Mar del Plata. Funes 3350, 7600-Mar del Plata, Buenos Aires, Argentina; arqueolab@gmail.com.
3Instituto de Geología de Costas y Cuaternario, Universidad Nacional de Mar del Plata. Funes 3350, 7600-Mar del Plata, Buenos Aires, Argentina; gamarti2003@yahoo.com.ar
Resumen
Este trabajo analiza los carbones arqueológicos hallados en seis sitios arqueológicos con cronologías en la transición Pleistoceno–Holoceno (9570 ±150–10465 ± 65 años AP) y Holoceno medio (5089 ± 40–6654 ± 42 años AP) ubicados en las Sierras de Tandilia oriental, provincia de Buenos Aires, Argentina. Los carbones arqueológicos fueron usados para inferir el uso del recurso forestal como un signo de selección humana, la caracterización florística y las condiciones ambientales durante esos períodos. La asociación de carbones identificada para la transición Pleistoceno–Holoceno fue: Celtis tala (Cannabaceae), Salix humboldtiana (Salicaceae), Baccharis sp. y Senecio sp. (Asteraceae). Mientras que para el Holoceno medio fueron registradas: Celtis tala, Baccharis sp., Colletia sp. (Rhamnaceae) y Schinus sp. (Anacardiaceae). Colletia sp., Baccharis sp. y Celtis tala tienen alto poder calórico y larga duración de combustión y fueron una excelente fuente de calor y probablemente fueron usados por los cazadores-recolectores como calefacción, para cocinar y para iluminar. El registro de Celtis tala en tres sitios arqueológicos durante la transición Pleistoceno–Holoceno indica la presencia de bosques xerofíticos al menos desde ca. 10000 años AP en las Sierras de Tandilia oriental. La vegetación se desarrolló bajo un clima frío árido a semi-árido.
Palabras clave: Sierras de Tandilia oriental; Transición Pleistoceno-Holoceno y Holoceno medio; Carbones arqueológicos; Estrategias de selección y uso de recursos forestales; Poblamiento temprano; Cazadores-recolectores.
Abstract
Selection and use of the timber resource in hunter-gatherers in the Pleistocene-Holocene transition and middle Holocene, eastern Tandilia range, Argentina. This paper analyses the anthracological records founded in six archeological sites with chronologies in Pleistocene–Holocene transition (9570 ±150–10,465 ± 65 years BP) and middle Holocene (5089 ± 40–6654 ± 42 years BP) located in the eastern Tandilia range, Buenos Aires province, Argentina. The archaeological charcoals were used to infer the use of forest resources as a sign of human selection, floristic characterization and the environmental conditions during this period. The charcoal assemblages identified for Pleistocene–Holocene transition was: Celtis tala (Cannabaceae), Salix humboldtiana (Salicaceae), Baccharis sp. and Senecio sp. (Asteraceae). While for the middle Holocene were recorded: Celtis tala, Baccharis sp., Colletia sp. (Rhamnaceae) and Schinus sp. (Anacardiaceae). Colletia sp., Baccharis sp. and Celtis tala have high caloric values and long combustible duration and were an excellent source of heat and were probably used by hunter-gatherers for heating, cooking and lighting. The record of Celtis tala in three archaeological sites during the Pleistocene–Holocene transition indicates the presence of xerophytic forests at least from the ca. 10,000 years BP in the eastern Tandilia range. The vegetation developed under a cold arid to semi-arid climate.
Key words: Eastern Tandilia range; Pleistocene–Holocene transition and middle Holocene; Strategies of selection and use of timber; Early settlements; Hunter-gatherers.
INTRODUCCIÓN
Los registros antracológicos no sólo son considerados en los estudios arqueológicos para realizar fechados radiocarbónicos sino que también se analizan la madera y la leña desde el punto de vista anatómico. Los recursos forestales son empleados en un gran número de actividades domésticas y productivas, en la construcción de viviendas y en la producción de artefactos (Piqué i Huerta, 1995, 1999, 2006; Marconetto, 2002, 2007, 2008; Caruso et al., 2008).
El proceso de abastecimiento de los recursos forestales como materia prima para obtener luz, calor y cocción de alimentos de los cazadores-recolectores que habitaron las sierras de Tandilia oriental durante la transición Pleistoceno–Holoceno y Holoceno medio es muy poco conocido (Martínez et al., 2013; Mazzanti et al., 2013). Esta contribución permite formular hipótesis sobre la disponibilidad de los recursos madereros durante ambos períodos así como también, sobre el uso y las necesidades sociales, mediante el reconocimiento de los vegetales quemados intencionalmente. Estos últimos, habrían sido utilizados primariamente como combustibles para cocción de alimentos, calor y/o obtención de luz; función que involucra el aprovisionamiento, traslado, procesamiento y descarte de las especies vegetales en los sitios arqueológicos.
La importancia de la identificación taxonómica de los carbones arqueológicos en los sitios con ocupaciones tempranas tiene valor científico porque permite inferir cuáles fueron los recursos forestales utilizados y evaluar las estrategias de selección y uso de los vegetales por parte de los grupos humanos que ocuparon los sitios arqueológicos. La identificación de especies permite además realizar inferencias paleoclimáticas para los periodos considerados.
Con este propósito se analizaron muestras de carbones arqueológicos recolectados en áreas de combustión y fogones de los cinco sitios arqueológicos con cronologías en la transición Pleistoceno–Holoceno: Cueva Burucuyá, Cueva La Brava, Cueva El Abra, Lobería I sitio 1 y Abrigo Los Pinos, ubicados en reparos rocosos de las Sierras de Tandilia oriental (Figs. 1 y 2). Asimismo y con la finalidad de realizar un análisis comparativo con los carbones del Holoceno medio se analizaron muestras antracológicas de las ocupaciones humanas datadas en ese período que provienen de: Alero El Mirador, Cueva El Abra, Lobería I sitio 1 y Abrigo Los Pinos (Figs. 1 y 2).
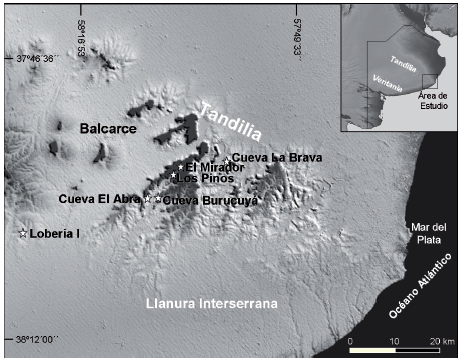
Fig. 1. Ubicación geográfica del área de estudio en las sierras de Tandilia oriental, provincia de Buenos Aires, Argentina donde se observan los sitios arqueológicos estudiados.

Fig. 2. Perfiles estratigráficos de los sitios arqueológicos: Abrigo Los Pinos, Cueva Burucuyá, Cueva La Brava, Cueva El Abra, Alero El Mirador, Lobería 1 sitio 1.
Las características geomorfológicas de esta región, conformada por sierras y serranías distribuidas de forma aislada y separadas por extensos valles interiores, han constituido un entorno muy adecuado para el asentamiento humano durante un periodo en el cual predominaron condiciones climáticas frías, áridas-semiáridas y/o secas como fue la transición Pleistoceno–Holoceno que contrastaron con condiciones en general más cálidas y húmedas del Holoceno.
MATERIALES Y MÉTODOS
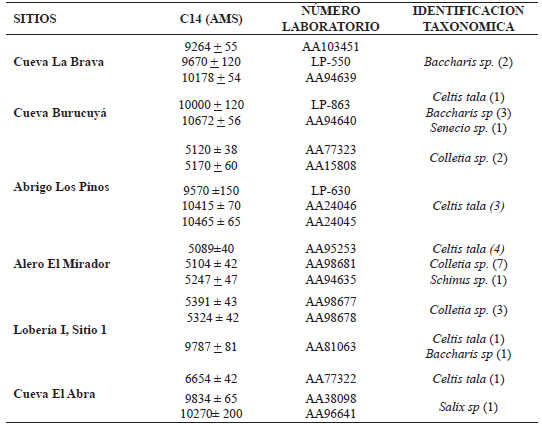
Los materiales recuperados provienen de seis sitios arqueológicos y corresponden a muestras de maderas carbonizadas dispersas en plantas de excavación y otras concentradas en fogones. Estos contextos arqueológicos presentan dataciones de la transición Pleistoceno–Holoceno y Holoceno medio (Tabla 1).
Tabla 1. Dataciones radiocarbónicas de los contextos arqueológicos que conservaron las muestras antracológicas analizadas. El número de ejemplares se muestra entre paréntesis.
Los carbones arqueológicos se estudiaron por fractura a mano en sus tres cortes diagnósticos: corte transversal (CT), longitudinal radial (CLR) y longitudinal tangencial (CLT) y se siguieron los criterios del International Association of Wood Anatomits (IAWA Committee, 1989) para la determinación de los caracteres anatómicos. Se utilizaron claves y descripciones anatómicas para determinar taxonómicamente el material carbonizado (Metcalfe & Chalk, 1950; Carlquist, 1962; Tuset, 1963; Tortorelli, 1956; Tuset & Duran, 1970; Garibotti, 1998; Marconetto, 2008; Roig & Videla, 2006–2009). Se utilizó la base de datos disponible online del InsideWood database (InsideWood 2004–en adelante; Wheeler, 2011) con la finalidad de comparar anatómicamente con material actual. Además, se utilizó la colección de material carbonizado de referencia del Laboratorio de Paleobotánica del CICYTTP-CONICET, Diamante (Zucol et al., 2014). Se realizaron las descripciones anatómicas de los carbones arqueológicos y las asignaciones de los materiales a nivel familiar, genérico y/o específico cuando fue posible. Las microfotografías fueron tomadas con un microscopio estereoscópico Nikon SMZ-1000 y una cámara digital Nikon Coolpix S4. El material antracológico se encuentra depositado en el Laboratorio de Arqueología, Universidad Nacional de Mar del Plata.
Las deformaciones y alteraciones por procesos fisicoquímicos producto de la termoalteración no modifican los caracteres cualitativos que permiten determinar taxonómicamente a los carbones arqueológicos (Scheweingruber, 1988; Solari, 2000). Por este motivo, para la determinación del material carbonizado se utilizaron los caracteres cualitativos, entre los que se destacan anillos de crecimiento, porosidad, disposición y contorno de los elementos de vasos; placas de perforación, tipo de punteaduras intervasculares, tipo de parénquima axial, tipo de radios, fibras y tipos de idioblastos (Couvert, 1970; Rivera & Fernández, 1997–1998; Garibotti, 1998; Pearsall, 2000; Marconetto, 2002; Marconetto et al., 2007).
ASPECTOS GEOMORFOLÓGICOS, ESTRATIGRÁFICOS Y DE VEGETACIÓNDEL ÁREA DE ESTUDIO
El área estudiada corresponde al sector oriental de Tandilia, que juntamente con Ventania constituyen los dos sistemas serranos de la provincia de Buenos Aires (Fig. 1). Tandilia se extiende de las proximidades de Olavarría hasta Mar del Plata, con alturas máximas relativas, en el sector estudiado de 200 msm.
Las características geomorfológicas de la región permitieron un dominio visual amplio sobre el paisaje para los grupos humanos que habitaron estas áreas. Además, la abundancia de fuente de agua dulce y la proximidad con el océano Atlántico ofrecieron potenciales recursos de origen continentales y costero (Martínez et al., 2013; Mazzanti & Bonnat, 2013).
Los sitios se ubican en reparos rocosos constituidos por cuevas, abrigos y aleros de variadas dimensiones, que generalmente se encuentran en distintas posiciones de las vertientes rocosas de los cuerpos serranos y se han formados por procesos de disolución (Martínez, 2007). Las matrices sedimentarias no sobrepasan los 70 cm (Fig. 2), a excepción del sitio Lobería I, cuya secuencia sedimentaria alcanza 1,80 m. Este sitio además, se encuentra en un contexto geomorfológico diferente, ya que se ubica en un afloramiento rocoso de poca expresión areal y altitudinal, en un paisaje de lomadas en la Llanura Interserrana (Mazzanti et al., 2013) (Fig. 1).
Las secuencias sedimentarias abarcan el lapso Pleistoceno tardío–Holoceno y están integradas por sedimentos limo areno-arcillosos, con facies subordinadas de psefitas como depósitos clásticos. Las secuencias son grano-decrecientes con depósitos más gruesos en la base (Pleistoceno tardío) y más finas hacia el Holoceno tardío. Esta disminución de la energía del ambiente se suma a indicadores de mayor humedad como horizontes edáficos y depósitos bioclásticos (diatomitas). El rasgo sedimentario más distintivo y recurrente de las secuencias de los sitios, corresponde la transición Pleistoceno tardío–Holoceno. Está constituido por una discordancia de erosión, que señala un cambio granulométrico, de coloración y grado de compactación de los sedimentos infra y suprayacentes. Se ubica inmediatamente por debajo de los niveles de ocupación temprana (ca. 9400–10500 años AP) y marca la coincidencia en la región con el mejoramiento climático postglacial (Martínez 2007; Martínez et al., 2013). Distintos proxies (estratigráficos, sedimentológicos, pedogenéticos y contenido de diatomeas y fitolitos) sugieren que los sitios de Tandilia oriental evolucionaron desde un ambiente seco hacia uno de mayor humedad durante el Holoceno medio–tardío, reflejando un aumento en el régimen de las precipitaciones regionales (Martínez et al., 1999, 2013; Martínez & Osterrieth, 2004).
La vegetación que caracteriza la región pampeana es la estepa de gramíneas, con una cobertura rala, donde dominan las poáceas, compuestas y leguminosas herbáceas (León, 1991). En el Sudoeste de la provincia de Buenos Aires, esta fisonomía se interrumpe como consecuencia de las variaciones bióticas altitudinales que presentan los cordones de Tandilia y Ventania, los cuales delimitan la denominada Llanura Interserrana (Fig. 1). En las sierras predominan las especies del género Baccharis L., compuestas arbustivas perennes de aproximadamente 1,5 m de altura, asociada con Colletia paradoxa (Spreng.) Escal. (curro), una ramnácea arbustiva subáfila de 1–1,5 m de altura, muy común en suelos arenosos y serranías adaptada a ambientes áridos, especies del género Senecio L., compuestas arbustivas o subarbustivas, perennes distribuidas por lo general entre grietas y zonas rocosas altas y Dodonaea viscosa (L.) Jacq. (chilca), un arbusto de la familia Sapindaceae (Cabrera, 1976). Esta región, posee numerosos endemismos entre los que se encuentran Baccharis tandilense Speg. y Senecio bravensis (Katinas et al., 2007; Alonso et al., 2009).
El área de estudio pertenece al Distrito Pampeano Oriental en donde la vegetación dominante es la estepa graminosa, que alterna con manchones de comunidades de arbustos, siendo también frecuentes los pajonales, espartillales y estepas de Spartina Schreb. Los bosques nativos dominados por Celtis tala Gillies ex Planch. (tala) son muy escasos y la distribución de esta comunidad edáfica está restringida a sitios particulares como los cordones de depósitos calcáreos de conchilla marinas paralelos a la costa del Río de La Plata o en las terrazas del río Salado, siendo las principales comunidades boscosas nativas de la región (Vervoorst, 1967; Goya et al., 1992; Lahitte & Hurrel, 1994; Stupino et al., 2004; Batista et al., 2005). El tala es común en las zonas ribereñas (ríos, arroyos y aguadas) principalmente en los lugares más elevados y médanos antiguos, y se distribuye en una faja estrecha y en manchones boscosos a lo largo de la costa atlántica. Su distribución más austral en la provincia de Buenos Aires es hasta ca. 38° S donde constituye bosques puros en la región de la laguna de Mar Chiquita (Parodi, 1940; Cabrera, 1976). Es una especie caducifolia que sirve de refugio a animales nativos, de fuente alimenticia y nidificación de aves. Celtis tala es un árbol espinoso, de pequeño a mediano porte y mide entre 8 y 12 m de altura.
Los bosques xerófitos denominados talares están conformados por diversas especies arbóreas: Scutia buxifolia Reissek (coronillo), Jodina rhombifolia (Hook. & Arn.) Reissek (sombra de toro), Schinus longifolius (Lindl.) Speg. (molle), Sambucus australis Cham. & Schltdl. (sauco), Phytolacca dioica L. (ombú), Sapium haematospermum Müll. Arg. (curupí) y Colletia spinosissima J.F. Gmelin. (Cabrera, 1976; Pérez Meroni et al., 2010). Las especies que integran estos bosques extrazonales se relacionan estrechamente con los bosques Chaqueños emplazados a más de 1000 km al norte y constituyen la única formación boscosa de la provincia de Buenos Aires (Parodi, 1940; Vervoorst, 1967; Torretta & Basilio, 2009). Actualmente, los talares presentan distintos grados de degradación debido a la expansión de la frontera agrícola, a la tala selectiva de sus árboles para leña, a la actividad ganadera y a la extracción de cochilla que produce el desmonte total del bosque (Parodi, 1940; Goya et al., 1992; Mérida & Athor, 2006; Torretta & Basilio, 2009).
El molle, es un árbol entomófilo, perennifolio de buen porte, de 10 a 20 m de altura, tolerante a la sequía (Lahitte & Hurrell, 1994). Salix humboldtiana Wild. conocida vulgarmente como sauce, es un árbol caducifolio, de 5 a 18 m de altura que se distribuye en lo largo de los riberas de ríos y arroyos conformando asociaciones boscosas casi puras y densas (Lahitte & Hurrell, 1994; Lahitte et al., 1997).
REGISTROS ANTRACOLÓGICOS ENLAS SIERRAS DE TANDILIA ORIENTAL
Del total del material carbonizado recuperado en los sitios arqueológicos fue posible identificar a nivel taxonómico 21 muestras de las 23 analizadas. Muchos otros fragmentos recuperados no pudieron ser identificados por la pobre preservación de los caracteres anatómicos y/o por el tamaño de los fragmentos. La cantidad de carbones arqueológicos con los que se puede estudiar en los sitios arqueológicos de la región pampeana, resulta ser bastante menor en relación a otras regiones (Marconetto, 2008; Andreoni & Caparelli, 2012). La escasa recuperación de maderas carbonizadas probablemente esté relacionada con el medio ambiente donde se preservó el material. A excepción de cueva Tixi, los sitios en cuevas y aleros de Tandilia, analizados en este trabajo, poseen pHs ligeramente ácidos (5,5) (Bonnat & Mazzanti, en prensa), que habrían destruido parcial o totalmente los restos orgánicos. A continuación se describen los carbones identificados en los sitios arqueológicos estudiados en este trabajo:
Familia Cannabaceae
Celtis tala "tala" (Fig. 3, A–D)

Fig. 3. Carbones arqueológicos. A–D, materiales asignados a Celtis tala. A. CT, vista general mostrando vasos, radios y parénquima axial confluente y abundante (Abrigo El Mirador, D4/5-6); B. CT, detalle de vasos solitarios y múltiples radiales, radios y parénquima axial (Abrigo El Mirador, D4/5-6); C. CT, detalle de los vasos y el parénquima axial (Cueva El Abra, D3/a/5); D. CLR, detalle de los radios heterocelulares y los elementos vasales cortos con tabiques oblicuos a rectos (Cueva El Abra, D3/a/5). E. material asignado a Colletia sp. (Abrigo El Mirador, D4/d/3); CT, detalle de los vasos con tendencia a la disposición ulmoide y parénquima axial abundante y confluente. F. material asignado a Schinus sp. (Abrigo El Mirador, C3/4); CT, detalle de los vasos con tendencia a la disposición dendrítica. G. material asignado a Baccharis sp. (Cueva La Brava, G/3/b/3); CT, detalle de los vasos solitarios y múltiples radiales, parénquima axial vasicéntrico y confluente. H–J. material asignado a Senecio sp. (Cueva Burucuyá, C6/a/2); H. CT, vista general; I. CT, detalle de los vasos dispuestos en series múltiples radiales y solitarios, radios multiseriados y parénquima axial escaso y vasicéntrico; J. CLR, detalle de los radios heterocelulares. Escalas gráficas: en A = 500 μm; B–G, I = 250 μm; H = 1000 μm.
Material estudiado: Cueva Burucuyá (C6/c), Lobería I sitio 1 (D8/d/18) y Abrigo Los Pinos (F2/d/10bis, C2/a/13), Cueva El Abra (E3/b/8) y Alero El Mirador (D4/b/3; D4/5-6; C4/c/3) (Tabla 1)
Descripción: en corte transversal, se observan los anillos de crecimiento demarcados por la disposición de los vasos. La porosidad es circular. La disposición de los vasos es en arreglo tangencial. Los vasos son pequeños, vacíos y de contorno circular a elíptico. La mayoría son solitarios y en menor proporción múltiples radiales de 2–3 elementos, siendo muy escasos en cadenas. El parénquima axial es confluente y abundante. Los radios son visibles bajo lupa, son anchos y de recorrido rectilíneo. En corte longitudinal tangencial, los radios son uniseriados y multiseriados de 3–4 células de ancho. Los elementos de vaso son cortos y con tabiques oblicuos a rectos. Las placas de perforación son simples y las punteaduras intervasculares son areoladas y alternas. En corte longitudinal radial, los radios son heterocelulares constituidos por células procumbentes en el cuerpo y 1–2 hileras de células cúbicas, la altura de los radios en número de células es de 4–18.
Observaciones: carbones asignados a Celtis tala fueron datados en la transición Pleistoceno–Holoceno y Holoceno medio (Tabla 1).
Familia Salicaceae
Salix humboldtiana "sauce" (Fig. 4, A–C).
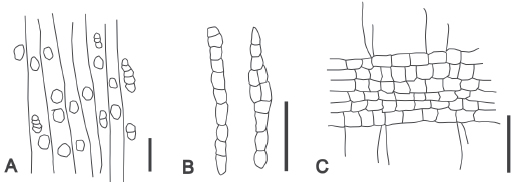
Fig. 4. Esquemas de los caracteres anatómicos de Salix humboldtiana (Cueva El Abra, E4/a/17). A. CT, vasos predominantemente solitarios y múltiples radiales, radios delgados de trayecto rectilíneo y parénquima axial no visible bajo lupa. B. CLT. radios uniseriados y parcialmente biseriados. C. CLR. Radios homogéneos constituidos por células procumbentes y raras cúbicas. Escala gráfica: 500 μm.
Material estudiado: Cueva El Abra (E4/a/17) (Tabla 1).
Descripción: en corte transversal, los vasos tienen contorno hexagonal y la porosidad es difusa. Se presentan predominantemente solitarios, raramente múltiples radiales y agrupados. Vasos de contorno hexagonal. Los radios son muy delgados y cortos. El parénquima axial es muy escaso y no visible bajo lupa, probablemente apotraqueal. En corte longitudinal tangencial, los radios son uniseriados, parcialmente biseriados y biseriados. En corte longitudinal radial, los radios son homogéneos constituidos por células procumbentes y raras cúbicas. La altura de los radios en número de células es de 7–27.
Obervaciones: el área de combustión muestreada corresponde a las actividad humana más temprana del sitio (Tabla 1).
Familia Rhamnaceae
Colletia sp. "curro" (Fig. 3, E).
Material estudiado: Alero El Mirador (D4/d/3; D4/a/4), Abrigo Los Pinos (C2/B/5) y Lobería I sitio 1 (D3/c/17, sondeo 1/17 y 18) (Tabla 1).
Descripción: en corte transversal, se observan anillos de crecimiento demarcados, la porosidad es difusa con tendencia a semicircular. El arreglo de los elementos vasales tiene una tendencia a la disposición ulmoide. Los vasos son pequeños y de contorno angular a casi hexagonal. Los vasos se disponen mayoritariamente solitarios y múltiples radiales de 2 elementos, también se observan múltiples radiales de 5–12 elementos siendo muy raros los agrupados. El parénquima axial es confluente y abundante. Los radios son anchos y angostos. En corte longitudinal tangencial, los elementos de vasos tienen punteaduras intervasculares areoladas y alternas con tabiques oblicuos. Los radios son abundantes, uniseriados biseriados y multiseriados. En corte longitudinal radial, los radios son heterogéneos conformados por células procumbentes y verticales. La altura de los radios en número de células es de 4–13.
Observaciones: los carbones identificados corresponden a ocupaciones humanas datadas en el Holoceno medio (Tabla 1).
Familia Anacardiaceae
Schinus sp. "molle" (Fig. 3, F).
Material estudiado: Alero El Mirador (C3/4) (Tabla 1).
Descripción: En corte transversal, anillos de crecimiento demarcados. Porosidad difusa a semicircular. Vasos con tendencia a la disposición dendrítica, con patrones diagonales y/o radiales. Vasos de contorno angular. Vasos agrupados, múltiples radiales y solitarios. Parénquima axial paratraqueal escaso. Radios delgados, bajos y poco visibles bajo lupa. En corte longitudinal tangencial, los elementos vasculares con trayecto rectilíneo. Radios uniseriados, biseriados y triseriados. En corte longitudinal radial, los radios son heterogéneos constituidos por células procumbentes y cúbicas y/o verticales.
Observaciones: estos carbones asignados a Schinus sp. corresponden a la ocupación humana del Holoceno medio (Tabla 1).
Familia Asteraceae
Baccharis sp. "chilca" (Fig. 3, G)
Material estudiado: Cueva La Brava (G/3/b/3), Cueva Burucuyá (C7/b/5; C8/a/4; D7/a/5), Sitio Lobería I sitio 1 (D4/c/18) (Tabla 1).
Descripción: en corte transversal, la porosidad es semicircular y los vasos se disponen en arreglo diagonal. Los vasos son medianos y con contorno angular. Se presentan solitarios, múltiples radiales cortos de 2–3 elementos y agrupados. El parénquima axial es vasicéntrico y confluente. Los radios son muy visibles bajo lupa, y se observan claramente que son de dos tipos, anchos y angostos. Los mismos tienen recorrido rectilíneo en sección transversal. En corte longitudinal tangencial, los radios son uniseriados y fusiformes. En corte longitudinal radial, los radios son de dos tipos: homogéneos, constituidos exclusivamente por células procumbentes y heterogéneos constituidos por células procumbentes y cuadradas.
Observaciones: las muestras provenientes de los dos primeros sitios se recolectaron dentro de fogones en cubetas datados en la transición Pleistoceno–Holoceno y el tercero corresponde a dataciones del Holoceno medio (Tabla 1).
Género Senecio sp. (Fig. 3, H–J).
Material estudiado: Cueva Burucuyá (C6/a/2) (Tabla 1).
Descripción: En corte transversal, los anillos de crecimiento están demarcados por las diferencias en el diámetro de los vasos, diámetro de fibras y por bandas de parénquima axial. La porosidad es semicircular. Los vasos están dispuestos en series múltiples radiales y también se presentan solitarios. El parénquima axial es escaso, paratraqueal vasicéntrico y bandeado. Los radios son visibles bajo lupa y muy anchos. En corte longitudinal tangencial, los radios son multiseriados y raramente biseriados, el ancho de los radios varía en número de células entre 2–14. En corte longitudinal radial, los radios son heterogéneos constituidos por células procumbentes y cúbicas. Estos materiales corresponden al contexto datado en la transición Pleistoceno–Holoceno (Tabla 1).
CONSIDERACIONES FINALES
El análisis antracológico permitió identificar taxonómicamente muestras provenientes de carbones dispersos y de fogones de contextos arqueológicos, los cuales aportan conocimientos novedosos sobre los recursos forestales existentes en el ambiente de Tandilia oriental. Estos recursos han sido utilizados por los cazadores-recolectores de la región al menos durante la transición Pleistoceno-Holoceno y el Holoceno medio.
Las especies identificadas para la transición Pleistoceno–Holoceno fueron: Celtis tala (Cannabaceae), Salix humboldtiana (Salicaceae), Baccharis sp. y Senecio sp. (Asteraceae) y abarcan el lapso 9570 ±150–10672 ±56 + años AP (Tabla 1). Mientras que para el Holoceno medio se registraron Celtis tala, Baccharis sp., Colletia sp. (Rhamnaceae) y Schinus sp. (Anacardiaceae), para el lapso 5089 ± 40–6654 ± 42 años AP (Tabla 1).
El recurso maderero utilizado como combustión para fogones por los cazadores-recolectores incluía subarbustos, arbustos y árboles. Colletia, Baccharis y Senecio sp. son géneros muy abundantes en la región estudiada con diversas especies endémicas. Los carbones asignados a Senecio poseen muchas semejanzas con S. subulatus, D. Don ex Hook & Arn. que es la única especie del género descripta xilológicamante para Argentina (Roig & Videla, 2006–2009) aunque también comparte muchos caracteres anatómicos con las especies de Senecio descriptas por Carlquist (1962), motivo por el cual los carbones arqueológicos fueron asignados a este género.
Senecio subulatus es una especie arbustiva perenne, presente en suelos arenosos de las regiones áridas del centro de Argentina (provincia del Monte), desde Mendoza al sur de Buenos Aires y Río Negro (Roig y Videla, 2006–2009) mientras que Senecio bravensis, una especie endémica de las sierras bonaerenses (Katinas et al., 2007) es un arbusto perenne de poco porte subaéreo que habita zonas rocosas y su hábito camefítico, le permite soportar las estaciones desfavorables. El registro de este taxón refuerzan la postura junto con la presencia de Celtis tala y Baccharis sp., de que durante el lapso Pleistoceno–Holoceno la vegetación estaba adaptada a condiciones xerofíticas y probablemente más frías que en la actualidad. Existe un consenso generalizado con respecto a las condiciones paleoclimáticas para la transición Pleistoceno final en la región pampeana. Numerosos proxies señalan que entre los ca. 12000–10450 años AP las condiciones climáticas fueron frías, áridas-semiáridas y/o secas con condiciones locales de humedad (Prieto, 2000; Zarate et al., 2000; Tonni et al., 2003; Quattrocchio et al., 2008; Tonello & Prieto, 2010; Martínez et al., 2013).
Salix humboldtiana fue hallada en el sitio Cueva El Abra en niveles asignados a la transición Pleistoceno–Holoceno. Este taxón fue parte del paisaje ribereño integrando los escasos bosques en galería característicos de esta región. S. humboldtiana es muy común en ríos y arroyos, donde los pulsos de inundación son el factor clave para el establecimiento de los bosques de sauces (Liotta, 2001). Los sauzales viven en regiones subhúmedas y semiáridas bajo un clima templado a templado-cálido, donde los suelos se caracterizan por una buena disponibilidad de agua, ser arenosos y con buen drenaje. La presencia de S. humboldtiana en el sitio Cueva El Abra, localizado sobre el valle del arroyo La Vigilancia, estaría relacionado con la presencia de un pequeño arroyo que pierde su caudal durante eventos de sequía. Los bosques en galería que caracterizan la vegetación en estos valles de sierras bajas habrían requerido un régimen de precipitación suficiente para mantener el caudal permanente de este arroyo. Así, la presencia de carbones arqueológicos de S. humboldtiana indicarían un régimen de precipitación más húmedo que se habría establecido aproximadamente a los 10000 años AP. Martínez et al. (2013), consideran a partir de diferentes proxies de los sitios de Tandilia Oriental, que hacia los 10500 años AP, las condiciones de aridez-semiaridez, cambiaron a un régimen de mayor pluviosidad y temperatura. Este cambio (entre 11500–9000 años AP) ha sido reportado en numerosos registros Proxy de la región pampeana (Prado et al., 1987; Alberdi et al., 1989; Tonni & Cione,1994; Zarate et al., 1995, 2000; Prieto, 1996; Zárate, 1997; Borrero et al., 1998; Bonadonna et al., 1999; Martínez, 1999, 2001, 2007; Tonni et al., 1999; Prieto et al., 2009; Grill et al., 2007; Gutiérrez & Martínez, 2008; Quattrocchio et al., 2008.;Tonello & Prieto 2010; Gutiérrez et al., 2011). Así, las poblaciones humanas que habrían arribado a la región alrededor de ca. 11000–10500 años AP, vivieron bajo condiciones más frías y subhúmedas-secas, que cambiaron a más húmedas y de mayor temperatura ca. 10500 años AP (Borrero et al. 1998, Martínez, 2001, Politis et al. 2004, Martínez et al. 2013).
El hallazgo de estructuras de combustión conteniendo carbones de Celtis tala en los si tios arqueológicos Cueva Burucuyá, Lobería I sitio 1 y Abrigo Los Pinos confirma la presencia de bosques xerofíticos durante la transición Pleistoceno–Holoceno, habiéndose registrado también para el Holoceno medio (Abrigo El Mirador y El Abra). Esto indica la presencia de estos bosques al menos desde los ca. 10000 años AP en las sierras de Tandilia oriental. Los registros palinológicos más antiguos de Celtis fueron hallados en el sitio arqueológico Paso Otero 4 (Districto de Necochea, Buenos Aires) en sedimentos del Miembro Guerrero asociados al paleosuelo Puesto Callejón Viejo con edades que oscilan entre los ca. 10400 años AP y los 9400 años AP (Grill et al., 2007). Mientras, que en el sitio arqueológico Paso Otero 5 se registran granos de polen y cistolitos de Celtis sp. desde los ca. 8900 años AP (Gutiérrez et al., 2011). Cistolitos de Celtis sp. fueron hallados en el sitio arqueológico Amalia (Partido de Balcarce, Buenos Aires) en sedimentos con cronologías entre ca. 1676–7700 años AP (Zucol et al., 2007).
Para el Holoceno medio la presencia de carbones arqueológicos de Celtis tala, Baccharis sp., Colletia sp. (Rhamnaceae) y Schinus sp. (Anacardiaceae), para el lapso 5089 ± 40–6654 ± 42, podrían sugerir condiciones semiáridas. Este lapso se corresponde, según diferentes registros proxies del sudeste de la región pampeana, con el cambio de las condiciones mas cálidas y húmedas del período Hypsitermal (ca. 8000–6000 años AP) (Aguirre & Whatley, 1995; Bonadonna et al., 1999; Prieto, 1996; Prado et al. 1987; Tonni et al. 1999; Grill et al, 2007; Hassan et al., 2009), a un retorno hacia condiciones más áridas (ca. 5400–4500 años AP): (Aguirre, 1995; Bonadonna et al., 1999; Prieto, 1996; Tonni et al., 1999; Figini et al., 1985; Zárate & Flegenheimer, 1991; Zárate & Blasi 1993; Zarate et al. 2000; Martínez, 2001; Cavallotto et al., 2004; Stutz et al., 2006; 2010; Vilanova et al., 2006). No obstante, estas oscilaciones climáticas de menor magnitud del Holoceno, podrían no verse reflejadas en variaciones de las especies consideradas, ya que el ambiente serrano, a diferencia de otros ambientes de la llanura pampeana, los recursos de agua superficial y subsuperficial (Martínez et al., 2013; Mazzanti & Bonnat, 2013) se podrían haber mantenido sin mayores variaciones frente a dichas oscilaciones.
Los recursos forestales seleccionados y transportados a los sitios arqueológicos debieron solucionar necesidades económico-sociales de los grupos humanos al ser utilizados como recursos energéticos para producir fuente de calor y luz. Celtis tala y Schinus poseen leñas duras con alto valor calórico por lo que muy probablemente se hayan seleccionado como combustible. La característica calórica del tala es su poca llama pero abundantes brasas lo que hace a este recurso material de gran valor energético (Tortorelli, 1956; López, 2006). Por otro lado, Salix humboldtiana posee una madera blanda que se utiliza como iniciador o avivador del fuego; hace mucha llama pero su duración es breve y de bajo poder calórico, no produce brasas sino cenizas. La selección de diversas especies con diferentes propiedades de transferencia de energía favoreció la optimización del uso del fuego. La leña blanda se habría utilizado para iniciar y/o avivar el fuego, produciendo mucha llama pero de poca duración. Mientras que la leña dura habría favorecido la producción de brasas y cenizas para la manutención del fuego en los fogones utilizados para cocinar, obtener calor y/o luz. La combinación de tallos de Senecio con leña blanda y dura (tala, curro y chilca) evidencia el encendido y el mantenimiento de las estructuras de combustión (Tortorelli, 1956; Demaio et al., 2002; López, 2006; Brea et al., 2013).
Colletia sp. y Baccharis sp. son dos arbustos comunes del ámbito serrano, que se utilizan en la actualidad como material de combustión debido a que tienen alto poder calórico. La utilización de estos recursos por parte de los cazadores-recolectores permite inferir que la exploración maderera se realizó en el entorno de los sitios, sin necesidad de ampliar el área de captación.
AGRADECIMIENTOS
Este trabajo fue financiado por la ANPCyT, PICT2008-Nº 1390, PICT2013-Nº 1979 y por la Universidad Nacional de Mar del Plata HUM390/13. Los autores agradecen a M.L. Pochettino y a un revisor anónimo por sus sugerencias y comentarios que permitieron mejorar esta contribución.
BIBLIOGRAFÍA
1. Aguirre, M.L. 1995. Cambios ambientales en la región costanera bonaerense durante el Cuaternario tardío. Evidencias malacológicas. IV Jornadas de Geología Bonaerenses, Actas 1: 35–45.
2. Aguirre, M.L. & R.C. Whatley. 1995. Late Quaternary marginal marine deposits and paleoenvironments from northeastern Buenos Aires province: a review. Quaternary Science Reviews 14: 223–254.
3. Alberdi, T.A., J.L. Menégaz, J.L. Prado & E.P. Tonni. 1989. La fauna local Quequén Salado-Indio Rico (Pleistoceno tardío) de la Provincia de Buenos Aires, Argentina. Aspectos paleoambientales y bioestratigráficos. Ameghiniana 25: 225–236.
4. Alonso, S.I., I.R. Guma, M.C. Nuciari, M.C. & A.van Olphen. 2009. Flora de un área de la Sierra La Barrosa (Balcarce) y fenología de especies con potencial ornamental. Revista de la Facultad de Ciencias Agronómicas, Universidad Nacional de Cuyo 41: 23–44.
5. Andreoni, D. & A. Caparelli. 2012. El ser humano y la leña en la cordillera de Mendoza (Argentina) a lo largo del Holoceno: Sitio arqueológico Arroyo Malo 3. Magallania 40: 203–228.
6. Batista, W.B., M.A. Taboada, R.S. Lavado, S.B. Perelman & R.J.C. León. 2005. Asociación entre comunidades vegetales y suelos en el pastizal de la Pampa Deprimida. En: M. Oesterheld, M.RAguiar, C.M. Ghersa & J.M. Paruelo (Eds.), La heterogeneidad de la vegetación de los agroecosistemas. Un homenaje a Rolando León, pp 113–129, Editorial Facultad de Agronomía, Buenos Aires.
7. Borrero, L.A., M. Zárate, L. Miotti & M. Massone. 1998. The Pleistocene-Holocene transition and human occupations in the southern cone of South America. Quaternary International 49/50: 191–199.
8. Bonadonna, F.P., G. Leone, & G. Zanchetta. 1999. Stable isotope analyses on the last 30 ka molluscan fauna from Pampa grassland, Bonaerense region, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 153: 289–308.
9. Bonnat, G & D. Mazzanti. en prensa. Análisis de la tecnología lítica de las ocupaciones humanas efímeras durante la transición Pleistoceno-Holoceno: el caso de Cueva La Brava (Buenos Aires, Argentina). Intersecciones en Antropología (en prensa). [ Links ]
10. Brea, M., M.J. Franco, M. Bonomo & G. Politis. 2013. Análisis antracológico preliminar del sitio arqueológico Los Tres Cerros 1 (Delta Superior del río Paraná), provincia de Entre Ríos. Revista del Museo de La Plata, Sección Antropología 13 (87): 346–360.
11. Cabrera, A.L. 1976. Regiones fitogeográficas Argentinas. Enciclopedia Argentina de Agricultura y Jardinería. Fascículo 1, Tomo 2. Editorial ACME, Buenos Aires, 85 pp. [ Links ]
12. Carlquist, S. 1962. Wood anatomy of Senecioneae (Compositae). Aliso 5: 123–146.
13. Caruso, L., M.E. Mansur & R. Piqué. 2008. Voces en el bosque: El uso de recursos vegetales entre cazadores-recolectores de la zona central de Tierra del Fuego. Darwiniana 46: 202–212.
14. Cavallotto, J.L., R.A. Violante & G. Parker. 2004. Sea-level fluctuations during the last 8600 years in the La Plata river (Argentina). Journal of Coastal Research 8: 125–133.
15. Couvert, M. 1970. Etude des charbons prehistoriques. Preparation des coupes minces et analyse des structures. Travaux du Centre de recherches. Anthropologiques Préhistoriques et Ethnographiques 5: 5–88.
16. Demaio, P., U.O. Karlin & M. Medina. 2002. Árboles nativos del centro de Argentina. L.O.L.A. Buenos Aires, 210 pp. [ Links ]
17. Figini, A.A., R.A. Huarte, J.E. Carbonari, G.J. Gomez, A.C. Zubiaga, E.Tonni & F. Fidalgo. 1985. Edad isotópica de carbonatos de la cueva Tixi, partido de General Alvarado, provincia de Buenos Aires. Primeras Jornadas Geológicas Bonaerenses, Actas: 128–130.
18. Garibotti, I.A. 1998. Análisis de la estructura anatómica de carbones arqueológicos de sitios Incaicos (ca. 1480–1530 d.C.) del Valle de Uspallata (Mendoza, Argentina). Boletín de la Sociedad Argentina de Botánica 33: 195–205.
19. Goya, J., G. Placci, M. Arturi & A. Brown. 1992. Distribución y características estructurales de los Talares de la Reserva de Biosfera "Parque costero del sur". Revista de la Facultad de Agronomía (La Plata) 68: 53–64.
20. Grill, S., A. Borromei, G. Martínez, M.A. Gutiérrez, M.E. Cornou & D. Olivera. 2007. Palynofacial analysis in alkaline soils and paleoenvironmental implications: The Paso Otero 5 archaeological site (Necochea district, Buenos Aires province, Argentina). Journal of South American Earth Sciences 24: 34–47.
21. Gutiérrez, M.A. & G. Martínez. 2008. Trends in the faunal human exploitation during the late Pleistocene and early Holocene in the Pampean region (Argentina). Quaternary International 191: 53–68.
22. Gutiérrez, M.A., G. Martínez, H. Luchsinger, S. Grill, A.F. Zucol, G.S. Hassan, M.P. Barros, C.A. Kaufmann & M.C. Álvarez. 2011. Paleoenvironments in the Paso Otero locality during Late PleistoceneeHolocene (Pampean region, Argentina): An interdisciplinary approach. Quaternary International 245: 37–47.
23. Hassan, G.S., E. Tietze, & C.G. De Francesco. 2009. Modern diatom assemblages in surface sediments from shallow lakes and streams in southern Pampas (Argentina): palaeoenvironmental implications. Aquatic Sciences 71: 1015–1621.
24. IAWA Committee. 1989. IAWA list of microscopic features for hardwood identification. IAWA Bulletin n.s. 10: 219–332.
25. InsideWood. 2004–en adelante. InsideWood database http://insidewood.lib.ncsu.edu/search [10/07/2014].
26. Katinas, L., D.G. Gutiérrez, M.A. Grossi & J.V. Crisci. 2007. Panorama de la familia Asteraceae (= Compositae) en la República Argentina. Boletín de la Sociedad Argentina de Botánica 42: 113–129.
27. Lahitte, H. B. & J.A. Hurrell. 1994. Los Árboles de la Isla Martín García árboles y arbustos arborescentes (nativos y naturalizados) de la Reserva Natural y Cultural Isla Martín García. La Plata: Comisión de Investigación Científicas, La Plata (CIC), 200 pp. [ Links ]
28. Lahitte, H.B., J.A. Hurrell, M.J. Belgrano, L.S. Jankowski, K. Mehltreter, M.P. Haloua & G. Canda. 1997. Plantas de la Costa. Las plantas nativas y naturalizadas más comunes de las costas del Delta del Paraná, Isla Martín García y Ribera Platense. L.O.L.A., Buenos Aires, 200 pp. [ Links ]
29. León, R.J.C. 1991. Geographic limits of the region, Geomorphology and geology, Regional subdivisions, Floristic aspects, Description of the vegetation. En: R.T. Coupland (Ed.). Natural Grasslands: Introduction and Western Hemisphere. pp. 369–387, Elsevier, Amsterdam.
30. Liotta, J. 2001. Rasgos biológicos de Salix humboltiana Wild. y régimen de pulsos de inundación. Interciencia 26: 397–403.
31. López, L. 2006. Uso de recursos combustibles madereros en pampas de altura: Los casos Río Yuspe 11 y Río Yuspe 14 (Córdoba). X Congreso Nacional de Estudiantes de Arqueología, Mendoza,16 pp. [ Links ]
32. Marconetto, M.B. 2002. Análisis de los vestigios de combustión de los sitios Alero Don Santiago y Campo Moncada. En: Pérez de Micou, C. (Ed.), Plantas y cazadores en Patagonia, pp. 33–53, Universidad de Buenos Aires, Buenos Aires.
33. Marconetto, M.B. 2007. Aportes de la antracología a la cronología del Valle Ambato. En: M.B. Marconetto, P. Babot & N.Oliszewski (Eds.). Paleoetnobotánica del Cono Sur: Estudios de casos y propuestas metodológicas, pp. 197–218, Ferreyra Editor, Córdoba.
34. Marconetto, M.B. 2008. Recursos forestales y el proceso de diferenciación social en tiempos Prehispanicos en el Valle de Ambato, Catamarca, Argentina. South American Achaeology series 3: 1–207. BARInternational Series.
35. Marconetto, M. B., P. Babot & N. Oliszewski (Eds.). 2007. Paleoetnobotánica del Cono Sur: Estudios de casos y propuestas metodológicas. Ferreyra Editor, Córdoba, 220 pp. [ Links ]
36. Martínez, G., 1999. [Tecnología, subsistencia y asentamiento en el curso medio del Río Quequén Grande: Un enfoque arqueológico. Tesis Doctoral inédita, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, 185 pp. [ Links ]
37. Martínez, G. 2001. Archaelogical research in Paso Otero 5 site. "Fish-tail" projectile points and megamammals in the Pampean region of Argentina. Antiquity 75: 523–528.
38. Martínez, G.A. 2007. Procesos de formación de sitios en reparos rocosos de Tandilia. Cazadores-Recolectores del Cono Sur. Revista de Arqueología 3: 105–127.
39. Martínez, G.A., M. Osterrieth & D.L. Mazzanti. 1999. Estratigrafía de sitios arqueológicos en reparos rocosos en las sierras de La Vigilancia y Valdés, sistema de Tandilia, provincia de Buenos Aires. XII Congreso Nacional Arqueología Argentina, La Plata, Actas III, pp. 139–144.
40. Martínez, G. & M. Osterrieth. 2004. The Pleistocene-Holocene stratigraphical record from early archaeological sites in caves and rockshelters of eastern Tandilia, Pampean Region, Argentina. En: L. Miotti, M. Salemme & N, Flegenheimer (Eds.), Ancient evidences for paleo south americans: From where the south winds blow, pp. 63–68, Center for the Studies of the First Americans (CSFA) and Texas A&M, University Press.
41. Martínez, G.A., D.L. Mazzanti, C. Quintana, A.F. Zucol, M.M. Colobig, G.S., Hassan, M. Brea & E. Passeggi. 2013. Geoarchaeological and Paleoenvironmental context of the human settlement in the Eastern Tandilia Range, Argentina. Quaternary International 299: 23–37.
42. Mazzanti, D.L. & G.F. Bonnat. 2013. Paisajes arqueológicos y cazadores-recolectores de la transición Pleistoceno-Holoceno. Análisis de las cuencas de ocupación en Tandilia oriental, provincia de Buenos Aires, Argentina. Relaciones de la Sociedad Argentina de Antropología 38: 521–541.
43. Mazzanti D.L., G. Martínez, M. de los M. Colobig, A.F. Zucol, E. Passeggi, M. Brea, G.F. Bonnat, G. Hassan, J.L. Soria, J.A. Vera & C.A. Quintana. 2013. Avances en los estudios arqueologicos, geoarqueologicos y paleoambientales en las Sierras Orientales de Tandilia. Resultados preliminares de los sitios Alero El Mirador y Abrigo La Grieta. Revista del Museo de La Plata, Sección Antropología 13 (87): 59–76.
44. Metcalfe, C.R. & L. Chalk. 1950. Anatomy of the Dicotiledons, 2 vol. Claredon Press, Oxford, 724 pp. [ Links ]
45. Mérida, E. & J. Athor. 2006. Talares bonaerenses y su conservación. Fundación de Historia Natural Félix de Azara, Buenos Aires, 259 pp. [ Links ]
46. Parodi, L. 1940. La distribución geográfica de los talares de la provincia de Buenos Aires. Darwiniana 4: 33–56.
47. Pearsall, D.M. 2000. Paleoethnobotany. A handbook of procedures. Academia Press, San Diego, 700 pp. [ Links ]
48. Pérez Meroni, M., M.C. Paleo, M.L. Pochettino & V.S. Lema. 2010. Procesamiento y consumo de vegetales por grupos cazadores-recolectores del Holoceno tardío en los Partidos de Magdalena y Punta Indio, provincia de Buenos Aires. En: M. Berón, L. Luna, M. Bonomo, C. Montalvo, C. Aranda & M. Carrera Aizpitarte (Eds.), Mamül Mapu: pasado y presente desde la arqueología pampeana. Tomo I, pp. 215–226, Editorial Libros del Espinillo (Ayacucho, Pcia. de Buenos Aires).
49. Piqué i Huerta, R. 1995. La gestión de los recursos forestales en Tunel VII. En: J. Estévez Escalera & A. Vila Mitra (Coords.): Encuentros en los concheros fueguinos, pp. 194–238, Treballs d´Etnoarqueologia 1, CSIC, Barcelona.
50. Pique í Huerta, R. 1999. Producción y uso de combustible vegetal arqueológico: Una evaluación arqueológica. Treballs d`Etnoarqueología Nº 3, 307 pp, Universidad Autónoma de Barcelona. Madrid. [ Links ]
51. Piqué i Huerta, R. 2006. Los carbones y las maderas de contextos arqueológicos y el paleoambiente. Ecosistemas 15: 31–38.
52. Politis, G., P. Messineo & C. Kauffman. 2004. El poblamiento temprano de las llanuras pampeanas de Argentina y Uruguay. Complutum 15: 207–224.
53. Prado, J.L., A. Menégaz, E.P. Tonni & M. Saleme. 1987. Los mamíferos de la fauna local Paso Otero (Pleistoceno tardío), provincia de Buenos Aires. Aspectos paleoambientales y bioestratigráficos. Ameghiniana 24: 217–233.
54. Prieto, A.R. 1996. Late quaternary vegetational and climatic changes in the pampa grassland of Argentina. Quaternary Research 45: 73–88.
55. Prieto, A.R. 2000. Vegetational history of the Late glacial–Holocene transition in the grasslands of eastern Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 157: 167–188.
56. Prieto, A.R., A.M. Blasi, C.G. De Francesco & C. Fernández. 2004. Environmental history since 11,000 14Cyr BP of the northeastern Pampas, Argentina, from alluvial sequences of the Luján River. Quaternary Research 62: 146–161.
57. Prieto, A.R., I. Vilanova, M.S. Tonello & S. Stutz. 2009. Reconstrucción de la vegetación y del clima de los pastizales pampeanos durante el Pleistoceno tardío – Holoceno a través del análisis palinológico. Quaternário do Rio Grande do Sul: integrando conhecimentos. Monografias da Sociedade Brasileira de Paleontologia, pp.107–120.
58. Quattrocchio, M.E., A.M. Borromei, C.M. Deschamps, S. Grill & C. Zabala. 2008. Landscape evolution and climate changes in the late Pleistocene-Holocene southern Pampa (Argentina): evidence from palynology, mammals and sedimentology. Quaternary International 181: 123–138.
59. Rivera, S. M. & V.M. Fernández. 1997–1998. Identificación de material leñoso y otros vestigios macrovegetalesarqueológicos de la cueva Epullén Grande, provincia de Neuquén. Algunos aspectos metodológicos y culturales. Paleoetnologica 9: 33–48.
60. Roig, F.A. & E. Videla. 2006–2009. Anatomía de la madera de arbustos de montaña del NW de Mendoza, Argentina. Xama 19–23: 157–238.
61. Scheweingruber, F.H. 1988. Tree-ring, Basics and Applications of Dendrochronology. D. Reidel Publishing Company, Boston, 273 pp. [ Links ]
62. Solari, M.E. 2000. Antracología, modo de empleo: en torno a paisajes, maderas y fogones. Revista Austral de Ciencias Sociales 4: 167–174.
63. Stupino, S.A., M.F. Arturi & J.L. Frangi. 2004. Estructura del paisaje y conservación de los bosques de Celtis tala Gill ex Planch del NE de la provincia de Buenos Aires. Revista de la Facultad de Agronomía, La Plata 105: 37–47.
64. Stutz, S., A.R. Prieto & F. Isla. 2006. Holocene evolution of the Mar Chiquita coastal lagoon area (Argentina) indicated by pollen analsis. Journal of Quaternary Science 21: 17–28.
65. Stutz, S., C.M. Borel, S.L. Fontana, L. del Puerto, H. Inda, F. García-Rodriguez & M.S. Tonello. 2010. Late Holocene climate and environment of the SE Pampa grasslands, Argentina, inferred from biological indicators in shallow, freshwater Lake Nahuel Rucá. Journal of Paleolimnology 44: 761–775.
66. Tonello, M.S. & A.R. Prieto. 2010. Tendencias climáticas para los pastizales pampeanos durante el Pleistoceno tardío-Holoceno: estimaciones cuantitativas basadas en secuencias polínicas fósiles. Ameghiniana 47: 501–514.
67. Tonni, E.P. & A.L. Cione. 1994. Los mamíferos y el clima en el Pleistoceno y Holoceno de la provincia de Buenos Aires. Jornadas Arqueológicas Interdisciplinarias, Programa de estudios Prehistóricos, pp. 127–142.
68. Tonni, E.P., A.L. Cione & A. Figini. 1999. Predominante of arid climates indicated by mammals in the pampas of Argentina during the late Pleistocene and Holocene. Palaeogeography, Palaeoclimatology, Palaeoecology 147: 247–281.
69. Tonni, E.P., R.A. Huarte, J.E. Carbonari & A.J. Figini. 2003. New radiocarbon chronology for the Guerrero Member of the Luján Formation (Buenos Aires, Argentina): palaeoclimatic significance. Quaternary International 109–110: 45–48.
70. Torretta, J.P. & A.M. Basilio. 2009. Dispersión polínica y éxito reproductivo de cuatro especies arbóreas de un bosque xerófito de Argentina. Revista de Biología Tropical (International Journal of Tropical Biology and Conservation) 57: 283–292.
71. Tortorelli, L.A. 1956. Maderas y Bosques Argentinos. Editorial ACME, Buenos Aires, 910 pp. [ Links ]
72. Tuset, R. 1963. Descripción y clave macroscópica de maderas indígenas del Uruguay. Silvicultura 19: 5–55.
73. Tuset, R. & F. Duran. 1970. Descripción y clave macroscópica de maderas comerciales en Uruguay. Facultad de Agronomía, Universidad de la República Boletín 114: 1–63.
74. Vervoorst, F.B. 1967. Las comunidades vegetales de la Depresión del Salado. La Vegetación de la República Argentina. Serie Fitogeográfica N° 7. Buenos Aires INTA. 259 p.p. [ Links ]
75. Vilanova, I., A.R. Prieto & S. Stutz. 2006. Historia de la vegetación en relación con la evolución geomorfológica de las llanuras costeras del este de la Provincia de Buenos Aires durante el Holoceno. Ameghiniana 43: 147–159.
76. Wheeler, E. 2011. InsideWood – A web resource for hardwood anatomy. IAWA Journal 32: 199–211.
77. Zárate, M., 1997. Late Pleistocene geoarchaeology of the southern pampas, Buenos Aires, Argentina. Anthropologie 35: 197–205.
78. Zárate, M. & A. Blasi. 1993. Late Pleistocene–Holocene eolian deposits of the southern Buenos Aires province, Argentina: A preliminary model. Quaternary International 17: 15–20.
79. Zárate, M., M. Espinoza & L. Ferrero. 1995. La Horqueta II, río Quequén Grande: ambientes sedimentarios de la transición Pleistoceno–Holoceno. IV Jornadas Geológicas y Geofísicas Bonaerenses, Actas 1, Junín, pp. 195–204.
80. Zárate, M. & N. Flegenheimer. 1991. Geoarchaeology of the Cerro La China Locality (Buenos Aires, Argentina): Site 2 y Site 3. Geoarchaeology: an International Journal 6: 273–294.
81. Zárate, M.A., R.A. Kemp, M. Espinosa & L. Ferrero. 2000. Pedosedimentary and paleoenvironmental significance of a Holocene alluvial sequence in the southern Pampas, Argentina. The Holocene 10: 481–488.
82. Zucol, A.F., M.M. Colobig & D.L. Mazzanti. 2007. Análisis de microrrestos en muestras sedimentarias del Sitio 2 de la localidad arqueológica Amalia, provincia de Buenos Aires (Argentina). XVI Congreso Nacional Arqueología Argentina, Actas, Tomo I, pp. 141–147, S.S. de Jujuy.
83. Zucol, A.F., M. Brea, E. Passeggi & M.J. Franco. 2014. Colecciones del Laboratorio de Paleobotánica y Procesamiento de material sedimentario del CICYTTP-Diamante (CONICET), Entre Ríos, Argentina. Boletín de la Asociación Latinoamericana de Paleobotánica y Palinología 14: 71–82.
Recibido: 18-IX-2014
Aceptado: 18-XII-2014

