










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
versión On-line ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.16 no.2 Ciudad Autónoma de Buenos Aires dic. 2014
PALEONTOLOGÍA
Novedades palinológicas de las formaciones San José y Chiquimil (Mioceno medio y tardío), noroeste de Argentina
Lilia René Mautino¹ & Luisa Matilde Anzotegui
¹Centro de Ecología Aplicada del Litoral, Ruta 5 km 2,5, 3400 Corrientes, Argentina liliamautino@yahoo.com.ar
Resumen
En esta contribución se dan a conocer tres nuevos géneros fósiles: Cordiapollenites gen. nov., Euphortricolporites gen. nov. y Prosocolporites gen. nov., cuatro especies nuevas: Cordiapollenites espinulata sp. nov., Euphortricolporites perforata sp. nov., Prosocolporites argentina sp. nov., Cyperaceaepollis kyllingoides sp. nov., y una nueva combinación en Ailanthipites gamerroi Archangelsky. Las afinidades botánicas de estos taxones son a: Cordia spp., Boraginaceae (Cordiapollenites espinulata); Euphorbia spp., Euphorbiaceae (Euphortricolporites perforata); Prosopis spp., Fabaceae (Prosocolporites argentina) y Kyllinga spp., Cyperaceae (Cyperaceaepollis kyllingoides). Se realiza el análisis cronoestratigráfico de las familias de los taxones y se concluye que la mayoría de las afinidades botánicas de los géneros estudiados constituyen los primeros reportes palinológicos para el cenozoico argentino (lapso Paleoceno-Plioceno) a su vez ellos son incluidos en las comunidades previamente establecidas: hidrófila palustre dulceacuícola, higrófila, xerófila y de bosque de altura.
Palabras clave: Palinología; Mioceno; Taxones nuevos; Afinidades botánicas.
Abstract
New palynological taxa in the San Jose and Chiquimil formations (middle and late Miocene), northwest of Argentina. In this contribution three new fossil genera Cordiapollenites gen. nov., Euphortricolporites gen. nov., Prosocolporites gen. nov. and four new species Cordiapollenites espinulata sp. nov., Euphortricolporites perforata sp. nov., Prosocolporites argentina sp. nov. and Cyperaceaepollis kyllingoides sp. nov. and one new combination: Ailanthipites gamerroi Archangelsky are described. The botanical affinities of these taxa are with: Cordia spp., Boraginaceae (Cordiapollenites espinulata), Euphorbia spp. Euphorbiaceae (Euphortricolporites perforatae), Prosopis spp. Fabaceae (Prosocolporites argentina) and Kyllinga spp. Cyperaceae (Cyperaceaepollis kyllingoides). Chronostratigraphic analysis of the families of the taxa is realized. It is concluded that most of the botanical affinities of the genera under study constitute the first palynological reports of the Argentine Cenozoic (Paleocene-Pliocene interval). The proposal is that the paleocommunities in which the fossil taxa developed were: hydrophytic freshwater marsh, higrophyla vegetation, xerophytic plant community and upland forest.
Key words: Palynology; Miocene; New taxa; Botanical affinity.
INTRODUCCIÓN
En el análisis de la palinoflora de las formaciones San José (Mioceno medio) y Chiquimil (Mioceno tardío) Mautino (2010) detectó la predominancia de Angiospermae, sobre Gimnospermae (Spermatophyta), Monilophyta, Briophyta sensu lato, Zygnematales y "algas verdes". En éste trabajo se dan a conocer las últimas novedades taxonómicas en Angiospermae halladas en esas formaciones y que corresponden a tres nuevos géneros y una nueva combinación. La afinidad botánica de estos taxones a Cordia spp. Boraginaceae (Cordiapollenites espinulata), Euphorbia spp. Euphorbiaceae (Euphortricolporites perforata), Prosopis spp. Fabaceae (Prosocolporites argentina) y Kyllinga spp. Cyperaceae (Cyperaceaepollis kyllingoides) constituyen las primeras citas para la palinología del cenozoico argentino, en el lapso Paleoceno-Plioceno. Estas novedades taxonómicas se suman a otras previamente publicadas en Mautino & Anzótegui (1998, 2002a), Mautino et al. (2004) y Mautino (2007a, 2011).
Los taxones nuevos aquí descriptos son incluidos en las paleocomunidades ya establecidas en trabajos previos: hidrófila palustre dulceacuícola, bosque higrófilo, xerófila y bosque de altura (Mautino & Anzótegui, 1998, 2000, 2002a, b; Mautino et al., 2004 y Mautino, 2007a, 2010, 2011) y su presencia reafirma la existencia de las mismas. Al realizar el análisis de los registros cronoestratigráficos de sus respectivas familias, surge que la mayoría de los afines actuales, constituyen las primeras citas palinológicas en el Cenozoico de Argentina. Se señala, además, su distribución estratigráfica para diferenciar las formaciones de las que provienen.
Marco geológico
El grupo Santa María (Bossi & Palma, 1982) está conformado por las siguientes Formaciones: Saladillo, San José, Las Arcas, Chiquimil (Miembro A- Paraconglomerado Ampajango y Miembro B), Corral Quemado, Andalhuala y Yasyamayo; es un complejo sedimentario que corre paralelo al río Santa María sobre su margen derecha, adosándose por falla contra el basamento precámbrico de la sierra del Aconquija y Cumbres Calchaquíes. Las formaciones San José y Chiquimil se encuentran en la sección basal de dicho grupo. La primera (Galván & Ruiz Huidobro, 1965), aflora de N-S a lo largo del sector oriental del valle; su base está separada mediante una suave discordancia angular de la infrayacente Formación Saladillo y en transición de la suprayacente Formación Las Arcas. Su espesor, en general, varía entre 250- 315m y hacia el norte alcanza los 1000 m a la altura de la quebrada de Agua Negra en Salta. La litología más conspicua de esta Formación, consiste de una arenisca fina a mediana, de colores variados: pardos verdes, amarillos y grises, con predominancia del amarillo-verdoso, presenta estratificación de tipo lenticular (con plantas y palinomorfos, moluscos y microfósiles) tabular, cruzada y/o laminación ondulítica; las areniscas medianas están acompañadas por pelitas en general macizas o con laminación paralela, a veces en "paper shales". Posee un origen predominantemente lacustre, aunque hacia el norte se vuelve fluvial (Bossi et al., 1998). Estos autores adjudican a la formación una edad correspondiente al Mioceno medio (13 y 12 Ma) por la posición y relación estratigráfica de la misma con vulcanitas y tobas regionales de las que se tienen dataciones radimétricas.
La Formación Chiquimil, suprayace a la Formación Las Arcas, en el tope de la sección inferior del Grupo Santa María. Sus afloramientos se extienden desde el Río Ampajango al sur en Catamarca, hasta el Cerro Paranilla en Salta. Litológicamente se caracteriza por la alternancia de areniscas finas en conjuntos macizos, laminados, paralelos o con estratificación cruzada, que alternan con limolitas y arcilitas de colores variados entre pardo amarillento, rojo y verde claro en general es moderadamente salina y se hallan frecuentes venillas de yeso. En muchos niveles y principalmente en los superiores se intercalan conglomerados finos con rodados redondeados de vulcanitas (basaltos, andesitas, dacitas y metamórficas). La participación de rodados volcánicos es aquí mucho mayor que en las unidades subyacentes. Esta unidad ha sido dividida en dos miembros B (inferior) y A (superior) por Stahlecker (Marshall & Patterson, 1981) basados en vertebrados. El espesor de esta Formación varía entre los 480 metros en la localidad de Entre Ríos alcanzando un máximo de 1199 m en la quebrada de Agua Negra en Catamarca (Ibáñez, 2001). Las mediciones radimétricas señalan que su edad comprendida entre los 9,3-7,14 Ma corresponde al Mioceno tardío (Bossi et al., 1998; Kleinert & Strecker, 2001; Latorre et al., 1997). Los niveles con plantas, palinomorfos, moluscos y microfósiles, se encuentran generalmente en las porciones inferiores y medias de la Formación.
MATERIALES Y MÉTODOS
El material analizado proviene de los perfiles de 4 localidades muestreadas, Río Salinas y Km 107 (Tucumán) en la Formación San José; Río Vallecito y Quebrada de Jujuil (Catamarca) en la Formación Chiquimil (Ver figura 1). Las muestras fueron procesadas siguiendo las técnicas estándar en palinología consistentes de eliminación de silicatos con ácido fluorhídrico y oxidación con ácido nítrico; el montaje se realizó en glicerina-gelatina. Las preparaciones están depositadas en la colección CTES-PMP (Corrientes Preparados Micropaleontológicos) de la Universidad Nacional del Nordeste (UNNE). El análisis de las muestras se realizó con un microscopio óptico Leitz Diaplan y para las fotografías se utilizó una Cámara digital Canon Power Shot 550; las del microscopio electrónico de barrido, se tomaron con el equipo Jeol de la Universidad Nacional del Nordeste. Con el objetivo de ajustar la afinidad botánica se comparó el material fósil con actual a partir de preparados de referencia que pertenecen a la Palinoteca Corrientes (PAL-CTES), colección de la Universidad Nacional del Nordeste y se incorporaron nuevas preparaciones de botones florales de polen obtenidos del Herbario Corrientes (CTES).

Fig. 1. Mapa de ubicación del área de estudio (Modificado de Mautino, 2011).
En este trabajo, para establecer los registros previos de los taxones que resultaron novedades se consideró el lapso Cretácico tardío - Plioceno de Argentina.
SISTEMÁTICA PALINOLÓGICA
Clase TRICOLPORATAE Iversen & Troels Smith, 1950
Género. Cordiapollenites gen. nov.
Especie tipo. Cordiapollenites espinulata sp. nov.
Diagnosis. Pollen grains oblate spheroidal, isopolar, radially symmetrical, subcircular to subtriangular amb. Tricolporatae. Colpi short. Ora with annulus. Exine tectate with spinula and grana.
Diagnosis. Granos de polen oblatos esferoidales, isopolares, radiosimétricos, ámbito subcircular a subtriangular. Tricolporados. Colpos cortos. Ora con anillo. Exina tectada, con espínulas y gránulos.
Discusión. El polen del género Cordia L. fue reconocido en el Mioceno de las Islas Marshall en el oeste del Pacífico por Leopold (1969) y fue descripto como cf. Cordia subcordata Lam. se diferencia de la especie nueva aquí descripta porque son granos tricolpados con escultura clavada.
Los géneros fósiles con mayores similitudes son Compositoipollenites Potonié 1960 y Spinulaepollenites Krutzsch 1962. El primero incluye formas triporadas con dudosos colpos, exina equinada y reticulo negativo. Spinulaepollenites tiene ámbito hexagonal, posiblemente es tricolporado con espinas pequeñas, tectado y está relacionado a la especie Arceuthobium oxycedri Bieb. de la familia Loranthaceae.
Como consecuencia, el género Cordiapollenites es creado para reunir a granos de polen notoriamente tricolporados, con exina tectada, espínulas y gránulos; con afinidad botánica al género Cordia L. (Boraginaceae).
Derivatio nominis. El nombre alude a la afinidad botánica con el género actual Cordia L. de la familia Boraginaceae.
Afinidad botánica. Cordia es un género euripalínico y comprende granos triporados,
tricolpados, tricolporados a tetracolporados, con exina tectada con espínulas, verrugas, gránulos o semitectada reticulada a estriado-reticulada. El género fósil Cordiapollenites gen. nov. está vinculado a las especies tricolporadas con exina tectada, espínulas y escasos gránulos.
Cordiapollenites espinulata sp. nov.
(Figs. 2A-D)
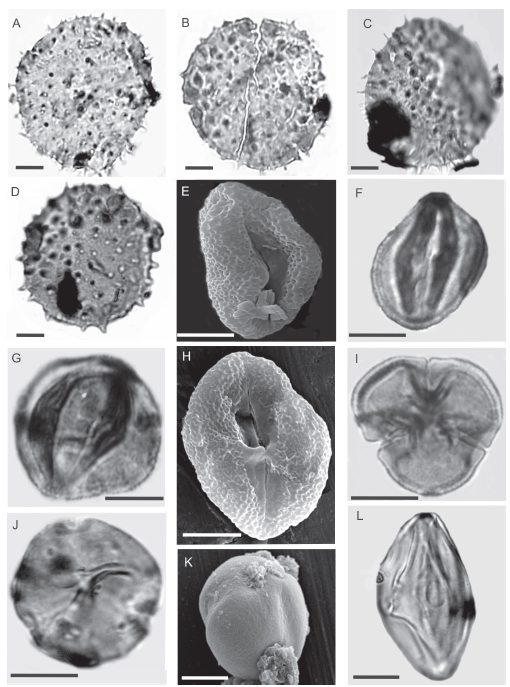
Fig. 2. Cordiapollenites espinulata sp. nov. A-D. A, Holotipo. CTES-PMP 1948b: 42,9/102. Foco en las aperturas. B, Paratipo. CTES-PMP 1841c: 43,8/108,5. C, CTES-PMP 1948b 45,5/104,5. D, CTES-PMP 1841c 39,5/104,7. Euphortricolporites perforata sp. nov. E-I. E-H. MEB 2268. F-G, Vista sub-ecuatorial CTES-PMP 1795a 45,3/92 y 1795b 42/91. I, Holotipo. CTES-PMP 1795g: 34,3/95,2. Prosocolporites argentina J-L. J, Paratipo CTES-PMP 2268e: 46,4/102,6. K, MEB CTES.PMP 2268 L, Holotipo. CTES-PMP 1942c: 41,4/ 98,7. Escalas gráficas = 10μm.
Compositoipollenites sp. Mautino 2007b. Reunión Anual de Comunicaciones de la Asociación Paleontológica Argentina: 15.
Diagnosis. Granos de polen oblatos esferoidales, isopolares, de ámbito subcircular a subtriangular. Tricolporados. Ora con anillos. Exina tectada, con estratificación nítida. Téctum con espínulas y escasos gránulos. Espínulas de ápice agudo.
Diagnosis. Pollen grains oblate spheroidal, isopolar, subcircular to sub-triangular amb. Tricolporate. Ora with thickenings. Tectate with distinct stratification. Tectum with spinules and little granules. Spinules of sharp pointed.
Descripción. Granos de polen oblato esferoidal de 42 (46) 50 μm de diámetro ecuatorial y 40 (44) 48 μm de diámetro polar, isopolares, de ámbito subcircular a subtriangular. Tricolporados. Ora de 2 μm de diámetro con anillo de 2-2,5 μm. Exina tectada, de 2-2,5 μm de espesor (sexina 1,5-2 y nexina de 0,5 μm) estratificación con báculas notorias. Téctum con espínulas y escasos gránulos. Apocolpios de 25-28 μm. Espínulas de ápice agudo de 1,5-2 μm de alto, base de 0,7-1 μm de espesor, distantes entre sí por 3,5-4 μm.
Holotipo. CTES-PMP 1948b: 42,9/102.
Paratipo. CTES-PMP 1841c: 43,8/108,5.
Localidad tipo. Km 107, provincia de Tucumán.
Edad. Mioceno medio.
Material adicional. Ver Anexo.
Derivatio nominis. El nombre de la especie hace alusión a las espínulas supratectales.
Comparaciones. Cordia cf. subcordata Leopold (1969), del Mioceno tardío de las Islas Marshall (región de Micronesia en el océano Pacífico), se diferencia de la especie aquí descripta, porque tiene granos tricolpados con sexina irregularmente clavada, en la que las cabezas de las clavas se unen en algunos sectores produciendo un modelo vermiculado. Estos granos se relacionan a la especie actual Cordia subcordata Lam.
Numerosos son los estudios palinológicos actuales del género Cordia (Nowicke & Ridgway (1973), Nowicke & Miller (1990), Scheel & Barth (1995) y Scheel et al. (1996); que permiten asignar a Cordiapollenites espinulata sp. nov. al Tipo Polínico 1 "Tricolporado espinoso" establecido por Scheel & Barth (1995) y Scheel et al. (1996) quienes a su vez lo dividen en Subtipo 1 y 2. Si bien existe bastante similitud con todas las especies de ambos subtipos, se observan mayores semejanzas con Cordia sellowiana Cham., Cordia ecalyculata Vell. (ambas del subtipo 1) y Cordia trichotoma (Vell.) Arráb.ex Steud. Por lo expuesto, a pesar de que el género Cordia L. es euripolínico la especie nueva se vincula con las formas tricolporadas equinadas escasamente granuladas.
Los caracteres palinológicos definidos y la clara afinidad botánica con el género Cordia ameritan la creación de este nuevo taxón.
Afinidad botánica. Boraginaceae, Cordia sellowiana Cham., Cordia ecalyculata Vell. y Cordia trichotoma (Vell.) Arráb. ex Steud.
Distribución geográfica y ambiente de los taxones actuales afines. La familia Boraginaceae comprende hierbas y en menor proporción árboles y arbustos. Alberga 100 géneros y 2000 especies de distribución cosmopolita. Cordia es uno de los géneros más numerosos y cuenta con cerca de 300 especies, principalmente tropicales en ambos hemisferios (Cronquist, 1981).
Cordia trichotoma, Cordia sellowiana y Cordia ecalyculata tienen porte arbóreo. La primera de ellas ("peteribí o loro negro") crece en las Provincias de Corrientes, Jujuy, Misiones, Salta y Tucumán. Integra la selva de transición de la Provincia de las Yungas, y la comunidad clímax de la Provincia Paranaense (Cabrera, 1976). Esta especie también se encuentra en Brasil, donde se destaca por su predominancia en el estrato emergente de la Floresta subtropical y en el sub-bosque de floresta de Araucaria (Scheel & Barth, 1995). Por otro lado, Cordia sellowiana es nativa de Brasil y no se encuentra en Argentina. Finalmente, Cordia ecalyculata habita en selvas en galería de Argentina (Misiones y Corrientes) y Brasil. Además fue hallada en la provincia de Corrientes, en bosques higrófilos del Iberá (Araujo et al., 2004; Tressens et al., 2002).
Género. Euphortricolporites gen. nov.
Especie tipo. Euphortricolporites perforata sp. nov.
Diagnosis. Granos de polen subprolatos a prolatos, isopolares, radiosimétricos, ámbito subtriangular a lobado de lados rectos o ligeramente convexos y ángulos redondeados. Tricolporados. Colpos largos que llegan al área polar, rodeados por un margen. Ora lalongados. Exina tectada-perforada, con sexina y endexina bien diferenciadas.
Diagnosis. Pollen grains subprolate to prolate, isopolar, radially symmetrical, amb subtriangular to lobate, straight sides and rounded point. Tricolporatae. Colpi long reaching the area polar and surrounded by a margin. Ora lalongate. Exine tectate-perforate with sexine and endexine very distinct tectate perforate.
Discusión. Se crea el género Euphortricolporites para reunir granos de polen tricolporados, con margen rodeando los colpos, exina tectada perforada y afinidad a Euphorbia L. (Euphorbiaceae). Si bien el género fósil Euphorbiacites Sung & Lee también reúne granos de polen tricolporados con afinidad a Euphorbia L., se diferencia del creado en esta contribución, dado que tiene exina reticulada (semitectada) y no tectada perforada (Sung & Lee, 1976).
Derivatio nominis. El nombre genérico se refiere a la afinidad botánica con el género Euphorbia L. de la familia Euphorbiaceae y al tipo de aperturas que caracterizan a estos granos de polen.
Afinidad botánica. De la comparación de los granos fósiles con los actuales se encontró mayor afinidad al tipo polínico Euphorbia villosa Willd., particularmente por la presencia de un margen en los colpos y por la sexina perforada. Este tipo polínico fue instituido por El-Ghazaly & Chaudhary (1993) para el género Euphorbia, quienes a partir de 60 especies examinadas exhaustivamente establecieron 7 tipos polínicos.
Euphortricolporites perforata sp. nov.
(Figs. 2 E-I)
Rhoipites sp. Mautino 2007b. Reunión de Comunicaciones de la Asociación Paleontológica Argentina. 50° Aniversario de Ameghiniana: 15.
Diagnosis. Granos de polen subprolatos a prolato esferoidal, isopolares, radiosimétricos, ámbito subtriangular a lobado de lados rectos y ángulos redondeados. Tricolporados. Colpos acompañados por un margen psilado y amplio. Ora lalongados. Exina tectada perforada.
Diagnosis. Pollen grains subprolate to prolate spheroidal, isopolar, radially symmetrical, sub-triangular to lobate amb with straight sides and rounded point. Tricolporate. Colpi accompanied by a smooth and ample margin. Ora lalongate. Exine tectate, perforated.
Descripción. Granos de polen de 22 (26,5) 31 μm de diámetro polar y 14 (19,5) 25 μm de diámetro ecuatorial, isopolares, radiosimétricos, ámbito subtriangular de lados rectos y ángulos redondeados, de forma subprolata a prolato esferoidal. Tricolporados, colpos sobrepasan el área polar y están limitados por un margen psilado y amplio de 4 a 7,5 μm de ancho, bien visible en vista polar. Ora lalongados de 2-3,5 μm x 4-9 μm. Exina tectada, de 2 a 3 μm de espesor, notoriamente estratificada (sexina de 1,5-2 μm y nexina de 0,5-1 μm) y perforada.
El MEB confirma la presencia de un margen (sexínico) psilado acompañando a los colpos y también a los ora (vista ecuatorial). Además revela que la membrana del colpo es granulada y que la exina es perforada, con las perforaciones simples homogéneamente distribuídas, rara vez dispuestas en grupos de 2 o 3 perforaciones más próximas.
Holotipo. CTES-PMP 1795g: 34,3/95,2.
Paratipo. CTES-PMP 1795b 40,8/100.
Localidad tipo. Río Vallecito, provincia de Catamarca.
Edad. Mioceno tardío.
Material adicional. Ver Anexo.
Derivatio nominis. El nombre hace alusión a la estructura perforada de la sexina.
Comentarios. Algunos palinomorfos que integran esta especie fueron asignados equívoca
mente a Rhoipites alveolatus (Couper) Pocknall y Crosbie emend. Mautino y Anzótegui (Mautino & Anzótegui, 2002b lámina 1 A, S y T), porque posee margen psilado y exina de 2-3 μm de espesor, pero se diferencia de la especie nueva, aquí descripta, porque los granos son de mayor tamaño (30-53 μm x 29-44 μm), con exina microrreticulada (diámetro polar: 30-53) y diámetro ecuatorial de forma oblata a subesferoidal; en cambio Euphorbiacites perforata tiene forma subprolata a prolata esferoidal y la exina es tectada perforada.
Comparaciones. En la literatura el género fósil más relacionado es Euphorbiacites Sung y Lee 1976 cuya especie tipo E. wallensenensis es de mayor tamaño (diámetro polar: 35-50 μm), tiene ora circulares y sexina reticulada. Otras especies del género con las que se comparó son Euphorbiacites reticulatus Sung y Lee, Euphorbiacites sp. 1 y Euphorbiacites sp. 2, las que difieren de la especie nueva aquí descripta, por poseer la sexina reticulada, aunque comparten el margen colpal psilado que las relaciona además a especies actuales de Euphorbiaceae. Algunas especies del género Rhoipites poseen exina tectada perforada, entre otras, Rhoipites exiguus Pocknall 1982 y Rhoipites rimulatus Pocknall 1982; pero la primera se diferencia de esta nueva especie porque es de forma esferoidal y no subprolata a prolata esferoidal, por las menores dimensiones en el tamaño del grano, de los ora y del espesor de la exina. Rhoipites rimulatus Pocknall 1982, se distingue por la presencia de rímula y de un mayor tamaño del diámetro polar (35-47 μm). Ambas especies además se diferencian de la especie nueva porque la exina se encuentra ligeramente engrosada a nivel del ecuador. "Rhoipites" cienaguensis (Dueñas) Barreda 1997 (Barreda, 1997a) a diferencia de esta nueva especie reúne granos oblatos, con exina semitectada, reticulada, homobrocada, de menor espesor (1 μm); y si bien al igual que estos granos son tricolporados, los márgenes colpales están débilmente engrosados.
Otras especies fósiles similares son mencionadas sin descripción, como cf. Bernardia P. Browne (figs. 122-123; Euphorbiaceae) por Graham (1976) en el Mioceno de México; Tipo Euphorbiaceae (lam. 21, fig.4) en el Cuaternario de Brasil (García, 1994) y Euphorbia plagiantha Drake (fig. 1 y 2) en Muller et al. (1989).
Al comparar la especie nueva con los 7 tipos polínicos establecidos por El-Ghazaly & Chaudhary (1993), a partir de especies actuales, surge un mayor grado de similitud con el tipo polínico 1a- Euphorbia villosa, en especial el subtipo 1a- E. villosa (láminas I y II) que reúne 15 especies con morfologías muy parecidas; particularmente comparten la sexina perforada, margen engrosado en las aperturas y membrana del colpo granulada (visto al MEB).
Por las características antes mencionadas se justifica la creación de esta nueva especie.
Afinidad botánica. Euphorbiaceae, Euphorbia L. (Tipo polínico Euphorbia villosa).
Distribución geográfica y ambiente de los taxones actuales afines. La familia Euphorbiaceae consta de 317 a 339 géneros y cerca de 8100 especies; tiene una distribución cosmopolita, predominando en regiones tropicales y subtropicales, con excepción de las zonas polares. El género Euphorbia es el más diverso de la familia e incluye aproximadamente 1.000 especies en todo el mundo (Martínez Gordillo & Morrone, 2005). En Argentina cuenta con 53 especies nativas (Dawson, 1965; Zuloaga & Morrone, 1999). Son hierbas y arbustos de regiones secas, que crecen en sabanas y praderas; varias especies de Euphorbia viven junto a Tribulus terrestris, Gomphrena spp., Prosopis spp. y Boungainvillea spp., conformando la comunidad climax de el "Jarillal" (Larrea nitida) en la Provincia del Monte (Cabrera, 1976).
El-Ghazaly & Chaudhay op cit. consideran que el tipo polínico 1- Euphorbia villosa esta ubicado en la base de la línea evolutiva propuesta para el género Euphorbia L. sobre un total de 7 tipos establecidos.
Por otra parte, Martínez Gordillo & Morrone (2005) en el análisis biogeográfico de la familia Euphorbiaceae, establecen que la subfamilia Euphorbioideae (que contiene el género Euphorbia) tiene mayor número de endemismos en la región Neotropical y Afrotropical, seguidos de la región Oriental.
Género. Prosocolporites gen. nov.
Diagnosis. Granos de polen prolatos, isopolares, radiosimétricos. Tricolporados, colpos con membrana apertural, ora con anillos. Exina tectada, escabrada.
Diagnosis. Pollen grains prolate, isopolar, radially symmetrical. Tricolporate, colpi with membrane, ora with annulus. Exine tectate scabrate.
Discusión. Este género es creado para introducir granos de polen de forma prolata, con exina escabrada, tricolporados con membrana colpal y con afinidad al género actual Prosopis L. Los géneros fósiles relacionados son Margocolporites (Ramanujan ex Srivastava) Pocknall y Mildenhall 1984, Alangiopollis Krutzsch 1962 y Dryadopollis Srivastava 1977. Estos géneros fósiles comparten con el nuevo taxón, las aperturas tricolporadas con membrana colpal aunque difieren porque la sexina es reticulada. Además Margocolporites incluye granos oblatos a suboblatos, colpos con margen liso, escultura granular o microreticulada y en Alangiopollis el tamaño de los granos y el espesor de la exina son superiores (92 μm y 5 μm respectivamente) además de la forma esferoidal y la sexina reticulado-estriada. Finalmente Dryadopollis Srivastava 1977, presenta exina con lúmenes más grandes en los mesocolpios que en los apocolpios y es de forma oblata a prolata.
Derivatio nominis. El nombre del género fósil deriva de la afinidad botánica con el género actual Prosopis L. (Fabaceae).
Afinidad botánica. Prosopis L. Fabaceae.
Prosocolporites argentina sp. nov.
(Figs. 2 J-L)
Margocolporites sp. Mautino, 2007c. Boletín de la Sociedad Argentina de Botánica (42): 120.
Margocolporites sp. Mautino, 2007b. Reunión Anual de Comunicaciones de la Asociación Paleontológica Argentina: 15.
Diagnosis. Granos de polen prolatos, isopolares, ámbito subcircular a subtriangular. Tricolporados, colpos con membrana apertural, ora con anillo. Exina tectada, escabrada.
Diagnosis. Grains of pollen prolate isopolar, subcircular to subtriangular amb. Tricolporatae, colpus membrane and oras with annulus. Exine tectate scabrate.
Descripción. Granos de polen prolatos de 17 (21) 25 μm de diámetro ecuatorial y 30 (32,4) 34,8 μm de diámetro polar, radiosimétricos, isopolares, de ámbito subcircular a ligeramente subtriangular. Tricolporados. Colpos, en vista polar, con límites poco definidos, miden en el ecuador 8 a 10 μm de ancho máximo, con membrana apertural psilada. Ora lalongados (2-4 μm) a subcirculares (4-4 μm, en el sentido ecuatorial/polar) con anillos de 1-1,5 μm. Apocolpio 3,5-4,2 μm de ancho. Exina tectada, de 1,5-2 μm de espesor, (sexina 1 a 1,5 y nexina de 0,5 μm). Téctum escabrado. El MEB revela que las membranas colpales son escabradas a psiladas y que la sexina es fósulo-perforada (Fig.2 K).
Observaciones. Los anillos de los ora son observados únicamente cuando los granos se encuentran en vista sub-ecuatorial y polar tanto en fósiles como actuales.
Holotipo. CTES-PMP 1942c: 41,4/ 98,7.
Paratipo. CTES-PMP 2268e: 46,4/102,6.
Localidad tipo. Km 107, provincia de Tucumán.
Edad. Mioceno medio.
Material adicional. Ver Anexo.
Derivatio nominis. El nombre de la especie se refiere al país (Argentina) en el que han sido hallados los palinomorfos.
Comparaciones. Los primeros registros palinológicos fósiles mundiales del género Prosopis son los de Prosopis quesneli del Oligoceno de Columbia Británica, Canadá (Piel, 1971) y Prosopis farcta encontrado en el Mioceno de Francia (Guinet & Bessedik, 1984). El primero de estos presenta exina reticulada y está relacionado a Prosopis juliflora (Swartz) De Candolle; en cambio P. farcta se diferencia porque la forma del grano es oblata y la sexina perforada con areolas redondeadas o sinuosas distribuidas irregularmente. Por otra parte, Caccavari (1996), relaciona a Retitricolporites medius González Guzmán, 1967 (Eoceno de Colombia) con las especies actuales Prosopis caldenia y P. campestris de la sección Algarobia. Esta especie colombiana difiere del taxón aquí descripto dado que posee exina semitectada reticulada.
En sedimentos de Argentina, Prámparo et al., (1996) describen Margocolporites sp., en el Eoceno-Mioceno de San Juan, y lo relacionan a Jacaranda sp. (Bignoniaceae). No obstante también podría compararse con la nueva especie, aquí descripta, por la presencia de la membrana colpal, pero se diferencia por el mayor tamaño de los ora (6-7 μm) y la forma oblata de los granos.
Prosocolporites argentina sp. nov. por el tamaño pequeño a mediano, forma prolata, presencia de membrana colpal, ora con anillos, sexina tectada-escabrada (con MO) es similar a especies actuales del género Prosopis L. (Fabaceae) y particularmente al Tipo polínico Prosopis alba (Fernández Pacella et al., 2013). Este tipo polínico incluye especies tanto de la sección Algarobia como de Strombocarpa de acuerdo a las secciones establecidas por Burkart (1976). La nueva especie se convierte en el registro más antiguo de polen de Prosopis para Argentina.
Afinidad botánica. Fabaceae, Mimosoideae, Tipo polínico Prosopis alba.
Distribución geográfica y ambiente de los taxones actuales afines. Prosopis comúnmente integra regiones semiáridas y áridas de América del Sur, del Norte y Central, oeste de Asia y África; unas pocas especies viven en zonas tropicales sub-húmedas o subtropicales. Este gé
nero cuenta con 45 especies, pero en la Argentina solo con 28 especies (10 especies y 6 variedades son endémicas), donde tiene la máxima concentración desde Jujuy hasta al norte patagónico (Burkart, 1987; Catalano et al., 2008, Kurkbride et al., 2003). Comprende árboles y arbustos dominantes en la Provincia del Monte formando el jarillal en bolsones y llanuras de suelo arenoso o pedregoso-arenoso. También es uno de los principales componentes de los bosques xerófilos o abiertos del Iberá (Tressens et al., 2002) en la provincia de Corrientes.
Género. Ailanthipites Wodehouse 1933
Especie tipo. Ailanthipites berryi Wodehouse 1933
Observaciones al género. Milne (1988) sinonimiza Striatricolporites Hammen 1956 al género Ailanthipites Wodehouse 1933. Ambos morfogéneros reúnen granos con aperturas tricolporadas, ora lalongados y exina tectada a semitectada con sexina estriada a supraestriada-infrarreticulada. Por lo tanto, por el principio de prioridad debe usarse Ailanthipites. El carácter supraestriado-infrarreticulado de la sexina, lo relaciona con varias familias actuales de angiospermas, tales como Anacardiaceae, Fabaceae, Sapindaceae, Burseraceae, Rosaceae (Milne, 1988) y Simaroubaceae (Wodehouse, 1933) lo que entorpece la tarea de lograr la asignación botánica para estas formas.
Ailanthipites gamerroi (Archangelsky) nov. comb.
(Figs. 3 A-F)

Fig. 3. Ailanthipites gamerroi Archangelsky nov. comb. A-F. Grupo 1. A, MEB CTES-PMP 1695. B, 1695a: 47/106,5 C, vista polar CTES-PMP 1695m: 39,5/98,3. Grupo 2. D, MEB CTES-PMP 2268 E, CTES-PMP 1946d 39,2/107,5.F, CTES-PMP 2268 39.4/56,7. Cyperaceaepollis kyllingoides. G-I. G-H, MEB CTES-PMP 2265.I, Holotipo. CTES-PMP 1945c: 38,7/100,2. Escalas gráficas= 10 μm.
Basónimo.
1973. Striatricolporites gamerroi Archangelsky: 381-382, lám. 9, fig. 1,2,3.
Sinónimos.
1986. Striatricolporites gamerroi Archangelsky en Romero y Castro: 110-111, lám. 1, figs.1-3.
1986. Schinus sp. Anzótegui & Garralla: 130, lám. 6, fig. 70.
1986. Astronium sp. Anzótegui & Garralla: 130-131, lám. 5, fig. 69.
1987. Striatricolporites gamerroi Archangelsky en Baldoni: 389-390, Lám. VII, figs. 4,5,6.
1989. Striatricolporites gamerroi Archangelsky en Barreda: 125-127, lám. 26, figs. 10 a 12; lám. 27, figs. 6 a 9.
1996. Striatricolporites gamerroi Archangelsky en Ruiz & Quattrocchio: 5, lám. 2, K.
1997a. Striatricolporites gamerroi Archangelsky en Barreda: 136, lám. 3, figs. 9-12.
1997b. Striatricolporites gamerroi Archangelsky en Barreda: 285, lám. II, fig. 6.
1997. Striatricolporites gamerroi Archangelsky en Ruiz & Quattrocchio: 128, lám. C, Figs. 10 y 11.
1998. Striatricolporites gamerroi Archangelsky en Ottone et al.: 44, lám. 2, fig. 6.
1998. Striatricolporites gamerroi Archangelsky en Barreda et al.: 327, lám. 2, fig. 3.
2000b. Striatricolporites gamerroi Archangelsky en Barreda & Palamarczuck: 116, fig. 5.N.
2000. Striatricolporites gamerroi Archangelsky en Palamarzuck & Barreda: 234.
2001. Striatricolporites gamerroi Archangelsky en Guler et al., lám. 3, fig. 1.
2004. Striatricolporites gamerroi Archangelsky en Palazzesi & Barreda: 359, fig. 3.D.
2009. Striatricolporites gamerroi Archangelsky en Nañez et al.: 677, fig. 10, 3.
Descripción. Granos de polen prolatos de 23 (26,5) 30 μm de diámetro polar y 15 (18,5) 22 μm de diámetro ecuatorial, isopolares, radiosimétricos. Tricolporados, ora lalongados de 1-2,5 μm x 4-9 μm y colpos extendiéndose hasta las proximidades de los polos, con engrosamientos que en el ecuador miden de 3-4 μm. Exina de 1-1,5 μm de espesor; semitectada, supraestriada-infrarreticulada, estrías 0,5-0,7 μm de ancho y 2-4 μm de largo; dispuestas por lo general en el sentido del eje polar, muros de 0,5 μm de altura; microretículo con lúmenes de 0,5 μm de diámetro a 1 μm.
Principal material estudiado. Ver anexo.
Comentarios. Archangelsky (1973) establece que Striaticolporites gamerroi tiene afinidad botánica con las familias actuales Solanaceae y Rosaceae. Romero & Castro (1986), le confieren afinidad con las Anacardiaceae, Gentianaceae, algunas Rosaceae, Rutaceae y Nolanaceae; sin embargo, sugieren que la familia más afín es Anacardiaceae. Desde ese momento esta afinidad fue utilizada por varios palinólogos (entre otros Anzótegui & Garralla, 1986; Barreda, 1989; Ottone et al., 1998; Guler et al., 2001) y es el criterio seguido en este trabajo.
A pesar de que la familia Anacardiaceae es estenopalínica, Anzótegui (2001), reconoció varios tipos y subtipos polínicos; como consecuencia, en este trabajo es posible reconocer dos Grupos (1 y 2). Estos, no adquieren un rango taxonómico de especie porque para ello deberían revisarse todos los registros de Striatricolporites gamerroi que se recombinaron a Ailanthipites.
Striatricolporites gamerroi Grupo 1 corresponde al Subtipo Schinus weinmanniifolia var. weinmanniifolia Engl., tiene granos con estrías angostas y cortas (hasta 2,5 μm de largo), lúmenes del retículo de 0,5 μm y en vista polar se puede apreciar que la sexina tiende a ser reticulada. Striatricolporites gamerroi Grupo 2, se relaciona al subtipo Lithraea molleoides (Vell.) Engl., y presenta palinomorfos con estrías angostas y largas (2,5 a 4 μm), los lúmenes del retículo pueden llegar a 1 μm de diámetro, y las endoaperturas presentan un pliegue sobre el área ecuatorial.
Striatricolporites gamerroi Grupo 1, proviene de las localidades Río Vallecito y Quebrada de Jujuil (Formación Chiquimil); Striatricolporites gamerroi Grupo 2 de las localidades Km 107 y Río Salinas (Formación San José)
Striatricolporites gamerroi Grupo 1, es semejante a los hallados por Anzótegui (2004 y 2006) en materia orgánica amorfa (presumiblemente producto de la carbonización de inflorescencias) en los mismos niveles y entremezclados con impresiones de hojas de Schinus aff. terebinthifolia Raddi en la localidad Río Vallecito. Fleig (1987) incluye en sinonimia de Schinus terebinthifolia a Schinus weinmanniifolia Engl. y sus variedades, porque considera que la gran variabilidad que poseen estas especies no las habilita para diferenciarlas, criterio seguido por Anzótegui (2004).
Por otra parte y coincidentemente con los palinomorfos incluidos en Striatricolporites gamerroi Grupo 2, Anzótegui (2006) en la localidad Km 107 (Fm. San José) encontró foliolos de Lithraea molleoides ("molle de beber").
Schinus sp. y Astronium sp., palinomorfos hallados en el Mioceno de Formación Paraná por Anzótegui & Garralla (1986) son semejantes Striatricolporites gamerroi Grupo 1 y Lithrea aff. brasiliensis March. al Grupo 2.
Como antecedente, se debe mencionar que Archagenslky (1973) observó que A. gamerroi (Striatricolporites gamerroi) podría ser subdividido en dos o tres especies distintas, por lo que, en el futuro, ese material debería ser re-examinado. A. gamerroi de Formación Chenque (Barreda, 1989) a juzgar por las ilustraciones de MEB es muy parecido a los del Grupo 2 aquí establecido, especialmente por la longitud de las estrías.
En síntesis esta especie (A. gamerroi) reúne dos formas que fueron definidas usando el MEB. Se hace imperiosa la necesidad de una revisión mayor, que incluya imágenes al MEB de los registros previos mencionados en sinonimia (sinónimos y basónimos). Luego de una exhaustiva observación y descripción se podrían crear dos taxones con definida afinidad botánica.
Registros previos. Argentina: Cretácico a Paleoceno (Ruiz & Quattrocchio, 1996; Quattrocchio & Ruiz, 1999); Paleoceno a Mioceno (Archangelsky, 1973; Anzótegui & Garralla, 1986; Barreda, 1989, 1996, 1997a y 1997b; Barreda et al., 1998; Ottone et al., 1998; Romero & Castro, 1986; Barreda & Palamarczuck, 2000b; Palamarczuck & Barreda, 2000; Palazzesi & Barreda, 2004); Mioceno-Plioceno (Guler et al., 2001).
Afinidad botánica. Anacardiaceae, Striatricolporites gamerroi Grupo 1 a Schinus terebinthifolia y Striatricolporites gamerroi Grupo 2 a Lithrea molleoides.
Distribución geográfica y ambiente de los taxones actuales afines. La familia Anacardiaceae esta constituida por árboles, arbustos y lianas, raramente hierbas, consta de 60 a 80 géneros con aproximadamente 850 especies con una distribución principalmente pantropical, tropical y subtropical, pocas especies crecen en regiones templadas (Dong & Baas, 1993).
Schinus terebinthifolia y sus variedades son arbustos y árboles del noreste de Argentina que integran las Provincias fitogeográficas Paranaense, Chaqueña y del Espinal, se desarrollan en la costa de ríos con suelos anegadizos en formaciones boscosas (Cabrera, 1976; Muñoz, 1990), también son hallados en Paraguay, Uruguay y sur de Brasil.
Lithrea molleoides es un árbol o arbusto del centro, y norte de Argentina, sur de Brasil, Bolivia, Paraguay y Uruguay. Es una especie dominante del distrito Chaqueño Serrano de la Provincia fitogeográfica Chaqueña, donde constituye la comunidad clímax en los bosques de "Horco Quebracho" adquiriendo hábito orófilo. En el Iberá (Corrientes) integra los bosques xerófilos con Prosopis. En la provincia fitogeográfica Paranaense es una especie secundaria de las selvas marginales (Cabrera, 1976; Muñoz, 2000, Tressens et al., 2002).
Clase PERIPORATAE Iversen & Troels Smith, 1950
Género. Cyperaceaepollis Krutzsch 1970
Especie tipo. Cyperaceaepollis neogenicus Krutzsch 1970.
Cyperaceaepollis kyllingoides sp. nov.
(Figs. 3 G-I)
Diagnosis. Granos de polen subesferoidales a poliédricos, apolares. Periporados con 6 poroides de límites imprecisos. Exina tectada, escabrada.
Diagnosis. Pollen grain sub-spheroidal to polyhedral apolar. Periporate with 6 poroid, ill-de-fined. Exine tectate, scabrate.
Descripción. Granos de polen de 14 (18,5) 23 μm de diámetro, apolares, subesferoidales a poliédricos debido a la posición de las aperturas. Periporados con 6 poroides, de 4-6 μm de alto y 5 a 10 μm de largo, de límites algo imprecisos, de tamaño variable en un mismo ejemplar, en algunos poroides se observa la membrana apertural. Exina delgada de 0,5 μm de espesor, con estratificación muy sutil tectada, escabrada.
El MEB revela que el téctum posee microgranulaciones.
Holotipo. CTES-PMP 1945c: 38,7/100,2.
Paratipo. CTES-PMP 1795f: 41,5/95.
Localidad tipo. Km 107, Quebrada de Amaicha, noroeste de la provincia de Tucumán.
Edad. Mioceno Medio.
Material adicional. Ver Anexo.
Derivatio nominis. El nombre está referido a la afinidad al género Kyllinga Rotlb.
Comparaciones. Cyperaceaepollis kyllingoides, se diferencia de C. neogenicus Krutzsch 1970 (Oligoceno de la Formación Ganchaigou de Alemania) porque es un grano apolar, con 6 poroides en cambio C. neogenicus es heteropolar, con la típica forma de pera, ulcus distal (poro único de ubicación polar distal) y 4 poros subecuatoriales.
Esta forma es parecida al Tipo Cyperaceae 1 (Lámina 29; Fig. 4 a/b/c) ilustrado por García (1994) de las turberas cuaternarias de Brasil, y también a la especie actual Kyllinga erecta Schum. y Tonn. (Ybert, 1979), por el tamaño y aspecto general; aunque K. erecta posee solamente cinco poroides.
De las 4 especies de Kyllinga Rottb. que se encuentran en Argentina, con la que tiene mayores similitudes es con K. pumila Michx. (PAL-CTES 7288) particularmente porque son granos apolares con la misma cantidad de poroides, aunque en esta especie la estratificación de la exina es más marcada que en la del fósil.
Afinidad botánica. Cyperaceae, Kyllinga spp., es la primera mención del género para el Mioceno argentino.
Distribución geográfica y ambiente de las especies actuales afines. La familia Cyperaceae tiene una distribución cosmopolita y cuenta en la actualidad con cerca de 240 especies herbáceas, relacionadas generalmente a ambientes húmedos y palustres. Las especies de Kyllinga son hierbas que crecen en distintos ambientes acuáticos y zonas bajas (Barros, 1947). K. pumila es una especie nativa y palustre de Argentina y se halla en las provincias de Tucumán, Salta, Misiones, Formosa, Entre Ríos y Corrientes (Biganzoli & Múlgura de Romero, 2004; Zuloaga & Morrone, 1999).
DISCUSIÓN Y CONCLUSIONES
En éste trabajo, en el que se dan a conocer las últimas novedades taxonómicas angiospérmicas halladas en las Formaciones San José y Chiquimil (tres nuevos géneros: Cordiapollenites gen. nov., Euphortricolporites gen. nov. y Prosocolporites gen. nov., cuatro especies: Cordiapollenites espinulata sp. nov., Euphortricolporites perforata sp. nov., Prosocolporites argentina sp. nov. y Cyperaceaepollis kyllingoides sp. nov. y una nueva combinación Ailanthipites gamerroi Archangelsky 1973, se realiza una revisión cronoestratigráfica de las familias que contienen las especies nuevas en el lapso Cretácico tardío-Plioceno de Argentina; se puntualiza la importancia estratigráfica de estos taxa para diferenciar las formaciones de las que provienen (Formaciones San José y Chiquimil) y se infieren las paleocomunidades que habrían integrado.
De la mencionada revisión se rescató lo siguiente:
Boraginaceae. Los únicos registros fósiles de esta familia derivan de los leños descriptos por Brea & Zucol (2006) Cordioxylon prototrichotoma afines a Cordia trichotoma y en menor grado a Cordia glabrata (Mart.) A. DC., en la Formación Peñas Coloradas, Paleoceno tardío de la provincia de Chubut. Constituyéndose Cordiapollenites espinulata sp. nov. en el primer registro de polen en el Cenozoico (lapso Paleoceno-Plioceno) de sedimentos argentinos. Los representantes actua les de este género en Argentina son integrantes de la selva higrófila.
Euphorbiaceae. La familia se halla documentada en Argentina con palinomorfos y hojas (ver Tabla 1). De la Tabla 1, se desprende que por primera vez, para el Mio-Plioceno se halla una especie afin a Euphorbia, en estratos argentinos.
Tabla 1. Registros de especies fósiles de Euphorbiaceae de Argentina en el lapso Paleoceno-Plioceno.

En la línea evolutiva sugerida para el género Euphorbia por El-Ghazaly & Chaudhary (1993), la nueva especie fósil (Euphortricolporites perforata), tendría una ubicación basal, lo que ofrece sustento a dicha propuesta filogenética; como así también a la evolución general de la familia en el Gondwana (Martínez Gordillo & Morrone, 2005), confirmando el alto grado de endemismo, especiación activa y escasa dispersión a otras regiones.
En la actualidad, los géneros endémicos de Euphorbiaceae en Argentina están distribuidos mayoritariamente en sabanas y praderas de ambientes xerófilos.
Fabaceae. Los registros fósiles de la familia Fabaceae en Argentina (impresiones de hojas, frutos, leños y palinomorfos) son numerosos y se encuentran detallados en Anzótegui et al. (2007a, cuadro 1, Pág. 135) los que se resumen en la Tabla 2; esos datos están actualizados con los aportados por Anzótegui et al. (2007b), Anzótegui & Horn (2011), Brea et al. (2013), Franco & Brea (2010) y Mautino (2010). Se omitieron los referidos a Prosopis, debido a que están relacionados al género tratado en este trabajo.
Tabla 2. Registro de Fabaceae, en Argentina, durante el Cenozoico. Modificado de Anzótegui et al. 2007 y actualizado con los aportes de Anzótegui et al. (2007b), Anzótegui & Horn (2011), Brea et al. (2013), Franco & Brea (2010) y Mautino (2010). Los tipos de fósiles son agrupados por regiones (tomado y modificado de Barreda et al., 2007).

De esta Tabla se rescata que el registro de Fabaceae en la Patagonia fue continuo desde el Paleoceno hasta el Mioceno temprano, en cambio en Cordillera Frontal, Valles Calchaquíes y Mesopotamia desde el Mioceno Medio hasta el Plioceno. La mayoría de los palinomorfos corresponden a políades (21 sp.), porque ofrecen un diagnóstico seguro para algunos géneros de las tribus Acacieae, Mimoseae e Ingeae de la subfamilia Mimosoideae; el resto de los géneros de la familia contienen granos de polen del tipo tricolporado, carácter muy común en diversas familias de las Magnoliofitas; excepto Margocolporites vanwijhei Germeraad, Hopping & Müller 1968 que siendo tricolporado ha sido asignado a una Caesalpinoideae (Ottone et. al., 1998; Mautino, 2010) por sus colpos con amplias membranas aperturales.
En la Tabla 3 se observa que Prosopis fue registrado solamente en el Mio-Plioceno de los Valles Calchaquíes, Cordillera Frontal y Mesopotamia. A partir del análisis de datos moleculares y análisis filogenéticos, Catalano et al. (2008) proponen que la evolución y diversificación del género Prosopis en el Neógeno fue coincidente con la expansión de las zonas áridas en América, es decir que el control climático fue determinante en la diversificación del grupo. Prosocolporites argentina sp. nov. también está asociado a ambientes xéricos.
Tabla 3. Registro del género fósil Prosopis (Fabaceae) de Argentina en el lapso Mio-Plioceno.

Tabla 4. Registro de especies fósiles de Anacardiaceae de Argentina en el lapso Cretácico tardío-Plioceno.

Anacardiaceae. El registro de Anacardiaceae en el país muestra que en la Patagonia se produjo desde el Cretácico tardío hasta el Mioceno, en tanto que en el centro-norte la mayoría se produjo en el Neógeno (excepto los palinomorfos de las Formaciones Tunal y Maíz Gordo en Salta). En cuanto a los palinomorfos la afinidad con actuales es incierta en la mayoría, siendo esta la primera vez que se logra este objetivo; en tanto que en leños y láminas foliares, es llamativo que solamente se encuentre una referencia a Schinus en la Patagonia (Schinus molleiformis) y ninguna a Lithrea tal vez una revisión a los taxones de láminas foliares (que fueron erigidos sobre un ejemplar y a veces incompletos) y a los palinomorfos (a partir de este trabajo) aporten alguna novedad a estos géneros. Solamente en los Valles Calchaquíes están presentes ambos géneros en láminas foliares.
Las especies de láminas foliares antes mencionadas avalan las afinidades halladas en palinomorfos (Grupo 1 y 2) en el presente trabajo. Es de destacar que 6 especies de Schinus son endémicas (Zuloaga et al., 1999; Muñoz, 2000) en Argentina. Por otra parte Terrazas-Salgado (en Ramírez & Cevallos-Ferriz, 2002), proponen que algunas regiones de América del Sur (Chaco, en Argentina) y México, son postuladas como importantes centros de diversificación debido a su alta diversidad y nivel de endemismo.
Cyperaceae. Cabe aclarar que los palinomorfos vinculados a esta familia tienen caracteres conservadores a lo largo de la columna estratigráfica, que facilitan su determinación. Cyperaceaepollis neogenicus sp. nov. y otras especies indeterminadas aparecen en sedimentos argentinos desde el Oligoceno y a través del Mioceno (Oligoceno-Mioceno: Palamarczuk & Barreda, 2000; Nañez et al., 2009 y Zamaloa, 2000. Mioceno: Barreda & Palamarczuk, 2000a; Palazzesi & Barreda, 2004 y Mautino, 2010, entre otros). La mayoría en la Patagonia y algunos en los Valles Calchaquíes. En las formaciones San José, Chiquimil, Palo Pintado, El Morterito, y Andalhuala (cuyas edades oscilan entre Mioceno medio y Plioceno), se han identificado alrededor de 16 especies de Cyperaceae en impresiones de hojas, tallos, espiguillas y núculas (Anzótegui et al., 2007c). Con respecto al género actual Kyllinga se lo documenta como fósil en porciones de tallos, vainas foliares y láminas en Scirpitis sp. 1 (afín a K. brevifolia) en Formación Chiquimil (Catamarca). Esto demostraría que la familia estaría diversificándose en el Mioceno-Plioceno en el noroeste de nuestro país (Anzótegui, 2004). Kyllinga como la mayoría de las Ciperáceas, está asociada a ambientes palustres dulceacuícolas.
En conclusión, este análisis demuestra que la mayoría de las familias (excepto Boraginaceae) que contienen las especies nuevas aquí estudiadas tuvieron un amplio registro cronoestratigráfico y distribución geográfica en Argentina. No obstante, como Euphorbia, Prosopis, Kyllinga, Schinus y Lithraeae afines actuales de Euphortricolporites perforata, Prosocolporites argentina, Cyperaceaepollis neogenicus y Ailanthipites gamerroi Grupo 1 y Ailanthipites gamerroi Grupo 2, se encuentran en el Neógeno, acompañados por otras especies de sus respectivas familias y considerando el alto grado de endemismo y diversificación de estos géneros en la actualidad, se reafirma una vez más lo que sostienen, entre otros, Anzótegui (2004), Catalano et al. (2008), Martínez Gordillo & Morrone (2005) y Terrazas-Salgado (com. pers. en Ramírez y Cevallos-Ferris, 2002), que en el Neógeno se habría producido la diversificación de esas familias en el sur de América del Sur.
Por otra parte, es de destacar que (entre otras) Cordiapollenites espinulata, Prosocolporites argentina y Ailanthipites gamerroi nov. comb. Grupo 2, acompañadas de Areolipollis pirenii (Acanthaceae) en Mautino, 2011; Sphaeroplea miocenica (zigosporas de "Algas verdes"), Ovoidites sp. 2 (Zignematales) en Mautino, 2007a; Malvacipolloides tucumanensis, Malvapantocolporites rafaelii, Malvapantocolporites sanjosesii, Baumanipollis sp. 2 (todas Malvaceae) en Mautino et al. 2004; son propias de la Formación San José. En tanto que Ailanthipites gamerroi Grupo 1 (Anacardiaceae) y Baumanipollis sp. 3 (Malvaceae) en Mautino et al. (2004), Dictyophyllidites chiquimilense (Matoniaceae o Cyatheaceae) Mautino y Anzótegui, 1998 lo son de la Formación Chiquimil.
Por último se ratifican las paleocomunidades de los taxones estudiados, obtenidas y brindadas previamente en Mautino (2010), es así que la comunidad hidrófila palustre dulceacuícola estaría integrada, entre otros taxones, por Cyperaceaepollis kyllingoides, el bosque higrófilo por Cordiapollenites espinulata junto a Ailanthipites gamerroi Grupo 1 y la comunidad xerófila por Euphorbiatricolporites perforata, Prosocolporites argentina y Ailanthipites gamerroi Grupo 2.
BIBLIOGRAFÍA
1. Anzótegui, L. M. 1998. Hojas de angiospermas de la Formación Palo Pintado, Mioceno superior, Salta, Argentina. Parte I: Anacardiaceae, Lauraceae y Moraceae. Ameghiniana 35 (1): 25-32 [ Links ]
2. Anzótegui, L. M. 2001. Anacardiaceae. En: Pire, S.M., Anzótegui L.M. y G. Cuadrado (Eds). Flora Polínica del Nordeste Argentino 2. EUDENE. UNNE: 19-26. [ Links ]
3. Anzótegui, L.M. 2004. Megaflora de la Formación Chiquimil, (Mioceno Superior), en los valles de Santa María y Villavil, provincias de Catamarca y Tucumán, Argentina. Ameghiniana 41 (3): 303-314. [ Links ]
4. Anzótegui, L.M. 2006. Paleofloras del Mioceno en los Valles Calchaquíes, Noroeste de Argentina. Universidad Nacional del Nordeste. Tesis doctoral. 266 pp. [Inédito]. [ Links ]
5. Anzótegui, L.M. & S.S. Garralla. 1986. Estudio palinológico de la Formación Paraná (Mioceno Superior) (Pozo Josefina, provincia de Santa Fe, Argentina). Parte 1. Descripciones sistemáticas. Facena 6: 101-176. [ Links ]
6. Anzótegui, L.M. & R. Herbst. 2004. Megaflora (hojas y frutos) de la Formación San José en río Seco, departamento Santa María, provincia de Catamarca. Argentina. Ameghiniana 41: 423-436. [ Links ]
7. Anzótegui, L.M., Garralla, S. & R. Herbst. 2007a. Fabaceae de la Formación El Morterito (Mioceno Superior) del valle del Cajón, provincia de Catamarca, Argentina. Ameghiniana 44(1): 183-196. [ Links ]
8. Anzótegui L.M., Horn Y. & R. Herbst. 2007b. Paleoflora (Fabaceae y Anacardiaceae) de la Formación Andalhuala (Plioceno Inferior), provincia de Catamarca, Argentina. Ameghiniana 44(3): 525-535. [ Links ]
9. Anzótegui, L.M., Horn, Y, Mautino, L.R. & S.S. Garralla. 2007c. Diversificación de Cyperaceae en los valles Calchaquíes durante el Mioceno Medio-Plioceno. XXXI Jornadas Argentinas de Botánica: 117. Corrientes. [ Links ]
10. Anzótegui, L.M. & P. Aceñolaza. 2008. Macrofloristic assemblage of the Paraná Formation (Middle-Upper Miocene) in Entre Ríos (Argentina). Neues Jahrbuch für Geologie und Paläontologie- Abhandlungen. 248: 159-170. [ Links ]
11. Anzótegui, L.M. & Y.M. Horn. 2011. Megaflora de la Formación Palo Pintado (Mioceno Superior) Salta, Argentina. Parte II. Revista Brasileira de Paleontología 14(3): 239-254. [ Links ]
12. Araujo, M.M., Longhi S.J., Brena, D.A., Contente de Barros P.L. & S. Franco. 2004. Análise de agrupamento da Vegetaçâo de um fragmento de Floresta estacional decidual aluvial Cachoeira do Sul, RS, Brasil. Ciencia Florestal, Santa Maria 14 (1): 133-147. [ Links ]
13. Archangelsky, S. 1973. Palinología del Paleoceno de Chubut. Parte 1: Descripciones Sistemáticas. Ameghiniana 10(4): 339-399. [ Links ]
14. Baldoni, A.M. 1987. Nuevas descripciones palinológicas en el área de Collón Curá (Terciario inferior) Provincia del Neuquén, Argentina. 6º Congreso Latinoamericano de Paleontología, Bolivia 1: 399-414. [ Links ]
15. Barreda, V.D. 1989. Palinología estratigráfica de las sedimentitas terciarias del "Patagoniano" en los alrededores de la ciudad de Comodoro Rivadavia Provincias de Chubut y Santa Cruz. Tesis doctoral. Universidad de Buenos Aires. Facultad de Ciencias Exactas y Naturales Departamento de Ciencias Biológicas. 361 pp. [Inédito]. [ Links ]
16. Barreda, V.D. 1997a. Palynomorph assemblage of the Chenque Formation Late Oligocene-Miocene from golfo San Jorge basin, Patagonia, Argentina. Part 3. Polycolpate and tricolporate pollen. Ameghiniana 34(2): 131-144. [ Links ]
17. Barreda, V.D. 1997b. Palinoestratigrafía de la Formación San Julián en el Área de Playa La Mina (Provincia de Santa Cruz), Oligoceno de la Cuenca Austral. Ameghiniana 34(3): 283-294. [ Links ]
18. Barreda, V.D. & M.A. Caccavari. 1992. Mimosoideae (Leguminosae) ocurrents in the Early Miocene of Patagonia (Argentina). Paleogeography, Paleoclimatology and Paleoecology 94: 243-252. [ Links ]
19. Barreda, V.D., Gutiérrez, P.R. & C.O. Limarino. 1998. Edad y paleoambiente de la "Serie del Yeso", Valle del Cura, Provincia de San Juan: Evidencias palinológicas. Ameghiniana 35(3): 321-335. [ Links ]
20. Barreda, V.D. & S. Palamarczuk. 2000a. Palinomorfos continentales y marinos de la Formación Monte León en su área tipo, provincia de Santa Cruz, Argentina. Ameghiniana 37(1): 3-12. [ Links ]
21. Barreda, V.D. & S. Palamarczuk. 2000b. Palinoestratigrafía de depósitos de Oligoceno Tardío-Mioceno en el área sur del Golfo San Jorge, provincia de Santa Cruz, Argentina. Ameghiniana 37(1): 103-117. [ Links ]
22. Barros, E. 1947. Cyperaceae. En: H. Descole (ed.) Genera et Species Plantarum Argentinarum. G. Kraft Ltd. Buenos Aires 4 (1): 1-243. [ Links ]
23. Berry, E.W. 1925. Miocene flora from Patagonia. Johns Hopkins University Studies in Geology 6: 183-223. [ Links ]
24. Berry, E.W. 1928. Tertiary fossil plants from the Argentine Republic. Proceedings of the United States National Museum 73: 1-27. [ Links ]
25. Berry, E.W. 1938. Tertiary flora from the Rio Pichileufú, Argentina. Geology Society American. Special 12: 1-149 [ Links ]
26. Biganzoli, F. & M.E. Múlgura de Romero. 2004. Inventario florístico del Parque Provincial Teyú Cuaré y alrededores (Misiones, Argentina). Darwiniana 42(1-4): 1-24. [ Links ]
27. Bossi, G. E. & R. Palma. 1982. Reconsideración de la estratigrafía del Valle de Santa María, Provincia de Catamarca, Argentina. Actas del 5º Congreso latinoamericano de Geología, Buenos Aires, 1982, 1, 155-172. [ Links ]
28. Bossi, G. E., Gavriloff, I. J. C. & G. Esteban. 1998. Terciario Estratigrafía, Bioestratigrafía y Paleogeografía. En: M. Gianfrancisco, N.E. Puchulu, J. Durango de Cabrera y G.F.Aceñolaza (eds.). Geología de Tucumán. Colegio de Graduados en Ciencias Geológicas de Tucumán, Tucumán, 87-108. [ Links ]
29. Brea, M. Aceñolaza, P.G. & A. F. Zucol. 2001. Estudio paleoxilológico en la Formación Paraná, Entre Ríos, Argentina. 11° Simposio Argentino de Paleobotánica y Palinología (Buenos Aires) Asociación Paleontológica Argentina, Publicación especial 8, 7-17. [ Links ]
30. Brea, M. & A. F. Zucol. 2006. Leños fósiles de Boraginaceae de la Formación Peñas Coloradas (Paleoceno Superior) Puerto Visser, Chubut, Argentina. Ameghiniana 43(1): 139-146. [ Links ]
31. Brea, M. & M.J. Franco. 2013. El registro de Anacardiaceae en el Cenozoico Tardío del Nordeste Argentino. Acta Geológica Lilloana 25: 21-36 [ Links ]
32. Brea, M., Zucol, A.F. & M.J. Franco. 2013. Paleoflora de la Formación Paraná (Mioceno), Cuenca Chaco-Paranense, Argentina En: D. Brandoni y J.I. Noriega (eds.). El Neógeno de la Mesopotamia Argentina. Asociación Paleontológica Argentina. Publicación Especial 14: 28-40. [ Links ]
33. Burkart, A. 1976. A monograph of the genus Prosopis (Leguminosae subfam. Mimosoideae). J. Arnold Arbor. 57: 219-249 [ Links ]
34. Burkart, A. 1987. Leguminosae. En N.S. Troncoso y N.M. Bacigalupo (eds.) Flora de Entre Ríos. Colección Científica del Instituto de Tecnología Agropecuaria 6 (3a): 1-763. [ Links ]
35. Cabrera, A. L. 1965. Flora de la Provincia de Buenos Aires. (Argentina). Dicotiledóneas Gamopétalas. (Ericáceas a Caliceráceas). Tomo IV. Parte V. Colección Científica del INTA. Buenos Aires. 434 pp. [ Links ]
36. Cabrera, A.L. 1976. Territorios fitogeográficos de la República Argentina. En L.R.Parodi (ed.). Enciclopedia Argentina de Agricultura y Jardinería. Editorial ACMÉ. 1, 85 pp. [ Links ]
37. Caccavari, M.A. 1996. Analysis of the South American fossil pollen record of Mimosoideae (Leguminosae). Review of Palaeobotany and Palynology 94: 123-135. [ Links ]
38. Caccavari, M. & L.M. Anzótegui. 1987. Polen de Mimosoideae (Leguminosae) de la Formación Ituzaingó, Plioceno superior de Corrientes, Argentina. 4° Congreso Latinoamericano de Paleontología Bolivia, (Bolivia). Actas: 443-458. [ Links ]
39. Caccavari, M. A. & V. Guler. 2002. Una nueva especie de Acaciapollenites del Neógeno de la Argentina. 8° Congreso Argentino de Paleontología y Bioestratigrafía. (Corrientes). Resúmenes: 77. [ Links ]
40. Catalano S.A., Vilardi J.C., Tosto D. & B.O. Saidman. 2008. Molecular phylogeny and diversification. Biological Journal of the Linnean Society 93: 621-640. [ Links ]
41. Cronquist, A. 1981. An Integrated System of Classification of Flowering Plants. Ed. Columbia University Press, 1262 pp. [ Links ]
42. Dong Z. & P. Baas. 1993. Wood anatomy of trees and shrubs from China. V. Anacardiaceae. IAWA Journal 14: 87-102. [ Links ]
43. Dusén, P. 1908. Über die Tertiäre flora der Seymour-Insel. Wissenschaftliche Ergebnisse der Schwedische Südpolar Expedition 1901-1903, Band III, Geologie und Paläontologie, 27 pp. [ Links ]
44. El-Ghazaly, G.A. & R. Chaudhary. 1993. Pollen morphology of some species of the genus Euphorbia L. Review of Palaeobotany Palynology 78: 293–319.
45. Fernández Pacella, L., Mautino, L.R. y L.M. Anzótegui. 2013. Fabaceae-Mimosoideae (Prosopis). En Pire, S.M., Anzótegui L.M. y G. Cuadrado (Eds). Flora Polínica del Nordeste Argentino 4. EUDENE. UNNE: 89-95. [ Links ]
46. Fiori, A. 1931. Filliti terziarie della Patagonia I. Filliti della riva meridionale del lago Nahuel Huapi. Giornale di Geologia (Ann. R. Mus. Geol. Bologna). Ser. II 6: 101-106. [ Links ]
47. Fleig, M. 1987. Anacardiaceae. En: H,M, Longhi Wagner (ed.) Flora ilustrada do Rio Grande do Sul. Boletim Instituto de Biociencias, Porto Alegre 42: 1-72. [ Links ]
48. Franco, M.J. 2009. Leños fósiles de Anacardiaceae en la Formación Ituzaingó (Plioceno-Pleistoceno), Toma Vieja, Paraná Entre Ríos, Argentina. Ameghiniana 46(4): 587-604. [ Links ]
49. Franco, M.J. & M. Brea. 2008. Leños fósiles de la Formación Paraná (Mioceno Medio), Toma Vieja, Paraná, Entre Ríos, Argentina: registro de bosques estacionales mixtos. Ameghiniana 45: 699-718. [ Links ]
50. Franco, M.J. & Brea, M. 2010. Microlobiusxylon paranaensis gen. nov et sp. nov. (Fabaceae-Mimosoideae) from the Pliocene- Pleistocene of Ituzaingó Formation, Paraná Basin, Argentina. Revista Brasileira de Paleontología 13: 103-114. [ Links ]
51. Frenguelli, J. 1953. Flora fósil de la Región del Alto río Chalía en Santa Cruz (Patagonia). Notas del Museo La Plata (Paleontología) 16: 239-257. [ Links ]
52. Galván, A. F. & O.J. Ruiz Huidobro. 1965. Geología del valle de Santa María. Estratigrafía de las formaciones Mesozoico-Terciarias. Acta Geológica Lilloana 7: 217-230 [ Links ]
53. García, M.J. 1994. Palinologia de turfeiras quaternárias do médio vale do Rio Paraíba Do Sul, Estado de Sao Paulo. Tesis doctoral, Universidade de São Paulo, São Paulo. 354 pp. [Inédito]. [ Links ]
54. Gonzalez Guzmán, A. E. 1967. A palynological study on the Upper Los Cuervos and Mirador Formations. (Lower and Middle Eocene; Tibú Area, Colombia). Leiden, E. J Brill. 90 pp. [ Links ]
55. Graham, A. 1976. Studies in Neotropical Paleobotany. 2. The Miocene communities of Veracruz, México. Annals of the Missouri Botanical Garden 63: 787-842. [ Links ]
56. Guinet, P. & M. Bessedik. 1984. Présence du genre Prosopis (Leguminosae-Mimosoideae) a L'Aquitanienbasal Dans L'aude (Languedoc-France). Pollen et spores 26: 101-108. [ Links ]
57. Guler, M.V., Guerstein, G.R. & M. E. Quattrocchio. 2001. Palinología del Neógeno de la Perforación Cx-1 Cuenca del Colorado, Argentina. Revista Española de Micropaleontología 33: 183-204. [ Links ]
58. Horn, M.Y. 2014. Palinofloras de las Formaciones El Morterito y Palo Pintado (Mioceno tardio-Plioceno temprano) noroeste de Argentina. Universidad Nacional de La Plata. Tesis Doctoral, 167pp. [Inédito]. [ Links ]
59. Hünicken, M., 1955. Depósitos Neocretácicos y Terciarios del extremo SSW de Santa Cruz (Cuenca carbonífera de Río Turbio). Revista del Instituto Nacional de Ciencias Naturales (Museo Argentino de Ciencias Naturales "Bernardino Rivadavia") Ciencias Geológicas 4: 1-164. [ Links ]
60. Hünicken, M., 1966. Flora Terciaria de los estratos de Río Turbio, Santa Cruz (Niveles plantíferos del Arroyo Santa Flavia). Serie Ciencias Naturales. Revista de la Facultad de Ciencias Exactas, Físicas y Naturales, Córdoba 56: 139-262. [ Links ]
61. Ibáñez, L. 2001. Análisis paleoambiental de la Formación Chiquimil en el valle de Santa María, Catamarca, Tucumán, Argentina. Universidad Nacional de Tucumán, Tesis Doctoral, 199 p. [ Links ]
62. Kleinert, K. & M. R. Strecker, 2001. Climate change in response to orographic barrier uplift: paleosol and stable isotope evidence from the late Neogene Santa Maria basin, northwestern Argentina. Geological Society of America 6: 728-242. [ Links ]
63. Kurkbride, J.H.; Gum, Ch.R. & A.L. Wetzman. 2003. Fruit and seed of genera in the subfamily Faboideae (Fabaceae). United States Department of Agriculture, 1208 pp. [ Links ]
64. Latorre, C., Quade, J., & Y. McIntosh, 1997. The expansion of C4 grasses and global cange in the table Miocene: stable isotope evidence from the Americas. Earth and Planetary Science Letters. 146: 83-96. [ Links ]
65. Leopold, E.B. 1969. Miocene pollen and spore Flora of Einewetok Atoll, Marshall Island. Geological Survey Professional Paper 260-II: 1133-1201. [ Links ]
66. Lutz, A.I. 1979. Maderas de Angiospermas (Anacardiaceae y Leguminosae) del Plioceno de la Provincia de Entre Ríos, Argentina. Facena 3: 39-63. [ Links ]
67. Lutz, A. I. 1980. Entrerrioxylon victoriensis nov. gen. et sp. (Leguminosae) del Mioceno Superior (Fm Paraná) la provincia de Entre Ríos, Argentina. Facena 4: 21-29. [ Links ]
68. Lutz, A.I. 1987. Estudio anatómico de maderas terciarias del valle de Santa María (Catamarca- Tucumán), Argentina. Facena 7: 125-144. [ Links ]
69. Lutz, A.I. 1991. Descripción anatómica de Mimoxosoxylon sp. del Plioceno (Formación Ituzaingó) de la provincia de Corrientes, Argentina. Rev. Asoc. Cs. Nat. Litoral. 22: 3-10. [ Links ]
70. Marshall, L.G. & Patterson, B. 1981. Geology and geochronology of mamad-bearing Tertiary of Valle de Santa María and rio Corral Quemado, Catamarca province, Argentina. Feldiana, Geology, n.s. 9: 1.80. [ Links ]
71. Martínez, L.C.A. 2010. Prosopisinoxylon anciborae nov. gen. et sp. (Leguminosae, Mimosoideae) from the Late Miocene Chiquimil Formation (Santa María Group), Catamarca, Argentina. Review of Paleobotany and Palynology 158(3-4): 262-271. [ Links ]
72. Martínez Gordillo, M. & J.J. Morrone. 2005. Patrones de Endemismo y disyunción de los géneros de Euphorbiaceae sensu lato: un análisis panbiogeográfico. Boletín de la Sociedad Botánica de México 77: 21-33. [ Links ]
73. Mautino, L.R. 2007a. Chlorophyta de los valles Calchaquíes (Mioceno medio y superior), Argentina. Revista Española de Micropaleontología 39(1-2): 81-102. [ Links ]
74. Mautino L.R. 2007b. Palinomorfos de las Formaciones San José y Chiquimil, Mioceno, Argentina. Novedades en afinidades botánicas. Reunión Anual de Comunicaciones de la Asociación Paleontológica Argentina. 50º Aniversario de Ameghiniana. Corrientes: 15. [ Links ]
75. Mautino, L.R. 2007c. Primer registro de polen de Prosopis (Fabaceae) del Mioceno Medio, Noroeste de Argentina. 23º Jornadas Argentinas de Botánica. Actas: 120. [ Links ]
76. Mautino, L.R. 2010. Palinofloras de las Formaciones San José y Chiquimil (Mioceno Medio y Superior), Noroeste de Argentina. Universidad Nacional del Nordeste. Tesis Doctoral, 444 pp. inédito [ Links ]
77. Mautino, L.R. 2011. Palinología de las Formaciones San José y Chiquimil (Mioceno Medio y Superior) noroeste de Argentina. Especies nuevas. Revista Brasileira de Paleontología 14(3): 279-290. [ Links ]
78. Mautino, L.R. & L.M. Anzótegui. 1998. Palinología de la Formación Chiquimil (Mioceno superior) Localidad Vallecito, Provincia de Catamarca. Parte 1: Especies Nuevas. Ameghiniana 35(2): 227-233. [ Links ]
79. Mautino, L.R. & L.M. Anzótegui. 2000. Esporas del Mioceno y retrabajadas del Mesozoico en la Formación Chiquimil, Vallecito, Provincia de Catamarca. Ameghiniana 37(1): 13-22. [ Links ]
80. Mautino, L. R. & L.M. Anzótegui. 2002a. Palinología de la Formación Chiquimil (Mioceno superior), Vallecito, provincia de Catamarca. Parte 2. Polen. Ameghiniana 39(3): 257-270. [ Links ]
81. Mautino, L.R. & L.M. Anzótegui. 2002b. Palinología de la Formación Chiquimil (Mioceno superior) Vallecito, provincia de Catamarca. Parte 3. Polen. Ameghiniana 39(3): 271- 284. [ Links ]
82. Mautino, L.R., Cuadrado, G. & L.M. Anzótegui. 2004. Novedades taxonómicas, diversidad y significado evolutivo del polen de Malvaceae en el terciario de Argentina. Revista Española de Micropaleontología 36: 467-483. [ Links ]
83. Milne, L.A. 1988. Palynology of a Late Eocene lignitic sequense from the western margin of the Eucla Basin, Western Australia. Memoir of the Association of Australasian Palaeontologist 5: 285-310. [ Links ]
84. Muller, J., Schuller M., Straka H. & B. Friedrich. 1989. Palynologia Madagassic et Mascarenica. Trop. Subtrop. Pflanzenwelt 67. 225 pp. [ Links ]
85. Muñoz, J. De Dios. 1990. Anacardiaceae. Flora del Paraguay.14. Conservatoire Botanique de Genève and Missouri Botanical Garden: 84pp. [ Links ]Muñoz, J.
86. Muñoz, J. De Dios. 2000. Anacardiaceae. Flora Fanerogámica Argentina. Fascículo 65. ProFlora: 28 pp [ Links ]
87. Nañez, C., Quattrocchio M.E. & L. Ruiz. 2009. Palinología y microaleontología de las Formaciones San Julián y Monte León (Oligoceno-Mioceno temprano) en el subsuelo de cabo Curioso, provincia de Santa Cruz, Argentian. Ameghiniana 46(4): 669-693. [ Links ]
88. Nowicke, J. W. & J.S. Miller 1990. Pollen morphology of the Cordioideae (Boraginaceae): Auxeinina, Cordia, and Patagonula. Plants Systematic and Evolution (Suppl. 5): 103-121. [ Links ]
89. Nowicke J.W. & J.E. Ridgway. 1973. Pollen Studies in the genus Cordia (Boraginaceae). American Journal of Botany 60(6): 584-591. [ Links ]
90. Ottone, E. G., Barreda, V.D. & D.J. Pérez. 1998. Basin evolution as reflected by Miocene palynomorphs from the Chinches formation, frontal cordillera (32ºS), San Juan province, Argentina. Revista Española de Micropaleontología 30(3): 35-47. [ Links ]
91. Palamarczuk, S. & V.D. Barreda. 2000. Palinología del Paleógeno tardío-Neógeno temprano, pozo Aries x-1, Plataforma Continental Argentina, Tierra del Fuego. Ameghiniana 37(2): 221-234. [ Links ]
92. Palazzesi, L. & V.D. Barreda. 2004. Primer registro palinológico de la Formación Puerto Madryn, Mioceno de la provincia de Chubut, Argentina. Ameghiniana 41(3): 355-362. [ Links ]
93. Piel K.M. 1971. Palynology of Oligocene sediments from central British Columbia. Canadian Journal of Botany 49: 1885-1920. [ Links ]
94. Prámparo, M.B., Papú, O.H. & J.P. Milana. 1996. Estudio palinológico del Miembro inferior de la Formación Pachaco, Terciario de la provincia de San Juan. Descripciones sistemáticas. Ameghiniana 33(4): 397-407. [ Links ]
95. Pujana, R.R. 2010. Una nueva Mimosoideae (Leguminosae) de la Formacion Salicas (Mioceno), provincial de La Rioja, Argentina. X Congreso Argentino de Paleontología y Bioestratigrafía. VII Congreso Latinoamericano de Paleontología: 203. [ Links ]
96. Quattrocchio, M. & L.C. Ruiz. 1999. Paleoambiente de la Formación Pedro Luro (Maastrichtiano?-Paleoceno) en base a palinomorfos, cuenca del Colorado , Argentina. Ameghiniana 36(1): 37-47. [ Links ]
97. Quattrocchio, M. & C. Del Papa. 2000. Paleoambiente de la secuencia Maiz Gordo (¿Paleoceno Tardío-Eoceno Temprano?), Arroyo Las Tortugas, Cuenca del Grupo Salta (NOArgentina), Palinologia y sedimentología. Revista Española de Micropaleontologia 15(1): 57-66. [ Links ]
98. Ramírez, J. L. & S.R.S., Cevallos-Ferriz 2002. A diverse assemblage of Anacardiaceae from Oligocene sediments, Tepexi de Rodriguez, Puebla, Mexico. American Journal of Botany 89: 535-545. [ Links ]
99. Romero, E.J. & M.T. Castro.1986. Material fúngico y granos de polen de angiospermas de la formación Río Turbio (Eoceno), provincia de Santa Cruz, República Argentina. Ameghiniana 23: 101-118. [ Links ]
100. Ruiz, L. & M.T. Quattrocchio. 1996. Stratigraphic palynology of Pedro Luro Formation (?Maastrichtian-Paleocene), Colorado Basin. Argentine. Elf Aquitaine. Mémoire 16: 361-371. [ Links ]
101. Scheel, R. & O.M. Barth. 1995. Catálogo sistemático do pólen das plantas arbóreas do Brasil Meridional. 29: Boraginaceae. Revista Brasileira de Biologia 55(4): 769-776. [ Links ]
102. Scheel, R., Ybert, J-P. & O.M. Barth. 1996. Pollen morphology of the Boraginaceae from Santa Catarina State (southern Brazil), with comments on the taxonomy of the family. Grana 35: 138-153. [ Links ]
103. Spegazzini, C. 1924. Sobre algunas impresiones vegetales Eocénicas de Patagonia. Comunicaciones del Museo Nacional de Historia Natural de Buenos Aires 2: 95-107 [ Links ]
104. Sung, T. & M. Lee. 1976. Characteristics of late Cretaceous to early Tertiary pollen and spores of Yunnan. En Acad. Sin., Nanjing Inst. Geol. Paleontol., Yunnan Province, Mesozoic Fossils, 1: 1-64. [ Links ]
105. Tressens, S.G., Vanni R.O. & M.G. López. 2002. Las plantas terrestres del macrosistema Iberá. En Arbo & Tressens, Flora del Iberá, Ed. EUDENE, Corrientes, Argentina: 201-379 [ Links ]
106. Wodehouse, R.P. 1933. Tertiary pollen. II. The oil shales of the Eocene Green River formation. Bull. Torrey Botanical Club 60: 479-524. [ Links ]
107. Ybert, J.P. 1979. Atlas de pollens de Cõte D'Ivoire. Office de la Recherche Scientifique et Technique Outre-mer. 40 pp. [ Links ]
108. Zamaloa, M. del C. 2000. Palinoflora y ambiente en el Terciario del nordeste de Tierra del Fuego, Argentina. Revista del Museo Argentino Ciencias Naturales Bernardino Rivadavia 2: 43-51. [ Links ]
109. Zamaloa, M. del C. & R.R. Andreis. 1995. Asociación palinológica del Paleoceno Temprano (Formación Salamanca) en Ea. Laguna Manantiales, Santa Cruz, Argentina. 6º Congreso Argentino de Paleontología y Bioestratigrafía, Actas: 301-305. Trelew. [ Links ]
110. Zetter, R.C.C., Hofmann, I. Draxler; J. Durango de Cabrera, Vergel, M.M. & F. Vervoorst. 1999. A rich Middle Eocene microflora at Arroyo de los Mineros, near Cañadón Beta, NE Tierra del Fuego Province, Argentina. Abhandlungen der Geologischen Bundesanstalt 56: 438-460. [ Links ]
111. Zuloaga, F.O., Morrone, O. & D. Rodríguez. 1999. Análisis de la biodiversidad en plantas vasculares de la Argentina. Kurtziana 27: 17-167. [ Links ] [ Links ]
Recibido: 30-X-2013
Aceptado: 4-IX-2014

