










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
versión On-line ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.17 no.1 Ciudad Autónoma de Buenos Aires jun. 2015
PALEONTOLOGÍA
Primer registro de Cryptocaryoxylon Leisman, una Lauraceae de la Formación El Palmar (Pleistoceno tardío), Entre Ríos, Argentina
R. Soledad Ramos1, Mariana Brea1 & Daniela M. Kröhling2
1Laboratorio de Paleobotánica, Centro de Investigaciones Científicas y Transferencia de Tecnología a la Producción (CICYTTP-CONICET), Dr. Materi y España SN, E3105BWA, Diamante, Entre Ríos, Argentina, laresole@hotmail.com, cidmbrea@gmail.com
2CONICET y Universidad Nacional del Litoral, Facultad de Ingeniería y Ciencias Hídricas CC217, 3000 Santa Fe, Argentina, dkrohling@santafe-conicet.gov.ar
Resumen
Este trabajo describe el primer registro de Cryptocaryoxylon Leisman del Pleistoceno tardío de Argentina. El ejemplar fue hallado en el Paraje Salto Grande (Cuenca del Río Uruguay, Entre Ríos, Argentina) en sedimentos fluviales de la Formación El Palmar. Los caracteres diagnósticos del material fósil son: Porosidad difusa a semicircular; vasos pequeños a medianos y numerosos, solitarios y múltiples radiales cortos; tilosis abundante; punteaduras radio-vasculares simples; parénquima axial vasicéntrico, aliforme y confluente; radios heterocelulares, uniseriados, biseriados raros, altos; fibras no septadas con paredes gruesas y presencia de numerosas células oleíferas en los extremos de radios. Las características anatómicas sugieren una relación con Cryptocarya (Lauraceae). Las características eco-anatómica son consistentes con un clima templado-cálido y húmedo durante este intervalo de tiempo.
Palabras clave: Cryptocaryoxylon sp. nov.; Lauraceae; Formación El Palmar; Pleistoceno tardío; Entre Ríos; Argentina.
Abstract
First record of Cryptocaryoxylon Leisman, a Lauraceae from the El Palmar Formation (late Pleistocene), Entre Ríos, Argentina
This paper described the first record of Cryptocaryoxylon Leisman from the late Pleistocene of Argentina. The specimen was recovered in Paraje Salto Grande locality (Uruguay River Basin, Entre Ríos, Argentina) in fluvial sediments of the El Palmar Formation. The diagnostic features of the fossil specimen are: diffuse to semi-ring porous wood; small to medium and numerous vessels, solitary and in short radial multiples; tyloses abundant; vessels-ray parenchyma pits simple; axial parenchyma vasicentric, aliform and confluent; heterocellular rays, uniseriate and rarely biseriate, high; non-septate fibres with thick-walled, and numerous "oil cells" in rays. These anatomical characteristics suggest a relationship with Cryptocarya (Lauraceae). The eco-anatomical characteristics are consistent with a temperate-warm and humid climate during this time interval.
Key words: Cryptocaryoxylon sp. nov.; Lauraceae; El Palmar Formation; Late Pleistocene; Entre Ríos; Argentina.
INTRODUCCIÓN
Lauraceae es una familia de origen pantropical con unas pocas especies distribuidas en áreas templadas, como es el caso de los bosques esclerófilos de Chile (Serra et al., 2005). Comprende 50 géneros con aproximadamente 2750 especies. En la región neotrópical existen 27 géneros y al menos 1000 especies (van der Werff & Rohwer, 1999). La Argentina cuenta con 5 géneros y 16 especies (Tressens, 1997), la mayoría nativas (Jozami & Muñoz, 1982; Hadid de Cheda, 1991) entre ellas el género Cryptocarya, el cual está constituido por una única especie, C. ashersoniana (Tressens, 1997).
Las familias Lauraceae, Magnoliaceae y Annonaceae integran el orden Laurales (Magnoliidae) caracterizado por ser un grupo con caracteres plesiomórficos respecto a otros grupos de plantas con flores (APG III, 2009). Esos caracteres menos derivados incluyen también a la anatomía de la madera en el sentido de Bailey (conocido como the Baileyan trends), entiéndase por caracteres plesiomórficos a los leños secundarios que presentan vasos solitarios, placas de perforación escalariformes, radios heterocelulares y parénquima apotraqueal (Baas et al., 2000).
Los registros más antiguos de Lauraceae se conocen a partir de restos florales de Mauldinia mirabilis Drinnan, Crane, Friis y Pedersen 1990 del Cretácico medio de los Estados Unidos (Drinnan et al., 1990) y desde el Cenozoico se registran fósiles preservados como impresiones foliares, polen, frutos y semillas en todo el mundo (Li et al., 2004; Taylor et al., 2009). La madera fósil de Lauraceae tiene un amplio registro a nivel mundial. Dupéron-Laudoueneix & Dupéron (2005) analizaron los registros de la familia proponiendo cambios nomenclaturales para muchos de los taxones conocidos. Entre los más importantes se menciona a Beilschmiedioxylon Dupéron-Laudoueneix & Dupéron 2005; Caryodaphnopsoxylon Gottwald 1992; Cinnamomoxylon Gottwald 1997; Cryptocaryoxylon Leisman 1986; Laurinoxylon Felix 1883; Machilusoxylon (Bande) Ingle 1974; Mezilaurinoxylon Wheeler & Manchester 2002; Paraperseoxylon Wheeler & Manchester 2002; Paraphyllanthoxylon Bailey 1924; Sassafrasoxylon Brezinová & Süss 1988; Richteroxylon Wheeler & Dillhoff 2009.
El registro de Lauraceae en la Mesopotamia Argentina se conoce a partir del estudio de cutículas con afinidad a Nectandra Rol. ex Rottb. y Ocotea Aubl. Estos taxones fueron hallados en la Formación Ituzaingó (Plio-Pleistoceno), en la localidad de Riachuelo, provincia de Corrientes, Argentina (Anzótegui, 1980). Recientemente, se describieron dos leños fósiles de lauráceas para la misma formación, Laurinoxylon artabeae (Brea) Dupéron-Laudoueneix & Dupéron 2005 y Curtiembrexylon poledrii Franco 2012 afines a Ocotea y Beilschmiedia Nees. respectivamente. Estos ejemplares provienen de las localidades de Hernandarias y Curtiembre, provincia de Entre Ríos, Argentina (Franco, 2012).
La Formación El Palmar hasta el momento contaba con dos géneros y tres especies fósiles de Lauraceae: Laurinoxylon artabeae (Brea) Dupéron-Laudoueneix & Dupéron y L. mucilaginosum (Brea) Dupéron-Laudoueneix & Dupéron (Brea, 1998), hallados en las localidades fosilíferas Punta Viracho (Departamento Concordia) y Arroyo Caraballo (Departamento Colón) respectivamente y Beilschmiedioxylon parataubertiana Ramos, Brea y Kröhling (Ramos et al., 2012), proveniente de la localidad fosilífera Parque Nacional El Palmar, integran la lista paleoflorística del Pleistoceno tardío de la Formación El Palmar, que aflora en el sector medio de la Cuenca del Uruguay en la provincia de Entre Ríos.
El objetivo del presente trabajo es dar a conocer una nueva especie de Cryptocaryoxylon sp. nov., proveniente de los depósitos fluviales de la Formación El Palmar, hallado en el Paraje Salto Grande, Departamento de Federación, Entre Ríos, Argentina (Fig. 1).
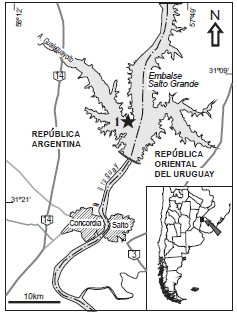
Fig. 1. Ubicación geográfica de la localidad fosilífera Paraje Salto Grande, Departamento Federación, Entre Ríos, Argentina.
Marco geológico
La Formación El Palmar, definida formalmente por Iriondo (1980) y caracterizada en detalle por Iriondo & Kröhling (2008), es una faja de acumulación fluvial vinculada a la margen derecha del río Uruguay que abarca ca. 15 Km de ancho. Está representada por una llanura aluvial abandonada, compuesta por depósitos de canal de alta energía y sedimentos asociados de inundación, y constituye la terraza alta del río Uruguay a lo largo de ca. 1000 km, desde Chapecó en Brasil (27°S) hasta Concepción del Uruguay en la frontera Argentina-Uruguay (32°30'S). La terraza es discontinua en la alta cuenca del río Uruguay y es, en su sector medio, donde alcanza buen desarrollo y continuidad abarcando el este de la provincia de Entre Ríos, Argentina (Iriondo & Kröhling, 2008; Kröhling, 2009). La Formación El Palmar, en este último sector, se extiende sobre la margen izquierda del Río Uruguay, a la República Oriental del Uruguay, donde es conocida como Formación Salto (Bossi, 1969; Iriondo, 1980; Veroslavsky & Montaño, 2004).
Las dataciones por termoluminicencia (localidades: Federación, Entre Ríos y Salto, Uruguay) de la Formación El Palmar indican edades entre 80 y 88 ka. AP para los sedimentos de la parte superior aflorante, representada por arenas de cauce (Iriondo & Kröhling, 2008; Kröhling, 2009). Las características sedimentológicas, estratigráficas y las dataciones absolutas de esta unidad sugieren que la misma se habría generado durante el Estadio Isotópico Marino del Oxígeno EIO5a, correspondiente a uno de los interestadiales cálidos a nivel global del Cuaternario superior (Kröhling, 2009).
MATERIALES Y MÉTODOS
El material fósil fue colectado en el Paraje Salto Grande (31˚ 12' 57.02"S, 57˚ 55' 41.07"O), que se ubica al noreste del antiguo curso del Arroyo Gualeguaycito, actualmente localizado dentro del embalse originado por la construcción de la represa hidroeléctrica de Salto Grande, Departamento Federación, Entre Ríos, Argentina (Fig. 1). El material fue recuperado en la década del 70 por Cristina Vassallo de Cettour, directora del Museo de Antropología y Ciencias Naturales de la ciudad de Concordia, en sedimentos de la Formación El Palmar.
El ejemplar permineralizado por silicificación sólo ha preservado la anatomía del xilema secundario. Las características exomorfológicas, el desgaste superficial y el tamaño de la muestra, permiten inferir que es de origen paraautócotono, la porción de leño mide 12 cm de largo y 9 cm de diámetro. Para su estudio se realizaron los cortes petrográficos en sus tres secciones estándares (corte transversal, longitudinal tangencial y longitudinal radial). El material fue estudiado con un microscopio óptico Nikon Labophot y Nikon Eclipse E200 y las microfotografías fueron tomadas con una cámara digital Nikon Coolpix S4.
Los valores cuantitativos de las descripciones anatómicas constituyen promedios de 30 medidas, en todos los casos se cita primero los valores promedios y entre paréntesis, las medidas mínimas y máximas de su rango de distribución.
Para su descripción se utilizaron los glosarios de términos de la Asociación Internacional de Anatomistas de Maderas IAWA Committee (1989), Kribs (1935), Tortorelli (1956), Carlquist (2001) y las clasificaciones estándar de Chattaway (1932). También se incluyeron las recopilaciones bibliográficas de Gregory (1994) y el catálogo de Gregory et al. (2009). Para la identificación del material fósil se utilizaron los atlas, las bases de datos disponibles en la web y las descripciones de Metcalfe & Chalk (1957) Richter (1987), Watson & Dallwitz (1992), Richter & Dallwitz (2000) e InsideWood database (InsideWood, 2004).
Se siguió la clasificación sistemática del APG III (2009). Para los nombres de las especies actuales y fósiles se utilizaron el International Plants Names Index (The Plant List, 2010) y el Index Nominum Genericorum (ING) (Farr & Zijlstra, 1996). El Holotipo se encuentra depositado en la Colección Paleobotánica del Museo de Antropología y Ciencias Naturales de Concordia (Entre Ríos, Argentina), bajo la sigla MACNC-Pb Nº 7546 y el isotipo en la Colección del Laboratorio de Paleobotánica CICyTTP-CONICET, Diamante, bajo los acrónimos CIDPALBO-MEG 131 y CIDPALBO-MIC1554.
SISTEMÁTICA PALEOBOTÁNICA
Cryptocaryoxylon Leisman 1986
Especie tipo: Cryptocaryoxylon gippslandicum Leisman 1986
Cryptocaryoxylon oleiferum sp. nov.
(Fig. 2. 1-13)

Fig. 2. Cryptocaryoxylon oleiferum sp. nov. Ramos, Brea y Kröhling. (CIDPALBO-MIC1554). A, Corte transversal (CT), distribución de los vasos y del parénquima axial; B, Corte longitudinal tangencial (CLT), disposición de los radios y los elementos de vasos; C, (CT) detalle de vasos dispuestos en serie múltiples radiales, la mayoría con depósitos oscuros, parénquima axial vasicéntrico escaso, trayecto rectilíneo de los radios y anillo de crecimiento; D, Corte longitudinal radial (CLR), radios heterocelulares y disposición de los numerosos idioblastos secretores "células oleíferas o mucilaginosas" (flecha); E, (CT) detalle de vasos solitarios, múltiples radiales, parénquima axial vasicéntrico aliforme y confluente y radios delgados (flecha); F, (CT) detalle de anillo de crecimiento (flecha) y disposición de fibras; G, (CLT) detalle de elementos de vasos con tabique oblicuo (flecha blanca), radios biseriados, uniseriados, con células agregadas y células "oleíferas" en el extremo del radio (flecha negra); H, (CLT) detalle de fibras no septadas y radio uniseriado; I, (CLR) detalle de células procumbentes de radio y células oleíferas (flecha); J, (CLT) detalle de radios uniseriados heterocelulares con cristales prismáticos (flecha); K, (CT) detalle de células oleíferas (flecha), fibras y parénquima axial confluente; L, (CLR) detalle de células procumbentes con vestigios de punteaduras radio-vasculares simples(flecha). Escala gráfica: A = 500 μm, B, C= 200 μm; D, E, G = 100 μm; F, H-K = 20 μm; L = 15 μm.
Holotipo. MACNC-Pb 7546.
Isotipo. CIDPALBO-MEG 131, CIDPALBO-MIC1554 (tres preparados microscópicos).
Procedencia geográfica. Paraje Salto Grande (31˚ 12' 57,02"S, 57˚ 55' 41,07"O), Entre Ríos, Argentina.
Procedencia estratigráfica. Formación El Palmar (Pleistoceno tardío).
Afinidad botánica. Familia Lauraceae, con una marcada afinidad al género actual Cryptocarya R. Br.
Derivatio nominis. El epíteto específico, oleiferum, se refiere a los abundantes idioblastos secretores probablemente de origen oleífero o mucilaginoso.
Diagnosis. Porosidad difusa a semicircular. Vasos pequeños a medianos y numerosos. Vasos solitarios y múltiples radiales cortos. Placa de perforación simple. Tilosis abundante. Punteaduras radio-vasculares simples. Parénquima axial vasicéntrico, aliforme y confluente. Radios heterocelulares uniseriados, raros biseriados. Fibras con paredes gruesas. Los extremos de radios presentan células oleíferas.
Diagnosis. Diffuse to semi-ring porous wood. Vessels small to medium and numerous. Vessels solitary and in short radial multiples. Simple perforation plates. Tyloses abundant. vessels-ray parenchyma pits simple. Axial parenchyma vasicentric, aliform and confluent. Heterocellular rays, uniseriate and rarely biseriate. Fibres with thick-walled. Oil cells associated with ray parenchyma.
Descripción. La descripción e identificación está basada en un ejemplar fósil (MACNC-Pb 7546). En corte transversal, los anillos de crecimientos están poco demarcados por compresión radial de las fibras, por parénquima axial inicial y por la disposición de los vasos. Porosidad difusa a semicircular, sin patrón de distribución (Fig. 2. A). Vasos solitarios (45%), múltiples radiales de 2-3 (47%) y de 4 hasta 7 elementos (8%), de contorno oval. Diámetro tangencial promedio de 76 (43-114) μm y diámetro radial promedio de 90 (38-140) μm, pequeños a medianos. Espesor promedio de la pared de 9 (7-13) μm, delgadas. Tilosis abundante y depósitos oscuros en el lumen de los vasos (Fig. 2. A, C, E, K). Densidad de vasos promedio de 28 (21-38) por mm², numerosos. Radios numerosos, de 13 (12-16) por mm lineal (Fig. 2. A, E). Parénquima axial paratraqueal vasicéntrico escaso, aliforme a confluente cuando los vasos están muy próximos (Fig. 2. A, C, E). Fibras ovales a angulares con distribución en hileras radiales, diámetro promedio de 12 (7-18) μm, paredes delgadas a gruesas con un espesor promedio de 4 (2-5) μm (Fig. 2. F, K).
En corte longitudinal tangencial, elementos de vasos con trayecto rectilíneo, longitud promedio de los miembros de vasos de 278 (220-320) μm (Fig. 2. B, G). Punteaduras intervasculares ovales, medianas y alternas, diámetro promedio de 9 (7-10) μm (Fig. 2. G). Placas de perforación simple de tabiques oblicuos (Fig. 2. G). Radios heterogéneos, generalmente uniseriados (90%), biseriados o uniseriados con células agregadas (10%), con un ancho promedio de 22 (20-42) μm y una altura promedio de 298 (190-483) μm y de 15 (4-28) células de altos (Fig. 2. B, H, J). Fibras no septadas (Fig. 2. H).
En corte longitudinal radial, los radios heterocelulares están constituidos por células procumbentes y células erectas o cúbicas en el cuerpo, mientras que en los extremos se observan células oleíferas de forma globosa que miden entre 25-50 μm de ancho (Fig. 2. D, I, L). Las punteaduras radio-vasculares son simples, similar a las punteaduras intervasculares siendo del tipo clase b de la clasificación de Richter (1981, 1987) (Fig. 2. I, L).
Comparación con especies actuales
Las Lauraceae se distingue por poseen la siguiente combinación de caracteres: vasos solitarios, múltiples radiales cortos, presencia de tilosis, placas de perforación simples, punteaduras intervasculares alternas, punteaduras radio-vasculares generalmente similares a las puntuaciones radios vasculares en forma y tamaño, parénquima axial escaso y vasicéntrico, radios predominantemente de 2-4 células de ancho, ocasionalmente uniseriados, homocelulares o heterocelulares, fibras septadas y no septadas e idioblastos secretores, gran parte de estos caracteres se compara con el material estudiado (Patel, 1987; Richter, 1987; Wheeler et al., 1987; Callado & Costa, 1997; Dupéron-Laudoueneix & Dupéron, 2005; León, 2000a, b, c, 2001; León & Espinosa de Pernía, 2000a, b; Parra, 2009). En la Tabla 1 se muestran los caracteres anatómicos de los géneros más afines a Cryptocaryoxylon oleiferum sp. nov.
Tabla 1. Comparación entre Cryptocaryoxylon oleiferum sp. nov. y los géneros actuales más afines (datos tomados de 1981, Metcalfe, 1987, Patel, 1987, Callado & Costa, 1997, León, 2000 a, b, 2001; Ramos et al., 2012) Referencias: Vasos MR= múltiples radiales, S = solitarios, = diámetro tangencial, PP = placas de perforación, PI = punteaduras intervasculares; Fibras S = septadas, NS = no septadas; P = presente, R= raro, O= ocasional; PA = parénquima axial, V = vasicéntrico, A = aliforme, C= confluente, M = marginal, = difuso disyunto; He = heterocelulares, Ho = homocelulares; Cristales P = presentes, A = ausentes; PRV = punteaduras radiovasculares, oleíferas = radios, F = fibras.
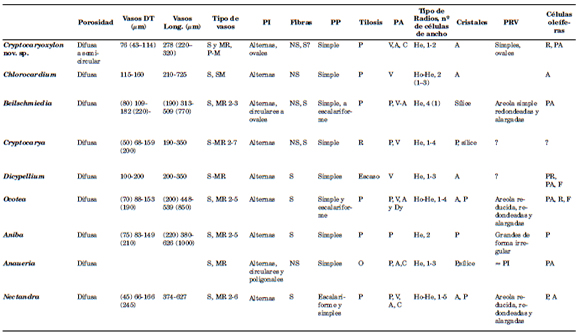
El ejemplar fósil estudiado comparte caracteres con los géneros Ocotea, Nectandra, Nees & Mart., Dicypellium Nees & Mart., Beilschmiedia y Cryptocarya. Los caracteres que distinguen a Ocotea del material estudiado son la porosidad de tipo difusa, las fibras septadas y los radios que son homocelular y heterocelular con 1 a 4 células de ancho. Nectandra se diferencia del ejemplar fósil por el predominio de las fibras septadas, radios pocos numerosos, con 2-4 células de ancho (Tortorelli, 1956). Con Dicypellium se diferencia por presentar fibras septadas y en la seriación de los radios. Beilschmiedia se diferencia del material estudiado en el tamaño de los vasos, que son de medianos a grandes (entre 80 y 220 μm de diámetro) y en menor densidad, radios predominantemente de 2 a 3 células de ancho y/o multiseriados, los uniseriados constituidos por células erectas o cuadradas (León, 2000c, 2001).
El material estudiado se relaciona estrechamente con Cryptocarya caracterizado por poseer radios heterocelulares de dos tipos, uniseriados y biseriados, siendo los primeros más frecuentes en las especies que integran el género, punteaduras intervasculares del tipo clase b de la clasificación de Richter (1981, 1987), con 8-12 μm de diámetro y puntuación radio-vasculares de contorno variable, circulares a ovales, a veces alargadas y con disposición horizontal, vertical y/o diagonal. La frecuencia de vasos en cadenas radiales es inusual en las Lauraceae, pero son comunes en Cryptocarya kurzii Hook. f. (Richter, 1981; Wheeler & Manchester, 2002), carácter que comparte con el espécimen en estudio. El material fósil comparte con Cryptocarya los siguientes caracteres diagnósticos: porosidad semi-circular a difusa, vasos dispuestos en series múltiples radiales, pequeños a medianos, radios comúnmente uniseriados y presencia de idioblastos secretores oleíferos asociados al parénquima radial.
En América del Sur se citan ca. 10 especies de Cryptocarya, distribuidas en Brasil, Chile, Guayana Francesa, Venezuela, Ecuador y Perú. El material fósil se comparó con las especies que cuentan con descripciones del xilema secundario. Cryptocarya alba (Molina) Looser se diferencia de Cryptocaryoxylon oleiferum sp. nov. en la presencia de cristales en radios y tilosis esclerosada (Díaz Vaz, 1979; Rancusi et al., 1987). Cryptocarya manii Hillebr. se diferencia por tener parénquima difuso y ausencia de células mucilaginosas (Detienne & Jacquet, 1983), y con C. aschersoniana Mez., única especie presente en Argentina, se diferencia por la ausencia de células mucilaginosas y el predominio de radios con más de 3 células de ancho (Richter, 1981, Metcalfe, 1987, Patel, 1987, Callado & Costa, 1997, León, 2000a, b, c, 2001).
Comparación con especies fósiles
En el presente trabajo se siguieron los criterios propuestos por Dupéron et al. (2008) respecto a la sinonimia de Laurinium Unger 1845 y Ulminium Unger 1842 a Laurinoxylon Felix 1883. El material estudiado fue comparado con los géneros fósiles de Lauraceae que presentan radios heterocelulares (Tabla 2). De este análisis se desprende que presenta sus mayores semejanzas con Cryptocaryoxylon caracterizado por presentar: vasos con placas de perforación simples, radios homocelulares y heterocelulares con hasta 5 células de ancho y menos de 1 mm de alto, parénquima axial escaso, idioblastos secretores (células oleíferas) generalmente adyacentes a los radios (o en sus extremos), o dispersas entre las fibras; siendo Cryptocarya el género actual con más afinidad (Richter, 1981; Metcalfe, 1987; Wheeler & Manchester, 2002).
Tabla 2. Comparación entre los géneros fósiles más afines de Lauraceae con Cryptocaryoxylon oleiferum sp. nov. (datos tomados de Leisman, 1986, Nishida et al., 1990, Dupéron-Laudoueneix & Dupéron, 2005, Ruiz Estrada-Martínez et al., 2010; Ramos et al., 2012). = diámetro tangencial, P = pequeños, M = medianos; PP = placa de perforación; PI = punteaduras intervasculares; Radios He = heterogéneos, Ho = homogéneos; Fibras S = septadas, NS = no septadas; Células oleíferas en PA = parénquima axial, PR = parénquima radial, F = fibras, = radios

Las características destacadas del espécimen fósil que comparte con las especies de Cryptocaryoxylon son: el tipo de disposición de los vasos y la presencia de células oleíferas en radios. En la Tabla 3 se muestran las semejanzas y diferencias entre las especies fósiles de Cryptocaryoxylon.
Tabla 3. Comparación de Cryptocaryoxylon oleiferum sp. nov. con especies de Cryptocaryoxylon (datos tomados de Leisman, 1986; Wheeler & Manchester, 2002) Referencias Vasos MR= múltiples radiales, S = solitarios, DT= diámetro tangencial, P = pequeños, M = medianos, PP = placas de perforación, PI = punteaduras intervasculares; PR= punteaduras radio-vasculares (según clasificación de Richter, 1987); Fibras S = septadas, NS = no septadas; PA = parénquima axial, V = vasicéntrico, A = aliforme, C= confluente, M = marginal, B = bandeado, T= terminal; Radios He = heterocelulares; CM = células mucilaginosas en R= radios, F = fibras; Cristales N= sin cristales, ? = sin dato.
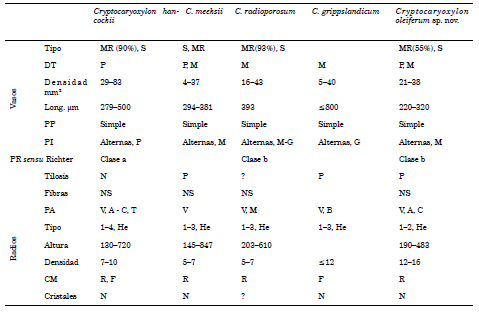
Cryptocaryoxylon oleiferum sp. nov. se asemeja a C. grippslandicum (Wheeler & Manchester, 2002) en la disposición y características de los vasos, la posición de las células oleíferas y en el tipo de radios, pero se diferencia en la presencia de punteaduras intervasculares de tamaño grandes (≥10 μm) y parénquima axial bandeado. Mientras que Cryptocaryoxylon meesksii difiere en la altura y en la densidad por mm lineal de los radios.
La combinación de caracteres diagnósticos (predominio de vasos dispuestos en series múltiples radiales, radios uniseriados, escasos biseriados, heterocelulares y células oleíferas en los extremos de los radios) presentes en el material estudiado no permiten ubicarlo dentro de una especie fósil conocida. Por este motivo se propone, la creación de una nueva entidad específica denominada Cryptocaryoxylon oleiferum sp. nov.
CONSIDERACIONES FINALES
La estructura anatómica de las Lauraceae es considerada muy uniforme. Sin embargo, la combinación de sus características anatómicas las diferencia de las otras familias (Richter, 1987). Muchos géneros de lauráceas no están bien definidos y algunas especies que pertencen a un género no siempre comparten los caracteres anatómicos. Estas dificultades usualmente están relacionadas con identificaciones taxonómicas poco precisas o caracteres diagnósticos faltantes de un ejemplar en una descripción. Además, se debe tener en cuenta cuales son los caracteres constantes y cuales son influenciados por el medio ambiente (Metcalfe & Chalk, 1957; Metcalfe, 1987).
La nueva especie, Cryptocaryoxylon oleiferum sp. nov., constituye el primer registro fósil afín a Cryptocarya registrado en el noreste de Argentina. La combinación única de los caracteres anatómicos diagnósticos la vincula claramente dentro de este género, siendo los caracteres más importantes la presencia de vasos en series múltiples radiales largas, los radios heterocelulares, predominantemente uniseriados y la abundancia de idioblastos secretores en los extremos de los radios (Sonnerat, 1782; León, 2001; León & Espinoza de Pernía, 2000a, b).
Los datos moleculares han señalado que los géneros Beilschmiedia y Cryptocarya divergen de su ancestro común más reciente a los 90 ±20 Ma., lo que indica también una migración directa a lo largo del supercontinente de Gondwana y una distribución de pre-deriva generalizada de ambos géneros (Li et al., 2004; Serra et al., 2005).
La presencia de especies fósiles afines a Beilschmiedia (Ramos et al., 2012) y Cryptocarya (este trabajo) en la Formación El Palmar avalarían la historia evolutiva y fitogeográfica de ambos géneros, incluyendo la hipótesis de un área tropical a lo largo de gran parte de Sudamérica que se interrumpiría con el levantamiento final de los Andes durante el Mio-Plioceno y que habría provocado la expansión de la gran diagonal árida, produciendo de este modo un aislamiento de especies en sentido noreste y suroeste del continente. Como consecuencia de estos acontecimientos, Beilschmiedia y Cryptocarya, tienen en la actualidad una distribución disyunta, encontrándose especies en los bosques neotropicales y en los bosques patagónicos subtropicales-templados de Chile y Argentina (Villagrán & Hinojosa, 1997; León, 2001; Moraes, 2007).
Los caracteres anatómicos fuertemente influenciados por el ambiente son el número de vasos por mm2, el largo y diámetro de los elementos de vaso, el porcentaje de vasos agrupados, la porosidad, el tipo de placas de perforación y la proporción de parénquima axial (Wheeler & Baas, 1993; Callado & Costa, 1997; Woodcock et al., 2000). Cryptocaryoxylon oleiferum sp. nov. se caracteriza por presentar porosidad difusa a semicircular, con una densidad baja de 28 vasos por mm2, vasos con diámetro pequeño a mediano (76 μm), con un alto porcentaje de vasos agrupados (55%), con elementos de vasos cortos (278 μm), con placas de perforación simples y parénquima axial paratraqueal vasicéntrico, aliforme y confluente. Todos estos caracteres probablemente han contribuido en buena medida a la eficiencia en el sistema de conducción del agua y una adaptación tendiente a la seguridad hidráulica (Wheeler & Baas, 1991, 1993).
El Índice de Vulnerabilidad (V) de Cryptocaryoxylon oleiferum sp. nov. es de 2,71 y el Índice de Mesomorfía (M) de 753, estos valores evidencian desarrollo de caracteres mesomórficos (Carlquist, 1977, 2001). Los caracteres eco-anatómicos y los Índices de V y M en concordancia con las interpretaciones geológicas (Kröhling, 2009) permiten inferir un clima tropical húmedo durante la depositación de gran parte de la Formación El Palmar (Kröhling, 2009).
AGRADECIMIENTOS
Los autores desean expresar su agradecimiento a la Directora del Museo de Ciencias Naturales y Antropología de Concordia (Entre Ríos, Argentina), Cristina Vassallo de Cettour, quien recuperó el ejemplar fósil en el Paraje Salto Grande, permitió el acceso a la colección científica bajo su cuidado y generosamente dispuso el préstamo del material fósil para su estudio. Se extiende el agradecimiento a los revisores L.C.A. Martínez y E. Ruiz-Estrada por sus valiosos comentarios que permitieron mejorar la versión final del manuscrito. A Amalia González por su ayuda con el mapa de ubicación. Este trabajo fue financiado por la Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT) PICT2008 N° 0176. El presente trabajo se realizó durante la Carrera de Doctorado de Ciencias Biológicas de la Universidad Nacional de Córdoba de una de las autoras (RSR).
BIBLIOGRAFÍA
1. APG III. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society 161: 105-121. [ Links ]
2. Anzótegui, L.M. 1980. Cutículas del Terciario superior de la provincia de Corrientes, República Argentina. Actas II Congreso Argentino de Paleontología y Bioestratigrafía y I Congreso Latinoamericano de Paleontología 3: 141-167. [ Links ]
3. Baas, P., E. Wheeler & M. Chase. 2000. Dicotyledonous wood anatomy and the APG system of angiosperm classification. Botanical Journal of the Linnean Society 134: 3-17. [ Links ]
4. Bailey, I.W. 1924. The problem of identifying the wood of Cretaceous and later dicotyledons: Paraphyllanthoxylon arizonense. Annals of Botany 38: 439-451. [ Links ]
5. Bossi, J. 1969. Geología del Uruguay. Colección Ciencias 12. 2da Edición. Departamento de Publicaciones de la Universidad de la República, Montevideo, 464 pp. [ Links ]
6. Brea, M. 1998. Ulminium mucilaginosum n. sp. y Ulminium artabeae n. sp., Dos leños fósiles de Lauraceae en la Formación El Palmar, provincia de Entre Ríos, Argentina. Ameghiniana 35: 193-204. [ Links ]
7. Brezinova, D. & H. Süss. 1988. Nadel- und Laubholzreste aus miozanen Hornsteinen von Lipnice, CSSR. Feddes Repert 99: 279-289. [ Links ]
8. Callado, C. & C. Costa. 1997. Wood anatomy of some Anaueria and Beilschmiedia species (Lauraceae). IAWA Journal 18: 247-259. [ Links ]
9. Carlquist, S. 1977. Ecological factors in wood evolution: a floristic approach. American Journal of Botany 64: 887-896. [ Links ]
10. Carlquist, S. 2001. Comparative Wood Anatomy. Systematic, ecological, and evolutionary aspects of dicotyledon wood, 2nd edition. Springer-Verlag, Berlin. 407 pp. [ Links ]
11. Chattaway, M. 1932. Proposed standards for numerical values used in describing woods. Tropical Woods 29: 20-28. [ Links ]
12. Díaz Vaz, J. E. 1979. Clave para la identificación de Maderas de árboles nativos y cultivados en Chile. Bosques 3: 15-25. [ Links ]
13. Drinnan, A., P. Crane, E. Friis & K. Pedersen. 1990. Lauraceous flowers from the Potomac Group (Mid-Cretaceous) of eastern North America. Botanical Gazette 151: 370-384. [ Links ]
14. Detienne, P. & P. Jacquet. 1983. Atlas d'identification des bois de l'amazonie et des regions voiseines. Centre Technique Forestier Tropical. Montpellier, France. 279 pp. [ Links ]
15. Dupéron-Laudoueneix, M. & J. Dupéron. 2005. Bois fossiles de Lauraceae: nouvelle découverte au Cameroun, inventaire et discussion. Annales de Paléontologie 91: 127-151. [ Links ]
16. Dupéron, J., M. Dupéron-Laudoueneix, J. Sakala & D. De Franceschi. 2008. Ulminium diluviale Unger: historique de la découverte et nouvelle étude. Annales de Paléontologie 94: 1-12. [ Links ]
17. Farr, E.R. & G. Zijlstra. 1996. Index Nominum Genericorum (Plantarum). [octubre 2013]. http://botany.si.edu/ing/ [ Links ]
18. Felix, J. 1883. Untersuchungen über fossile Hölzer. I. Zeitschrift der Deutschen Geologischen Gesellschaft 35: 59-92. [ Links ]
19. Franco, J. 2012. Maderas fósiles de Lauraceae de la Formación Ituzaingó (Plioceno-Pleistoceno), cuenca del río Paraná, Argentina. Revista del Museo Argentino de Ciencias Naturales 14: 307-324. [ Links ]
20. Gottwald, H. 1992. Hölzer aus Marinen Sanden des Oberen Eozän von Helmstedt (Niedersachsen). Palaeontographica B 225: 27-103. [ Links ]
21. Gottwald, H. 1997. Alttertiäre Kieselhölzer aus miozänen Schotten ostbayerischen Melasse bei Ortenberg. Documenta naturae 109: 1-83. [ Links ]
22. Gregory, M. 1994. Bibliography of systematic wood anatomy of dicotyledons. IAWA Journal supplement, 1: 1-265. [ Links ]
23. Gregory, M., I. Poole & E. Wheeler. 2009. Fossil dicot wood names, an annotated list with full bibliography. IAWA Journal Supplement, 6: 1-220 pp. [ Links ]
24. Hadid de Cheda, M. 1991. Lauraceae Juss Flora del Valle de Lerma. Aportes Bot. Salta, sér. Flora 1 (4): pp. 1-7, Salta, Argentina. [ Links ]
25. IAWA Committee. 1989. List of microscopic features for hardwood identification. IAWA Bull. n.s. 10: 219-332. [ Links ]
26. Ingle, S.R. 1978. Revision of the genus Machilusoxylon Bande. Botanique 5(1): 53-56. [ Links ]
27. InsideWood. 2004. Inside Wood. http://insidewood.lib.ncsu.edu/search [Abril 2013]. [ Links ]
28. Iriondo, M.H. 1980. El Cuaternario de Entre Ríos. Revista de la Asociación de Ciencias Naturales del Litoral 11: 125-141. [ Links ]
29. Iriondo, M.H. & D. Kröhling. 2008. Cambios ambientales en la cuenca del Uruguay (desde el Presente hasta dos millones de años atrás). Colección Ciencia y Técnica. Universidad Nacional del Litoral, Santa Fe, 360 pp. [ Links ]
30. Jozami, J.M. & J. de D. Muñoz. 1982. Árboles y arbustos indígenas de la Prov. de Entre Ríos. IPNAYS-CONICET-UNL, Santa Fe, 407 pp. [ Links ]
31. Kribs, D.A. 1935. Salient lines of structural specialization in the wood rays of dicotyledons. Botanical Gazette 96: 547-557. [ Links ]
32. Kröhling, D.M. 2009. La Formación El Palmar, una unidad fluvial asignable al subestadio cálido EIO5a (Pleistoceno Tardío) de la cuenca del río Uruguay. Natura Neotropicalis 40: 61-86. [ Links ]
33. Leisman, G.A. 1986. Cryptocaryoxylon gippslandicum gen. et sp. nov., from the Tertiary of eastern Victoria. Alcheringa 10: 225-234. [ Links ]
34. León, W. 2000a. Anatomía del leño de 17 especies del género Ocotea Aublet. Pittieria 29-30: 53-65. [ Links ]
35. León, W. 2000b. Estudio Anatomía del leño de ocho especies del género Aniba Aublet (Lauraceae). Revista forestal de Venezuela 44: 37-46. [ Links ]
36. León, W. 2000c. Estudio Anatomía del leño de siete especies del género Beilschmiedia Nees (Lauraceae). Revista forestal de Venezuela 44: 47-56. [ Links ]
37. León, W. 2001. Anatomía del leño de 85 especies de la familia Lauraceae Juss. Facultad de Ciencias Forestales y Ambientales, Universidad de Los Andes, Mérida, 114 pp. [ Links ]
38. León, W. & N. Espinoza De Pernía. 2000a. Estudios anatómicos del leño de ocho especies del género Aniba Aublet (Lauraceae). Revista Forestal Venezolana 44: 37-46. [ Links ]
39. León, W. & N. Espinoza de Pernía. 2000b. Estudios anatómicos del leño de siete especies del género Beilschmiedia Nees (Lauraceae). Revista Forestal Venezolana 44: 47-36. [ Links ]
40. Li, J., D. Christophel, J. Conran & H. Li. 2004. Phylogenetic relationships within the ‘core' Laureae (Litsea complex, Lauraceae) inferred from sequences of the chloroplast gene matK and nuclear ribosomal DNA ITS regions. Plant Systematics and Evolution 246: 19-34.
41. Metcalfe, C.R. 1987. Anatomy of the dicotyledons. Ed. 2, Vol. III. Magnoliales, Illiciales and Laurales. Clarendon Press, Oxford, 224 pp. [ Links ]
42. Metcalfe, C.R. & L. Chalk. 1957. Anatomy of the Dicotyledons. Volumen I. Clarendon, Oxford, 724 pp. [ Links ]
43. Moraes, P.L. de. 2007. Taxonomy of Cryptocarya Species of Brazil. Abc Taxa Series 3: 1-191. [ Links ]
44. Nishida, M., T. Ohsawa & H. Nishida. 1990. Anatomy and affinities of the petrified plants from the Tertiary of Chile (VI). The Botanical magazine, Tokyo 103: 255-268. [ Links ]
45. Parra, J. 2009. Anatomía del leño e identificación de siete especies de Laurales en San Eusebio (Mérida, Venezuela). Pittieria 33: 59-77. [ Links ]
45. Patel, R. 1987. Wood anatomy of the dicotyledons indigenous to New Zealand 16. Lauraceae. New Zealand Journal of Botany 25: 477-488. [ Links ]
46. Rancusi, M.H., M. Nishida & H. Nishida. 1987. Xylotomy of the important Chilean woods. En: M. Nishida (Ed.), Contributions to the Botany in the Andes II. 68-153. Academia Scientific Book Inc., Tokyo. [ Links ]
47. Ramos, R.S., M. Brea & D. M. Kröhling. 2012. Leños fósiles de la Formación El Palmar (Pleistoceno Tardío) en el parque Nacional El Palmar, Entre Ríos, Argentina. Ameghiniana 49: 606-622. [ Links ]
48. Ritcher, H. 1981. Wood and bark anatomy of Lauraceae. Aniba Aublet. IAWA Bulletin n.s. 2: 79-87. [ Links ]
49. Richter, H. 1987. Lauraceae. Mature Secondary Xylem. En: C. Metcalfe (ed.), Anatomy of the Dicotyledons 2 Edition. Vol. 3, pp. 167-171, Oxford Science Publications, Oxford. [ Links ]
50. Richter, H.G. & M.J. Dallwitz. 2000. Commercial timbers: descriptions, illustrations, identification, and information retrieval. En English, French, German, and Spanish. [versión: 18 de Octubre 2002]. http://delta-intkey.com [ Links ]
51. Serra, M.T., F. Luebert & M. Ricci. 2005. Nuevo registro para Avellanita bustillosii Phil. (Euphorbiaceae). Noticiario Mensual Museo Nacional de Historia Natural Chile 355: 11-15. [ Links ]
52. Sonnerat, P. 1782. Voyage aux Indesorientaleset à la Chine, fait par ordre duroi, depuis 1774 jusqu'en 1781. Danslequel on traité des moeurs, de la religion, des sciences & des arts des Indiens, des Chinois, des Pégouins & des Madégasses;suivid'observationssur le cap de Bonne-Espérance, les isles de France & de Bourbon, les Maldives, Ceylan, Malacca, les Philippines et les Moluques, & derecherchessurl'histoirenaturelles de ces pays, à Paris: chez l'auteur, Froulé, Nyon, Barrois le jeune, 2: 1-298. [ Links ]
53. Taylor, T.N., E.L. Taylor & M. Krings. 2009. Paleobotany. The Biology and Evolution of Fossil Plants. Second Edition. Academic Press, Elsevier, 1230 pp. [ Links ]
54. The Plant List. 2010. Version 1. http://www.theplantlist.org/ [Septiembre, 2014]. [ Links ]
55. Tressens, S.G. 1997. El género Cryptocarya (Lauraceae) en Argentina. Bonplandia 9: 209-212. [ Links ]
56. Tortorelli, L.A. 1956. Maderas y Bosques Argentinos. Editorial ACME, S.A.C.I., Buenos Aires, 910 pp. [ Links ]
57. Unger, F. 1842. Synopsis lignorum fossilium plantarum acramphibryarum. En: S. Endlicher (Ed.), Genera Plantarum. Secundum Ordines Naturales Disposita. Supplement II, Appendix, 100-102. Apud Fridericum Beck, Universitatis Bibliopolam, Vienna, Austria. [ Links ]
58. Unger, F. 1845. Synopsis plantarum fossilium. Leipzig, 330 pp. [ Links ]
59. van der Werff, H. & J.G. Rohwer. 1999. Lauraceae. En: Berry, P.E., K. Yatskievych & B.K. Holst (eds.), Flora of the Venezuelan Guayana, Vol 5: Eriocaulaceae-Lentibulariaceae, pp. 700-750, Missouri Botanical Garden, Missouri. [ Links ]
60. Veroslavsky, G. & J. Montaño. 2004. Sedimentología y estratigrafía de la Formación Salto (Pleistoceno). En: G. Veroslavsky, M. Ubilla y S. Martínez (eds.), Cuencas sedimentarias de Uruguay: geología, paleontología y recursos naturales, pp. 147-166, DI-RAC-Facultad de Ciencias, Cenozoico, Montevideo. [ Links ]
61. Villagrán, C. & F. Hinojosa. 1997. Historia de los bosques del sur de Sudamérica, II: Análisis fitogeográfico. Revista Chilena de Historia Natural 70: 241-267. [ Links ]
62. Watson, L. & M.J. Dallwitz. 1992. The families of flowering plants: descriptions, illustrations, identification, and information retrieval. [23 de Septiembre 2013]. http://delta-intkey.com [ Links ]
63. Wheeler, E., M. Lee & L.C. Matten. 1987. Dicotyledonous woods from the Upper Cretaceous of southern Illinois. Botanical Journal of the Linnean Society 95: 77-100. [ Links ]
64. Wheeler, E. & P. Baas. 1991. A survey of the fossil record for dicotyledonous wood and its significance for evolutionary and ecological wood anatomy. IAWA Bulletin n.s. 12: 275-332. [ Links ]
65. Wheeler, E. & P. Baas. 1993. The potentials and limitations of dicotyledonous wood anatomy for climatic reconstructions. Paleobiology 19: 487-498. [ Links ]
66. Wheeler, E. & S.R. Manchester. 2002. Woods of the Eocene Nut Beds Flora, Clarno Formation, Oregon, USA. IAWA Journal 3: 1-188. [ Links ]
67. Wheeler, E. & T.A. Dillhoff. 2009. The Middle Miocene Wood Flora of Vantage, Washington, USA. IAWA Journal, Supplement 7: 1-101 pp. [ Links ]
68. Woodcock, D.W., G. Dos Santos & C. Reynel. 2000. Wood characteristics of Amazon forest types. IAWA Journal 21: 277-292. [ Links ]
Recibido: 8-I-2015
Aceptado: 20-V-2015

