










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
versión On-line ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.18 no.2 Ciudad Autónoma de Buenos Aires dic. 2016
BOTÁNICA
Proposals for quantifying two characteristics of tracheid pit arrangement in gymnosperm woods
Roberto Román Pujana1, Daniela Paula Ruiz1, Leandro Carlos Alcides Martínez2 & Yi Zhang3
1 Museo Argentino de Ciencias Naturales-CONICET, Ciudad de Buenos Aires, Argentina.
2 Instituto de Botánica Darwinion (ANCEFN-CONICET), San Isidrio, Argentina.
3 College of Paleontology, Shenyang Normal University; Laboratory of Evolution of Past Life in Northeast Asia, Ministry of Land and Resources, Shenyang, China.
Abstract
Arrangement of tracheid radial pits is frequently superficially mentioned in extant gymnosperm wood anatomy descriptions. However, pit arrangement is a key character for delimiting fossil-genera and fossil-species in gymnosperm woods. We propose two new indices for measuring and quantifying the radial pit arrangement in fossil and extant woods. The first one, contiguity percentage index (Cp), indicates if the pits are contiguous (=in contact or touching) or overlapping with the adjacent upper and lower pits, and has values from 0 to 100%. The second one, seriation index (Si), is an average of the seriation of the pits and is given as a numerical value equal or greater than 1; when pits are exclusively ordered in uniseriate columns the wood has a value of 1. These indices are useful for quantifying the pit arrangement and for helping to characterize the type of radial pitting (araucarian/ mixed/abietinean), particularly in fossil woods. These new indices also allow to express these two characters (contiguity and seriation) as continuous, making them more suitable for certain taxonomic or statistical analysis.
Key words: Conifers; Wood anatomy; Fossils; Tracheid pit arrangement.
Resumen:
Propuestas para cuantificar dos características de la disposición radial de las punteaduras en maderas de gimnospermas
La disposición de las punteaduras en las paredes radiales es frecuentemente mencionada superficialmente en descripciones de maderas actuales de gimnospermas. Sin embargo, la disposición de las punteaduras es un caracter fundamental para delimitar los géneros y especies fósiles en maderas de gimnospermas. Proponemos dos nuevos índices para medir y cuantificar la disposición de las punteaduras en maderas fósiles y actuales. El primero, índice del porcentaje de la contigüidad (Cp) indica si las punteaduras son contiguas (=en contacto o tocándose) o solapadas con las punteaduras superiores e inferiores adyacentes y tiene valores del 0 al 100%. El segundo, índice de seriación (Si) es la media de la seriación de las punteaduras y es un valor númerico igual o mayor a 1; cuando las punteaduras son exclusivamente uniseriadas tiene un valor de 1. Estos nuevos índices son útiles para cuantificar la disposición de las punteaduras y para ayudar a caracterizar el tipo de punteaduras radiales (araucarioides/mixtas/abietinoides), particularmente en maderas fósiles. Asimismo, tales índices permiten expresar estos dos caracteres (contigüidad y seriación) como continuos, lo cual es más adecuado para ciertos análisis taxonómicos o estadísticos.
Palabras clave: Coníferas; Anatomía de maderas; Fósiles; Disposición de punteaduras radiales.
INTRODUCTION
Cell wall is an ancestral condition in plants, and most of plant cells are enclosed by walls, except some cells concerned with reproductive, or early ontogeny, processes (Kenrick & Crane, 1991; Beck, 2010). Cell wall is modified in different ways as the cell matures, and some of these changes imply extension, thickening, and modification of grosser physical structure. Most xylem cells are highly specialized in conducting water, and for this reason they have anatomical features of interest in physiology, ecology, evolution and systematic of vascular plants (Carlquist, 1975). Walls of secondary xylem cells have many characters of systematical and/or evolutionary significance: e.g. tracheid pit arrangement, seriation of tracheid pits, presence of torus or helical thickenings, type of cross-fields, etc. (IAWA committee, 2004; García-Esteban, 2002; Greguss, 1955, 1972). Pits are cavities in the wall of a plant cell where there is no secondary wall and where water and sap flow. Their arrangement varies through the different species of gymnosperms. Particularly, pit arrangement is often superficially mentioned in extant gymnosperm (or softwood) wood anatomical descriptions. However, the pit arrangement, which includes contiguity (or spacing) of the pits and seriation, is a key character for delimitation of fossil-genera and fossilspecies in gymnosperm fossil woods (Eckhold, 1922; Vogellehner, 1967; Philippe & Bamford, 2008). IAWA committee (2004) code has two characters related to radial pit arrangement, the first one is “predominantly uniseriate” vs. “predominantly biseriate or more” (a discrete character), and the second one is “biseriate or more seriate pitting, alternate” vs. “opposite” (also a discrete character). Neither of the two refers to the contiguity of the pits, and they are both discrete binary characters.
Nicol (1834) first mentioned the division of radial pitting into araucarian and abietinean (without using those terms). Later Kräusel (1917) and Eckhold (1921, 1922) divided fossil wood pit arrangements into three types: araucarian, mixed and abietinean, making emphasis on the mixed type of fossil woods. Mixed (also called generalized or transitional) is a transitional type of pitting between araucarian pitting (which is not exclusive of the Araucariaceae) and currently more common abietinean radial pitting. According to Bamford et al. (2016), this type of pitting is particularly common in Mesozoic woods. Woods with this type of radial pitting (mixed) were grouped into the “Protopinaceen” (=Protopinaceae) by Kräusel (1917) and this definition was followed by different authors (e.g. Eckhold, 1922; Vogellehner, 1967). Recently it was discussed by Bamford et al. (2016) and they recommended not to use this artificial family (Protopinaceae). A fourth type of pit arrangement was described by Müller-Stoll (1951) as xenoxylean, where radial pits are much flattened, more than twice as wide as high, and contiguous with neighbouring pits (Müller Stoll, 1951; Philippe & Bamford, 2008). This division of radial pitting is currently used for delimiting fossil genera (Vogellehner, 1967; Philippe & Bamford, 2008). However, methods for measuring or classifying the radial pits into these three types vary among different authors and they usually assigned the fossil woods to one of these categories without further details (Philippe et al., 2014). The contiguity of the pits plays a key role in these three types of radial pit arrangement. The seriation of radial pits (uniseriate, biseriate, etc.) is also a character used for fossil species delimitation (e.g. Penhallow, 1907; Pujana et al., 2014, 2015). Usually seriation is given as percentages of uniseriate, biseriate, triseriate, etc. pitting or a vague description is mentioned (“predominantly uniseriate”, “mostly biseriate”, etc.).
In addition, several authors have claimed that it is essential that anatomical characters are quantified (Falcon-Lang & Cantrill, 2000; Poole & Cantrill, 2001) and continuous characters are preferable and more informative for certain statistical or taxonomical analysis rather than discrete characters. For these reasons, we propose two new indices to measure with precise indications the contiguity and seriation of pitting of gymnosperm woods with circular or hexagonal tracheid pits (not scalariform or transitional pitting).
MATERIAL AND METHODS
Studied extant woods (one slide per species) are from the Xylarium of the Museo Argentino de Ciencias Naturales (acronym BA) and of the Facultad de Ciencias Agrarias y Forestales, UNLP (acronym UNLP). Fossil woods measurements were based on the holotypes of each fossil-species. Fossils are housed in the Museo Provincial Padre Molina (acronym MPM PB), Museo Argentino de Ciencias Naturales (acronym BA Pb) and Facultad de Ciencias Naturales y Museo, UNLP (acronym LPPB). A representation of the different radial pit arrangements was attempted.
Measurements of radial pit contiguity and seriation in different species of both fossil and extant gymnosperm woods were taken (Table 1, Fig. 1-4). For obtaining the two indices, measurements were taken in random different zones (including earlywood and latewood) of the slide (or different zones of the fragment observed with scanning electron microscopy). When it was possible, up to 60 horizontal lines of pits were measured for both indices in each zone. We define a horizontal line of pits as indicated in Fig. 2. In uniseriate rows a horizontal line of pits is a single pit, in biseriate rows are two pits, in triseriate rows are three pits and so on (Fig. 2, 4).
Table 1. Examples of measurements of extant (1-9) and fossil woods (10-15).
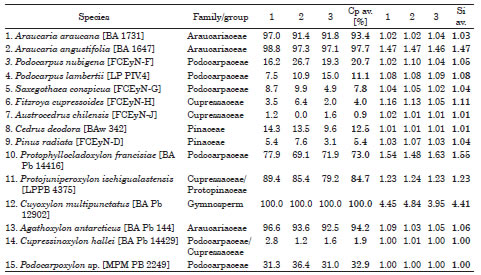
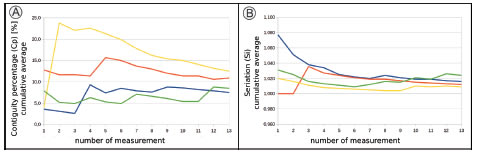
Fig. 1. Podocarpous lambertii [LP PIV.4] measurements. Number of zone measurement vs. cumulative average. Each line represents measurements of each author (4), the 13 zones of the wood are the same for all authors, but the order of the measurements is not the same. A. Cp. Note that after c. 8-10 zone measurements, average is stabilized. B. Si. Note that after c. 5 zone measurements, average is stabilized.

Fig. 2. Hypothetical examples of Cp (contiguity percentage) index measurements and equations. Note that biseriate (or multiseriate) horizontal lines are counted the same as uniseriate. Dashed lines indicate some horizontal lines of pits.

Fig. 3. Hypothetical examples of Si (Seriation index) measurements and equations. Dashed lines indicate some horizontal lines of pits.
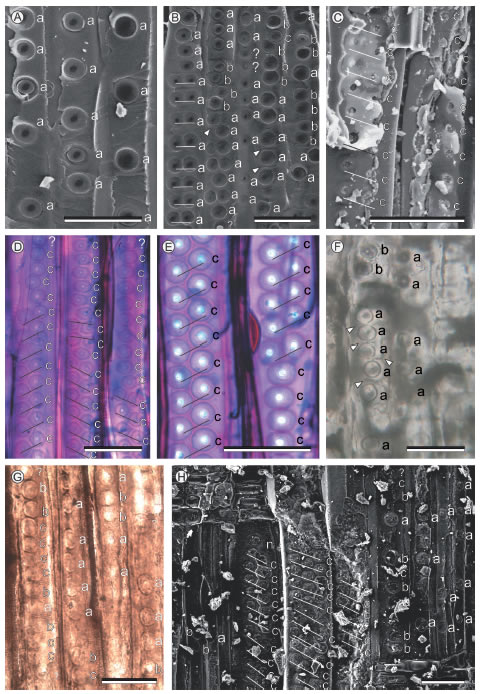
Fig. 4 (Previous page). Samples of radial pit arrangement for calculating Cp. Cp shown below is for each zone and not the Cp calculated for the species in Table 1. c=two touching walls, b=one touching wall, a=no touching walls, n=not counted. A. Austrocedrus chilensis (D. Don) Pichi Sermolli & Bizzarri [ (Cp=0%) [BA FCEyN-J]. B. Podocarpus parlatorei Pilger (Cp=14.8%) [BA FCEyN-A]. C. Agathoxylon sp. (Cp=100%) [BA Pb 14548]. D. Araucaria angustifolia (Cp=100%) [BA 1647]. E. Araucaria angustifolia (Cp=100%) [BA 1647]. F. Podocarpoxylon mazzoni Petriella (Cp=7.7%) [LPPB 9064]. G. Podocarpoxylon sp. (Cp=38.5%) [MPM PB 2249]. H. Protophyllocladoxylon francisiae (Cp=65.6%) [BA Pb 14416]. Bar for all images: 50 μm.
Pits should be counted in random different zones (provided with pits) of the wood to allow the observation of the specimen variation. Figures 2 and 3 show examples of hypothetical typical gymnosperm woods. Note that in those figures the indices were calculated in only one zone of the sample for the purpose of illustrating the methods.
INDICES
Contiguity percentage index (Cp)
The Cp is a percentage (0-100%) that characterizes whether the horizontal lines of pits have the upper and lower outer borders of the pits contiguous (=in contact or touching) or overlapping with the outer borders of the pits of neighbouring (lower and an upper) horizontal lines (Fig. 2). A Cp with a 0% value indicates that all the horizontal lines of pits are spaced, and a 100% value indicate that all the horizontal lines of pits are contiguous with the neighbouring horizontal lines of pits. Horizontal lines of pits must be contiguous, flattened or overlapped to be counted positively. Sometimes pits appear to be spaced, if the outer border of the pit is not clearly observed (see Bamford et al., 2016). In Figure 4D-E a typical Araucaria Jussieu wood with contiguous and flattened pits is shown. Note that the pits are apparently not contiguous, but they are even flattened and hexagonal indicating that they are contiguous. In addition, sometimes only the aperture of the pits is observed and it could be confused with spaced pits (Fig. 4C). Contiguous horizontal lines of pits have uniform distances among them (and the distance between the centers of the pits is the diameter of the pit) while in spaced (also called scattered) pits this distance is variable (Bamford et al., 2016).
The first horizontal line of pits next to a ray or at the end or beginning of a tracheid (which could have only lower or upper border in contiguity at most) should not be counted to allow a maximum of 100% contiguous pits (e.g. Fig. 4H). When crassulae (=bars of Sanio) are present between two horizontal lines of pits we consider them spaced (Fig. 4B, F). Horizontal lines of pits must be measured as in Figure 2, and they should be divided into 3 types: a. Both outer border of pits (upper and lower) separated from other lines of pits. b. Only one outer border, lower or upper, in contact with another line of pits. c. Upper and lower outer borders in contact with the neighbouring horizontal lines of pits. Following the equation in Figure 2, the Cp can be calculated. Note that Cp refers to a single specimen and not to a single tracheid. Consequently, we should count the state of the total horizontal lines of pits in the sample/specimen, and then calculate the Cp.
Seriation index (Si)
The Si index is an average of the seriation of the pits (Fig. 3). It can have a minimum of 1 (all horizontal lines of pits are uniseriate) and a maximum equal to the number of vertical rows (=columns) which a multiseriate pitting tracheid has. Variation in the same tracheid is common; biseriate pitting portions in mostly uniseriate pitting tracheid and vice versa are commonly found, for this reason we count individual horizontal lines of pits and not tracheids as a whole. In this index, we also count horizontal lines of pits and not single pits. Horizontal lines of pits are multiplied by one (when uniseriate), by 2 (when biseriate), by 3 (when triseriate) and so on and then a sum of them is calculated. To calculate the Si we finally divide the sum by the total number of horizontal lines we counted (Fig. 3). Si has no units. Converting previously descriptions of pit seriation is straightforward: for example, to convert a specimen with 68% biseriate lines, 7% triseriate lines and 25% uniseriate lines, Si equals ((1*25+2*68+3*7)/100)=1.82.
Notes
We determined the minimum number of zones to be measured based on successive zone measurements in mature wood of Podocarpus lambertii Klotzsch ex Endlicher. The cumulative average is sufficiently stabilized after approximately 8-10 measurements in contiguity percentage index (Cp) (Fig. 1A) and after approximately 5 measurements in seriation index (Si) (Fig. 1B). Therefore, we suggest that a minimum of 10 zones is preferable to calculate the indices. Both indices were calculated using the total number of horizontal lines of pits of one sample and not calculating a weighted average of the zones (although this latter can also be carried out if a significant different number of horizontal lines of pits in each zone are measured).
Other indices
In fossil wood literature more indices or methods to define radial pitting were used to quantify radial pitting. These can complement the Cp and Si:
1) “Coefficient d’écrasement” or pit flattening coefficient measure individual pit flattening, dividing the height by the width of each pit. It is used sporadically (e.g. Serra, 1966; Marguerier & Woltz, 1977; Desplats, 1979; Vozenin-Serra & Grant Mackie, 1996; Torres et al., 1994).
2) “Percentage touching pits” (Falcon-Lang & Cantrill, 2000) the ratio of tracheids with mainly touching, contiguous pitting to those bearing pits usually spaced more than one pit diameter apart. The main difference with Cp is that “percentage touching pits” method gives higher percentages because pits spaced with more than one pit diameter are more difficult to find, particularly in fossil woods. Some cases are not explained in detail, for example, whether biseriate rows should be counted as uniseriate rows.
3) “Contiguity value” (=“pit contiguity”) (Falcon-Lang & Cantrill, 2001) the length of continuous sequences of touching bordered pits on the radial tracheid walls expressed as the range and mean number of pits in each contiguous sequence.
4) Henderson & Falcon-Lang (2011) give the “mean of pit [vertical] rows”, which we supposed is very similar to Si. However, they do not give an explanation of how they calculated it, whether they measured whole tracheids, horizontal lines of pits or individual pits.
5) Philippe et al. (2014) assigned each pit observed to either one of the six types: UD (uniseriate distant); UC (uniseriate contiguous and usually flattened); BA (biseriate alternate); BOR (biseriate opposite round); BOS (biseriate opposite square). This is a very detailed method to describe radial pitting. However, as each type has a percentage, six different characters should be considered.
DISCUSSION
Radial pit arrangement is crucial for fossil wood genera/species delimitation. An agreement among authors must be found and followed for delimitation of fossil-genera and terms must have an unambiguous definition. One of the most comprehensive review of fossil wood genera is the article of Philippe & Bamford (2008), which is a reappraisal of most Mesozoic gymnosperm fossil genera. In the key, pit arrangement is particularly important to define the fossil-genera. Authors can use the indices proposed herein or give a detailed description of the radial pitting, giving quantified descriptions if possible. The indices proposed herein can be included in statistical analysis as continuous characters, which are usually more informative than binary or discrete characters as those proposed by IAWA committee (2004) for pit arrangement.
Measurements of typical extant araucarian pitting (Araucaria araucana (Molina) K. Koch and Araucaria angustifolia (Bertol.) Kuntze) give results of Cp always higher than 90% (Table 1). In woods of extant Podocarpaceae Cp ranges from 7.8% to 20.7%, in extant Cupressaceae from 0.9% to 4.0% and in extant Pinaceae from 5.3% to 12.5% (Table 1), the three families being usually considered to have abietinean radial pitting. Si exhibits no significant variation and has values close to 1 in most species, except for Araucaria angustifolia, which has frequently biseriate pit122 Pujana et al.: Quantifying tracheid pit arrangement 123 ting (Si=1.47). Table 1 shows the measurements made randomly by the authors and a variation between 0% and up to ca. 10% in Cp, may indicate some subjectivity (sometimes it is difficult to determine whether a line of pits is contiguous to another) in the measurements. These could also be caused by the different pit arrangement in the zones that were chosen to measure.
Two fossil woods measured for this study (Table 1), described with mixed radial pitting (Bonetti, 1966; Bodnar & Artabe, 2007; Pujana et al., 2014) gave a Cp of 73.0% (Protophyllocladoxylon francisae Pujana, Santillana & Marenssi) and 84.7% (Protojuniperoxylon ischigualastensis Bonetti). A Paleozoic wood, Cuyoxylon multipunctatus Pujana & Césari has a Cp of 100.0% (all horizontal lines of pits are contiguous), higher than extant Araucariaceae, and a Si=4.41, because of the bi- to 7-seriate radial pitting (Pujana & Césari, 2008). Agathoxylon antarcticus (Poole & Cantrill) Pujana, Santillana & Marenssi, a Cenozoic Araucariaceae (Pujana et al., 2015), has a Cp of 94.2%, similar to those of extant Araucariaceae. Podocarpoxylon sp. and Cupressinoxylon hallei Kräusel, usually considered genera with abietinean radial pitting (Philippe & Bamford, 2008), have a Cp of 32.9% and 1.9% respectively (Table 1). Cp is not directly proportional to whether a wood is mixed/araucarian/abietinoid, because Cp do not consider if the biseriate or multiseriate pits are opposite or alternate arranged. However, a relation is suggested because araucarioid woods have high values of Cp and abietinean low values according to our measurements (Table 1), while woods with mixed radial pitting have intermediate values (73.0% and 84.7%).
ACKNOWLEDGMENTS
We thank the anonymous reviewers for their thorough review and highly appreciate the corrections and suggestions, which significantly contributed to improving the quality of the article. This study was funded by the CONICET, Agencia Nacional de Promoción Científica y Técnológica (PICT 2012-0911) and National Natural Science Foundation of China (31470324).
BIBLIOGRAPHY
1. Bamford, M.K., M. Philippe & F. Thévenard. 2016. Long overdue extinction of the Protopinaceae. Review of Palaeobotany and Palynology 234: 25-30. [ Links ]
2. Beck, C.B. 2010. An Introduction to Plant Structure and Development. Cambridge University Press. Cambridge. 441 pp. [ Links ]
3. Bodnar, J. & A.E. Artabe. 2007. Estudio sistemático y paleodendrológico del leño de una Cupressaceae triásica de la Formación Ischigualasto, provincia de San Juan, Argentina. Ameghiniana 44: 303-319. [ Links ]
4. Bonetti, M.I.R. 1966. Protojuniperoxylon ischigualastensis sp. nov. del Triásico de Ischigualasto (San Juan). Ameghiniana 4: 211-216. [ Links ]
5. Carlquist, S. 1975. Ecological Strategies of Xylem Evolution. University of California Press, London. 259 pp. [ Links ]
6. Desplats, D. 1979. Sur une nouvelle espèce de Dadoxylon (Araucarioxylon) Kraus de l’Albien de Wissant, Boulonnais. 104e Congrès National des Sociétés Savantes: 185-195.
7. Eckhold, W. 1921. Die Hoftüpfel bei rezenten und fossilen Coniferen. Hochschulverlag, Breslau. 4 pp. [ Links ]
8. Eckhold, W. 1922. Die Hoftüpfel bei rezenten und fossilen Coniferen. Jahrbuch Königliche Preussische Geologische Landesansalt 42: 472-505. [ Links ]
9. Falcon-Lang, H.J. & D.J. Cantrill. 2000. Cretaceous (late Albian) coniferales of Alexander Island, Antarctica. 1: Wood taxonomy: a quantitative approach. Review of Palaeobotany and Palynology 111: 1-17. [ Links ]
10. Falcon-Lang, H.J. & D.J. Cantrill. 2001. Gymnosperm woods from the Cretaceous (mid-Aptian) Cerro Negro Formation, Byers Peninsula, Livingston Island, Antarctica: the arborescent vegetation of a volcanic arc. Cretaceous Research 22: 277-293. [ Links ]
11. García-Esteban, L. 2002. Anatomía e identificación de maderas de coníferas a nivel de especie. Fundación Conde del Valle de Salazar, Madrid, 421 pp. [ Links ]
12. Greguss, P. 1955. Identification of living Gymnosperms on the basis of xylotomy. Akadémiai Kiado, Budapest, 263 pp. [ Links ]
13. Greguss, P. 1972. Xylotomy of the living conifers. Akadémiai Kiadó, Budapest, 329 pp. [ Links ]
14. Henderson, E. & H.J. Falcon-Lang. 2011. Diversity and ontogeny of Pitus tree-trunks in the early Mississippian rocks of the Isle of Bute, Scotland: The importance of sample size and quantitative analysis for fossil wood systematics. Review of Palaeobotany and Palynology 166: 202-212. [ Links ]
15. IAWA Committee. 2004. IAWA list of microscopic features for softwood identification. IAWA Journal 25: 1-70. [ Links ]
16. Kenrick, P. & P.R. Crane. 1991. Water-conducting cells in early fossil land plants: implications for the early evolution in Tracheophytes. Botanical Gazette 152: 335-356. [ Links ]
17. Kräusel, R. 1917. Die Bedeutung der Anatomie lebender und fossiler Hölzer für die Phylogenie der Koniferen. Naturwissenschaften Wochenschrift 16: 305-311. [ Links ]
18. Marguerier, J. & P. Woltz. 1977. Anatomie comparée et systématique des Podocarpus malgaches. Adansonia 17: 155-192. [ Links ]
19. Müller-Stoll H (1951) Vergleichende Untersuchungen über die Abhängigkeit der Jahrringfolge von Holzart Standort und Klima. Bibliotheca Botanica 122: 1-93. [ Links ]
20. Nicol, W. 1834. Observations on the structure of recent and fossil Coniferae. Edinburgh New Philosophical Journal 16: 137-158. [ Links ]
21. Penhallow, D.P. 1907. A manual of the North American gymnosperms. Ginn & Company, Boston, 374 pp. [ Links ]
22. Philippe, M. & M.K. Bamford. 2008. A key to morphogenera used for Mesozoic conifer-like woods. Review of Palaeobotany and Palynology 148: 184- 207. [ Links ]
23. Philippe, M., A. Boura, C. Oh & D. Pons. 2014. Shimakuroxylon a new homoxylous Mesozoic wood genus from Asia, with palaeogeographical and palaeoecological implications. Review of Palaeobotany and Palynology 204: 18-26. [ Links ]
24. Poole, I. & D.J. Cantrill. 2001. Fossil woods from Williams Point beds, Livingston Island, Antarctica: a late Cretaceous southern high latitude flora. Palaeontology 44: 1081-1112. [ Links ]
25. Pujana, R.R. & S.N. Césari. 2008. Fossil woods in interglacial sediments from the Carboniferous Hoyada Verde Formation, San Juan province, Argentina. Palaeontology 51: 163-171. [ Links ]
26. Pujana, R.R., S.N. Santillana & S.A. Marenssi. 2014. Conifer fossil woods from the La Meseta Formation (Eocene of Western Antarctica): Evidence of Podocarpaceae-dominated forests. Review of Palaeobotany and Palynology 200: 122-137. [ Links ]
27. Pujana, R.R., S.A. Marenssi & S.N. Santillana. 2015. Fossil woods from the Cross Valley Formation (Paleocene of Western Antarctica): Araucariaceaedominated forests. Review of Palaeobotany and Palynology 222: 56-66. [ Links ]
28. Serra, C. 1966. Etude anatomique et paléogéographique de quelques espèces homoxylées du Sud Viêtnam et du Cambodge. Archives géologiques du Viêtnam 8: 59-131. [ Links ]
29. Torres, T., B. Cortinat & H. Meon. 1994. Microcachryxylon gothani nov. gen. n. sp., madera cretácica de la Isla James Ross, Antártica, in: VII Congreso Geológico Chileno. Concepción, Chile, pp. 1702-1706. [ Links ]
30. Vogellehner, D. 1967. Zur Anatomie und Phylogenie Mesozischer Gymnospermenhölzer 5: Prodromus zu einer Monographie der Protopinaceae. I. Die protopinoiden Hölzer der Trias. Palaeontographica Abteilung B 121: 30-51. [ Links ]
31. Vozenin-Serra, C. & J.A. Grant-Mackie. 1996. Les bois noriens des terrains Murihiku - Nouvelle Zélande - Intérêt paléophytogéographique. Palaeontographica Abteilung B 241: 99-125. [ Links ]
Recibido: 23-VIII-2016
Aceptado: 22-XI-2016

