










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkRevista del Museo Argentino de Ciencias Naturales
versión On-line ISSN 1853-0400
Rev. Mus. Argent. Cienc. Nat. vol.19 no.2 Ciudad Autónoma de Buenos Aires dic. 2017
PALEONTOLOGÍA
Nuevos registros paleobotánicos de la Formación Nestares (Jurásico Temprano), extremo austral de la Cuenca Neuquina, Argentina
Silvia C. Gnaedinger1 & Ana M. Zavattieri2
1 Área de Paleontología, Centro de Ecología Aplicada del Litoral, Consejo Nacional de Investigaciones Científicas y Técnicas (CECOAL-CCT CONICET Nordeste, UNNE), Facultad de Ciencias Exactas y Naturales y Agrimensura, Universidad Nacional del Nordeste (FaCENA-UNNE). Casilla de Correo 291, 3400 Corrientes, Argentina. scgnaed@hotmail.com.
2 Departamento de Paleontología, Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales (IANIGLA), Centro Científico Tecnológico, Consejo Nacional de Investigaciones Científica y Técnicas (IANIGLA-CCT CONICET Mendoza). Casilla de Correo 330, 5500 Mendoza, Argentina. amz@mendoza-conicet.gov.ar
Resumen
Se describen ejemplares de Rochipteris copiapensis (Solms-Laubach) Herbst, Troncoso & Gnaedinger y de Dejerseya lobata (Jones & de Jersey) Herbst los que corresponden a los primeros registros para la Formación Nestares (Jurásico Temprano), extremo austral de la Cuenca Neuquina, Argentina. El género Rochipteris estaba restringido hasta ahora al Triásico y la especie R. copiapensis (Solms-Laubach) Herbst, Troncoso & Gnaedinger al Noriano de Argentina y Chile por lo que este registro constituye la primera referencia del género en secuencias del Jurásico Inferior de Gondwana. Se propone una nueva combinación, Rochipteris truncata (McLoughlin, Tosolini & Drinnan) nov. comb. en reemplazo de Ginkgophytopsis truncata McLoughlin, Tosolini & Drinnan quedando en sinonimia Rochipteris truncata (Frengüelli) Morel, Artabe, Ganuza & Spalletti por prioridad, para lo cual se hace una revisión nomenclatural formal. El género Dejerseya tiene registro exclusivamente gondwánico y restringido al Triásico Superior-Jurásico Inferior, siendo este el segundo reporte del género en secuencias del Jurásico Inferior de Gondwana. Las especies asociadas de esta paleoflora son: Neocalamites carrerei (Zeiller) Halle, Cladophlebis oblonga Halle, C. mendozaensis (Geinitz) Frengüelli, Archangelskya protoloxsoma (Kurtz) Herbst, Otozamites albosaxatilis Herbst, O. ameghinoi Kurtz, O. bechei Brongniart, O. hislopii (Oldham) Feistmantel, Ptilophyllum acutifolium Morris, Alicurana artabei Herbst & Gnaedinger, A. nestarensis Herbst & Gnaedinger, Nilssonia taeniopteroides Halle y Elatocladus planus (Feistmantel) Seward. La edad de la Formación Nestares se asigna al Toarciano tardío sobre la base de su contenido palinológico previamente descripto.
Palabras clave: Paleobotánica; Formación Nestares; Jurásico Temprano; Cuenca Neuquina; Argentina.
Abstract
New paleobotanical records from the Nestares Formation (Early Jurassic), southern extreme of the Neuquén Basin, Argentina
Specimens of Rochipteris copiapensis (Solms-Laubach) Herbst, Troncoso & Gnaedinger and of Dejerseya lobata (Jones & de Jersey) Herbst are described for the first time from the Nestares Formation (Early Jurassic), southern extreme of the Neuquén Basin, Argentina. The genus Rochipteris was restricted up to now to Triassic, and the species R. copiapensis (Solms-Laubach) Herbst, Troncoso & Gnaedinger to Norian strata of Argentina and Chile, thus this record constitutes the first reference of the genus for Early Jurassic sequences of Gondwana. A new combination Rochipteris truncata (McLoughlin, Tosolini & Drinnan) nov. comb. is proposed to replace Ginkgophytopsis truncata McLoughlin, Tosolini & Drinnan leaving under synonymy Rochipteris truncata (Frengüelli) Morel, Artabe, Ganuza & Spalletti by priority for which a formal nomenclatural revision is given. The genus Dejerseya has exclusively gondwanic record restricted to Upper Triassic – Lower Jurassic being this, the second record of the genus in Early Jurassic strata of Gondwana. Associated species composing this paleoflora are: Neocalamites carrerei (Zeiller) Halle, Cladophlebis oblonga Halle, C. mendozaensis (Geinitz) Frengüelli, Archangelskya protoloxsoma (Kurtz) Herbst, Otozamites albosaxatilis Herbst, O. ameghinoi Kurtz, O. bechei Brongniart, O. hislopii (Oldham) Feistmantel, Ptilophyllum acutifolium Morris, Alicurana artabei Herbst & Gnaedinger, A. nestarensis Herbst & Gnaedinger, Nilssonia taeniopteroides Halle and Elatocladus planus (Feistmantel) Seward. The age of the Nestares Formation is assign to the late Toarcian on the basis of its palynological content previously described.
Key words: Paleobotany; Nestares Formation; Early Jurassic; Neuquén Basin; Argentina.
INTRODUCCIÓN
La Formación Nestares aflora en ambas márgenes del río Limay donde está enclavada la represa Alicurá, en el límite entre las provincias del Neuquén y Río Negro, en el borde sur-sureste de la Cuenca Neuquina (Legarreta & Gulisano, 1989; Gulisano, 1992; Gulisano & Gutiérrez Pleimling, 1994) constituyendo el único afloramiento y su localidad tipo. Esta unidad ha sido descripta en detalle por varios autores (Rosenfeld & Volkheimer, 1980; Arrondo et al., 1991; Spalletti et al., 1992; Spalletti, 1996; Zavattieri et al., 2008, entre otros). Las asociaciones microflorísticas a lo largo del perfil de Alicurá se componen de esporas, granos de polen de gimnospermas y fitoplancton (quistes de dinoflagelados, acritarcos, algas clorófitas, zignematáceas y prasinofíceas) reflejando condiciones de depositación continental fluvio-deltaico costero con desarrollo de subambientes locales (Zavattieri & Volkheimer, 2003; Zavattieri et al., 2008). Zavattieri et al. (2008) señalaron que los afloramientos de la Formación Nestares no serían más antiguos que Toarciano tardío sobre la base de su contenido palinológico (especies diagnósticas del microfitoplancton y de microfloras terrestres) y que esta unidad se superpone en relación de discordancia sobre los estratos de la Formación Paso Flores (Triásico Superior tardío) en el Valle del Río Limay. Sin embargo, Spalletti et al. (2007) y Morel et al. (2013) señalaron que la Formación Nestares se acota al Sinemuriano por considerar que su flora fósil es equivalente y sincrónica con las de las formaciones El Freno (Depocentro Atuel del norte de la Cuenca Neuquina, Mendoza) y Piedra del Águila (sur de la provincia del Neuquén). Esta última unidad fue referida por fechado radimétrico a dicha edad por Spalletti et al. (2010), aunque la relación geológico-estratigráfica con los afloramientos de la Formación Nestares no ha sido establecida, aflorando ambas unidades en áreas aisladas e independientes con secuencias sedimentarias y ambientes de depositación totalmente disímiles.
Registros paleobotánicos previos de la localidad tipo de la Formación Nestares fueron estudiados por Arrondo & Petriella (1980), Petriella & Arrondo (1982), Artabe (1982), Arrondo et al. (1984), Baldoni (1987) y Morel et al. (2013). La paleoflora de la Formación Nestares está compuesta por especies registradas en secuencias del Triásico Superior al Jurásico Inferior de Gondwana (Neocalamites carrerei (Zeiller) Halle, Cladophlebis mendozaensis (Geinitz) Frengüelli, Podozamites elongatus (Morris) Feistmantel, Elatocladus australis Frengüelli, Taeniopteris sp.); también contiene un grupo de especies cuyos registros se restringen al Jurásico Inferior (Equisetites frenguellii Orlando, Marattia münsteri (Goeppert) Zeiller, Gleichenites cf. sanmartini (Halle) Herbst, Cladophlebis oblonga Halle, Goeppertella diazii Arrondo & Petriella, Otozamites albosaxatilis Herbst, O. ameghinoi Kurtz, O. bechei Brongniart, O. hislopi (Oldham) Feistmantel, O. bengalensis Oldham & Morris, Ptilophyllum acutifolium Morris, Archangelskya protoloxsoma (Kurtz) Herbst, Scleropteris vincei Herbst, Nilssonia taeniopteroides Halle, Araucarites phillipsi Carruthers, Elatocladus planus (Feistmantel) Seward, Alicurana artabei Herbst & Gnaedinger y A. nestarensis Herbst & Gnaedinger y además por especies que se reportaron también en el Jurásico Medio como Elatocladus conferta (Oldham & Morris) Halle y Sagenopteris nilssoniana (Brongniart) Ward (Arrondo et al., 1991; Herbst & Gnaedinger, 2002; Zavattieri & Volkheimer, 2003; Quattrochio et al., 2007; Zavattieri et al., 2008).
En la presente contribución se describen por primera vez para la Formación Nestares hojas de Rochipteris copiapensis (Solms-Laubach) Herbst, Troncoso & Gnaedinger y fragmentos de pinnas asignadas a Dejerseya lobata (Jones & de Jersey) Herbst. Además, se da un listado de las plantas fósiles asociadas a dichas especies que ya fueron descriptas para esta unidad y que forman parte de los materiales depositados en el Museo Municipal “Carmen Funes”, Plaza Huincul, Provincia del Neuquén. Asimismo, se propone una nueva combinación, Rochipteris truncata (McLoughlin, Tosolini & Drinnan) nov. comb., en reemplazo de Ginkgophytopsis truncata McLoughlin, Tosolini & Drinnan, 2000 quedando consecuentemente en sinonimia Rochipteris truncata (Frengüelli), Morel, Artabe, Ganuza & Spalletti, 2011 por el principio de prioridad nomenclatural (ver Adenda, revisión nomenclatural).
MATERIALES Y MÉTODOS
Las plantas fósiles que se describen y mencionan en esta contribución proceden de los niveles basales aflorantes en la localidad tipo de la Formación Nestares, aguas abajo de la represa Alicurá (40° 34’ 57” S / 70° 44’ 53” O) (Fig. 1) y fueron coleccionadas cuando el dique estaba en construcción (1996). El perfil corresponde al publicado por Zavattieri et al. (2008) en el que se señalan los niveles con la megaflora fósil.

Fig. 1. Imagen del área de estudio de la Formación Nestares, localidad tipo, en los afloramientos del Dique Alicurá sobre el río Limay, límite entre las provincias del Neuquén y Río Negro. Los niveles fosilíferos se hallaron sobre la margen sur a pocos metros aguas abajo del paredón del dique. Imagen Landsat y Copernicus de Google Earth © 2016.
Las impresiones se estudiaron con microscopio estereoscópico Leica M50 y las fotografías fueron tomadas con cámara Leica EC3 con software Leica EZ. 3.2.1 y cámara Nikon Coolpix P100 del CECOAL-CONICET-UNNE-Corrientes. El material fósil estudiado se encuentra depositado en la colección paleontológica del Museo Municipal “Carmen Funes”, Plaza Huincul, Provincia del Neuquén, Argentina, bajo el prefijo MCF-PBPH (Museo Carmen Funes, Paleobotánica de Plaza Huincul).
DESCRIPCIÓN SISTEMÁTICA
Orden Petriellales Taylor, del Fueyo & Taylor, 1994
Familia Petriellaceae Taylor, del Fueyo & Taylor, 1994
Género Rochipteris (Herbst, Troncoso & Gnaedinger) Bomfleur, Decombeix, Schwendemann, Escapa, Taylor, Taylor & McLoughlin, 2014
Especie tipo. Rochipteris lacerata (Arber) Herbst, Troncoso & Gnaedinger, 2001
Rochipteris copiapensis (Solms-Laubach) Herbst, Troncoso & Gnaedinger, 2001
(Fig. 2A-G)

Fig. 2. (Pág. siguiente) A-E. Rochipteris copiapensis. A-C. dos fragmentos de la porción media-apical de la hoja; D. detalle de la venación de la porción apical de la hoja (flecha= anastomosis); E. porción basal de la hoja; F. detalle de la venación de la porción basal de la hoja (flecha= anastomosis); G. fragmento de la porción media-apical de la hoja. A(1), C-D= MCF-PBPH 186a; A(2), B= MCF-PBPH 186b; E-F= MCF-PBPH 157; G= MCF-PBPH 194. H-J. Dejerseya lobata. H-I. detalle de las pinnas con márgenes hendidos y/o partidos; J. detalle de la venación de los lóbulos de la pinna. H=MCF-PBPH 108; I=MCF-PBPH 112; J= MCF-PBPH 111. Escala gráfica: 1 cm, excepto D y F: 0,5 cm.
Descripción. Fragmentos de hojas de forma cuneada a triangular alargada angosta, sin base ni ápice preservado (Fig. 2A-C, G). Miden aproximadamente 10 cm de largo, el ancho máximo en la porción basal es de 0,5 cm (Fig. 2A-B) y en la porción apical del fragmento de 4,2 cm (Fig. 2A, C-D, G). Margen entero, un lado leve a marcadamente convexo, el otro cóncavo; ángulo de divergencia de los márgenes laterales desde la base es del orden de 12°-15°. Lámina dividida hasta dos o tres veces, formando segmentos de primer orden de 0,8-1,2 cm de ancho y de segundo orden de 0,6 cm (Fig. 2A-C). Venas bifurcadas y con escasas anastomosis (Fig. 2D, F). Densidad de venas: en la parte basal del fragmento es de 16 a 20 venas por cm; en la primera incisión 24 venas por cm y en los fragmentos terminales preservados hasta 12 venas por 0,5 cm.
Material estudiado. MCF-PBPH 011a, 143, 157, 178, 186a-b, 194.
Comentarios. El género Rochipteris incluye hojas gondwánicas previamente asignadas a Chiropteris Kurr ex Bronn (hojas sin anastomosis) o a Ginkgophytopsis Høeg (hojas pecioladas con venas dicotomizadas, género descripto para el Paleozoico del Hemisferio Norte) (ver Herbst et al., 2001 y Barone-Nugent et al., 2003). Retallack (1980) enmendó el género Ginkgophytopsis agregando el carácter de venación anastomosada y consecuentemente, cambiando por completo la diagnosis original. Anderson & Anderson (2003) crearon el género Kannaskoppifolia para este tipo de hojas del Triásico de África estableciendo como caracteres genéricos, la conexión a ejes y a estructuras reproductivas. Estos autores y, posteriormente Holmes & Anderson (2005), propusieron utilizar el nombre Rochipteris para hojas aisladas y Kannaskoppifolia para las mismas hojas conectadas a un eje. Bomfleur et al. (2014) proponen el género Kannaskoppifolia como sinónimo de Rochipteris, por prioridad nomenclatural, aunque en la descripción de la enmienda genérica no señalan el carácter de hojas conectadas a ejes. Sin embargo, luego en las observaciones incluyen el carácter de las hojas conectadas a ejes para el género Rochipteris. A pesar de que la enmienda genérica realizada por estos autores es idéntica a la propuesta por Herbst et al. (2001), se considera válido incluir a Kannaskoppifolia en sinonimia de Rochipteris y agregar la conexión de las hojas a ejes como carácter diagnóstico de este último (Bomfleur et al., 2014).
Los especímenes descriptos se identifican como Rochipteris copiapensis (Solms-Laubach) Herbst, Troncoso & Gnaedinger por los siguientes caracteres diagnósticos: ”hoja obovada a elíptica, cuneada a triangular alargada angosta con borde laterales entero, un lado leve a marcadamente convexo, el otro cóncavo, ápice inciso formando dos o tres segmentos, y venación dicotomizada con anastomosis escasas, principalmente a partir del segundo tercio basal de la lámina, aumentando su frecuencia hacia el ápice, formando una red de aréolas romboidales, muy angostas y largas” (Herbst et al., 2001, p. 266).
Actualmente, Rochipteris es un género estrictamente gondwánico que comprende 14 especies, cuyo registro se extiende desde el Triásico Medio al Triásico Superior. En el Cuadro 1 se muestra la distribución temporal y geográfica de las especies de Rochipteris. Para el Triásico se registraron cuatro especies en Argentina (Herbst et al., 2001; Morel et al., 2011), dos en Chile (Herbst et al., 2001; Troncoso et al., 2004), ocho en Australia (Barone-Nugent et al., 2003; Holmes & Anderson, 2005), dos en Nueva Zelandia (Retallack, 1980), una especie en Antártida (Bomfleur et al., 2014) y una en Sudáfrica (Kannaskoppifolia vincularis Anderson & Anderson, 2003). Existen diversos especímenes que fueron descriptos e ilustrados como Rochipteris sp. en Gondwana: Argentina (Artabe et al., 2007; Gnaedinger & Zavattieri, 2015), Australia (Barone-Nugent et al., 2003); Chile (Herbst et al., 2001; Troncoso et al., 2004; Herbst & Troncoso, 2012) y África (Anderson & Anderson, 2003; como Kannaskoppifolia sp.). Por otra parte, para el Cretácico Inferior de Australia, McLoughlin et al. (2000) identificaron este tipo de hojas como Ginkgophytopsis truncata McLoughlin, Tosolini & Drinnan, basándose en la enmienda de este género realizada por Retallack (1980) (Cuadro 1). Por todo lo mencionado anteriormente, se considera que los ejemplares descriptos por McLoughlin et al. (2000) como G. truncata requieren una combinación al género Rochipteris (ver adenda).
Cuadro 1. Distribución temporal y geográfica de especies de Rochipteris (Herbst et al., 2001; Barone-Nugent et al., 2003; Anderson & Anderson, 2003; Holmes & Anderson, 2005; Morel et al., 2011; Bomfleur et al., 2014; McLoughlin et al., 2000). *= Kannaskoppifolia. TM-S.= Triásico Medio y Superior; TS.= Triásico Superior; JI.= Jurásico Inferior; KI.= Cretácico Inferior. Ar.=Argentina; Ch.=Chile; Au.=Australia; NZ.=Nueva Zelandia; An.=Antártida y Af.=África. Gris claro= especies del Noriano; gris oscuro= especie descripta en esta contribución).

R. copiapensis (Solms-Laubach) Herbst, Troncoso & Gnaedinger fue registrada previamente en el Triásico Superior (Noriano) en la Formación La Ternera, Chile y en la Formación Laguna Colorada (Grupo El Tranquilo, Macizo del Deseado), Argentina (Herbst et al., 2001). En esta contribución se la identifica por primera vez con certeza para el Jurásico Inferior de Gondwana (Cuadro 1), aunque Charrier et al. (2007) indicaron que la parte superior de la Formación La Ternera de Chile puede llegar al Hettangiano?, que es de donde provienen los ejemplares de este taxón.
Adenda. Revisión nomenclatural.
Rochipteris truncata (McLoughlin, Tosolini & Drinnan) nov. comb.
Basónimo
2000. Ginkgophytopsis truncata McLoughlin, Tosolini & Drinnan, p. 497, fig. 8D
Sinónimos
1918. Ginkgo sp. Walkom, p. 9; pl. 2, fig. 6.
1946. Ginkgoites truncata Frengüelli, p. 121, lám. 1, figs. 1B, 2; lám. 4, fig. 4; lám. 6, fig. 2; lám. 7, figs. 1-2.
2011. Rochipteris truncata (Frengüelli) Morel, Artabe, Ganuza & Zúñiga, p. 524-526, fig. 2.1.
Diagnosis. De acuerdo con la diagnosis original de la especie dada por McLoughlin et al. (2000, p. 497).
Las especies Rochipteris chilensis Gnaedinger, R. obtriangulata Holmes & Anderson, 2005, Rochipteris truncata (Frengüelli) Morel, Artabe, Ganuza & Zúñiga y Ginkgophytopsis truncata McLoughlin, Tosolini & Drinnan son similares en la forma de la lámina foliar, entera, cuneada-triangular y márgenes rectos (McLoughlin et al., 2000; Herbst et al., 2001; Holmes & Anderson, 2005; Morel et al., 2011). La primera especie se distingue de las otras por tener el ápice curvado y bordes ondulados, mientras que las restantes poseen ápice truncado y bordes rectos. R. obtriangulata Holmes & Anderson, 2005 se distingue por la distribución de las anastomosis y la densidad de la venación. Las especies Rochipteris truncata (Frengüelli) Morel, Artabe, Ganuza & Zúñiga y Ginkgophytopsis truncata McLoughlin, Tosolini & Drinnan son esencialmente coincidentes en su morfología foliar (Cuadro 2).
Cuadro 2. Comparación morfológica entre especies de Rochipteris que comparten los caracteres de lámina foliar entera, cuneada-triangular y margen apical recto.
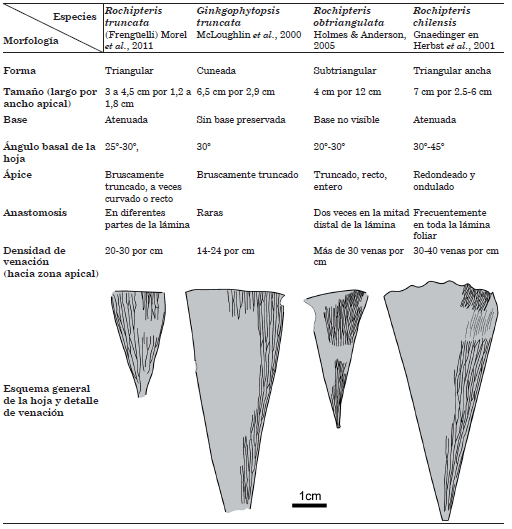
Ginkgophytopsis truncata McLoughlin, Tosolini & Drinnan fue creada por McLoughlin et al. (2000) siguiendo la enmienda del género realizada por Retallack (1980) e incluyeron como sinonimia al taxón identificado como Ginkgo sp. por Walkom (1918), ambos del Cretácico Inferior de Queensland, Australia. Por su parte, Rochipteris truncata (Frengüelli) Morel, Artabe, Ganuza & Zúñiga fue combinado a Rochipteris por Morel et al. (2011) sobre la base de ejemplares descriptos por Frengüelli (1946) como Ginkgoites truncata del Triásico de Potrerillos (Cuenca Cuyana), Mendoza, Argentina. Al crear la especie Ginkgoites truncata Frengüelli, 1946, el autor señaló la similitud con las hojas australianas descriptas por Walkom (1918) como Ginkgo sp.
Por lo previamente analizado, por la comparación realizada en el Cuadro 2 y de acuerdo con el principio de prioridad nomenclatural establecida por el Código Internacional de Nomenclatura para algas, hongos y plantas (McNeill et al., 2012) se establece formalmente la combinación de la especie Ginkgophytopsis truncata McLoughlin, Tosolini & Drinnan al género Rochipteris y los ejemplares descriptos por Frengüelli (1946) y Morel et al. (2011) resultan consecuentemente, sinónimos.
Orden Peltaspermales Meyen, 1987
Familia Matatiellaceae Anderson & Anderson, 2003
Género Dejerseya Herbst, 1977
Especie tipo. Dejerseya lobata (Jones & de Jersey) Herbst, 1977.
Dejerseya lobata (Jones & de Jersey) Herbst, 1977
(Fig. 2H-J)
Descripción. Fragmentos de pinnas lineares de 3,5 cm de largo por 1,4 cm de ancho en la parte basal, con márgenes distales hendidos y/o partidos. Vena media longitudinalmente estriada que mide en la base 0,1 cm. Los lóbulos de las pinnas miden 0,7 cm de ancho basal por 1 cm de largo aproximado; ápice obtuso a redondeado (Fig. 2H-I). A partir de la vena media de la pinna y en cada lóbulo, emerge una vena secundaria principal que se dicotomiza hasta 6-10 veces formando grupos de venas. Entre estos grupos se observan una o dos venas simples o bifurcadas que nacen de la vena media de la pinna (Fig. 2J).
Material estudiado. MCF-PBPH 017b, 104b, 105 (contraimpresión 109), 108 (contraimpresión 120), 110b, 111, 112b.
Comentarios. Herbst (1977) crea el género Dejerseya sobre la base de ejemplares descriptos como Rienitsia Walkom por Jones & de Jersey (1947) y posteriormente también por Douglas (1969). Las especies referidas para el nuevo género son, además de la especie tipo Dejerseya lobata (Jones & de Jersey) Herbst, D. incisa (Jones & de Jersey) Herbst y D. variabilis (Douglas) Herbst (para más detalle ver Herbst, 1977). Jones & de Jersey (1947) mencionan el carácter de “frondes pinnadas” para las especies Rienitsia lobata y R. incisa. Gnaedinger & Herbst (1998) confirmaron el carácter de hojas pinnadas en ejemplares del Triásico de Patagonia (Argentina) para Dejerseya lobata. Anderson & Anderson (1989) aceptaron el género Dejerseya utilizando como especie tipo a D. lobata, pero incluyeron todas las especies mencionadas en D. lunensis (Johnston), anulando la condición de especie tipo y redefinieron el género para incluir hojas enteras, hendidas, lobuladas hasta pinnadas. Gnaedinger & Herbst (2004) mantienen como especie tipo a Dejerseya lobata (Jones & de Jersey) Herbst ampliando la diagnosis genérica. Estos autores además, establecen claramente la diferencia entre D. lunensis (Johnston) Anderson & Anderson (hojas enteras con márgenes enteros, ondulados, lolulados, superficialmente pinnada (hojas hendidas) y profundamente pinnada (hojas pinnatipartidas) y la especie tipo D. lobata (Jones & de Jersey) Herbst (hojas monopinnadas a bipinnadas con pinnas con márgenes enteros, ondulados, hendidos a partidos), sobre la base de especímenes del Triásico de Chile (ver más detalles en Gnaedinger & Herbst, 2004). Por último, Bomfleur et al. (2011) al analizar ejemplares del Triásico de Antártida, incluyen –en parte- los registros de las especies D. lobata y D. lunensis en D. lobata, realizando una enmienda a la diagnosis genérica y específica, eliminando la condición de fronde pinnada, bipinnatifida a bipinnada, que es característico de D. lobata (ver más detalles en Gnaedinger & Herbst, 2004 y Bomfleur et al., 2011). En esta contribución se considera que Dejerseya Herbst comprende dos especies, Dejerseya lobata (Jones & de Jersey) Herbst y D. lunensis (Johnston) Anderson & Anderson de acuerdo con el criterio de Gnaedinger & Herbst (2004).
Los fragmentos de pinnas aquí se asignan a Dejerseya lobata (Jones & de Jersey) Herbst por su coincidencia con los caracteres diagnósticos dados por Herbst (1977) y por Gnaedinger & Herbst (1998, 2004). Los principales rasgos morfológicos son: hojas con pinnas lineares con márgenes distales hendidos y/o partidos con grupos de venas en cada lóbulo que se forman a partir de una vena secundaria principal que se dicotomiza hasta 6-10 veces; entre estos grupos de venas se observan una o dos venas simples o bifurcadas que nacen de la vena media de la pinna.
Dejerseya lobata (Jones & de Jersey) Herbst tiene registro gondwánico y restringido al Triásico Superior-Jurásico Inferior (Gnaedinger & Herbst, 1998; Anderson & Anderson, 2003; Spalletti et al., 2007). En el Jurásico Inferior esta especie fue previamente identificada en la Formación El Freno, Argentina (Spalletti et al., 2007), por lo que representa la segunda referencia del género en secuencias del Jurásico Inferior de Gondwana.
Otros taxones asociados
Se presenta a continuación una lista de taxones identificados resultado del estudio de la nueva colección paleobotánica procedente de la localidad tipo de la Formación Nestares y que fueron previamente descriptos para esta unidad (ver introducción). Luego del nombre específico y entre paréntesis, se incluye el número de colección del material estudiado y la referencia bibliográfica pertinente.
Orden EQUISETALES Dumortier
Neocalamites carrerei (Zeiller) Halle
(MCF-PBPH 167, 181, 191. Arrondo & Petriella, 1980)
Orden OSMUNDALES Bromhead
Cladophlebis oblonga Halle
(MCF-PBPH 022, 012, 187, 195. Arrondo & Petriella, 1980)
Cladophlebis mendozaensis (Geinitz) Frengüelli
(MCF-PBPH 123a. Arrondo & Petriella, 1980)
Orden PTERIDOSPERMALES Incertae sedis
Archangelskya protoloxsoma (Kurtz) Herbst
(MCF-PBPH 178a. Morel et al., 2013)
Orden BENNETTITALES
Otozamites albosaxatilis Herbst
(MCF-PBPH 126b, 130c. Arrondo & Petriella, 109 Gnaedinger & Zavattieri: Nuevos registros paleobotánicos de la Formación Nestares
1980; Morel et al., 2013)
Otozamites ameghinoi Kurtz
(MCF-PBPH 131, 135, 139, 147a, 156a (contraimpresión 163a). Arrondo & Petriella, 1980, Morel et al., 2013)
Otozamites bechei Brongniart
(MCF-PBPH 130a. Arrondo & Petriella, 1980, Morel et al., 2013)
Otozamites hislopii (Oldham) Feistmantel
(MCF-PBPH 021 (contraimpresión 106), 104a (contraimpresión107), 105b, 112a (contraimpresión 017), 113a, 120, 125a-b, 126a, 130 a-b 133a, 138, 145, 150a-b. Arrondo & Petriella, 1980; Morel et al., 2013)
Ptilophyllum acutifolium Morris
(MCF-PBPH 109a, 156b (contraimpresión 163b). Arrondo & Petriella, 1980; Morel et al., 2013).
Orden CYCADALES Dumortier
Alicurana artabei Herbst & Gnaedinger (=Kurtziana brandmayri Frengüelli)
(MCF-PBPH 125 a, c, 141 (contraimpresión 154), 146, 148, 151. Arrondo & Petriella, 1980, Herbst & Gnaedinger, 2002; Morel et al., 2013)
A. nestarensis Herbst & Gnaedinger (=Kurtziana cacheutensis (Kurtz) Frengüelli).
(MCF-PBPH 011, 125c, 151, 188. Arrondo & Petriella, 1980; Herbst & Gnaedinger, 2002; Morel et al., 2013)
Nilssonia taeniopteroides Halle
(MCF-PBPH 140,142. Arrondo & Petriella, 1980, Morel et al., 2013)
Orden CONIFERALES
Elatocladus planus (Feistmantel) Seward
(MCF-PBPH 133b. Arrondo & Petriella, 1980; Morel et al., 2013)
CONCLUSIONES
En esta contribución se describen por primera vez para la Formación Nestares, ejemplares de Rochipteris copiapensis (Solms-Laubach) Herbst et al. y fragmentos de Dejerseya lobata (Jones & de Jersey) Herbst que se suman a la composición de la paleoflora conocida hasta el momento para la localidad tipo de esta unidad sedimentaria.
Sobre la base de la nueva combinación propuesta de Rochipteris truncata, en esta contribución se extiende el registro temporal de Rochipteris desde el Triásico Medio-Superior hasta el Cretácico Inferior; para R. copiapensis (Solms-Laubach) Herbst et al. este registro constituye la primera referencia del género en secuencias del Jurásico Inferior de Gondwana. Por su parte, hojas de Dejerseya ya fueron descriptas para el Jurásico Inferior (Formación El Freno, norte de la Cuenca Neuquina, Mendoza, Argentina), por lo que las impresiones identificadas como Dejerseya lobata en Neuquén representan el segundo hallazgo para Gondwana. Este confirma la hipótesis dada por Bomfleur et al. (2011) al asignar a Dejerseya lobata (Jones & de Jersey) Herbst a las Matatiellaceae, un grupo de peltaspermales endémico del Hemisferio Sur, y la persistencia de hojas similares en el Jurásico y Cretácico Inferior indicaría que este grupo subsistió durante gran parte del Mesozoico.
Se identificaron conjuntamente con las especies aquí descriptas 13 taxones que fueron previamente registrados en el perfil tipo de esta unidad: Neocalamites carrerei (Zeiller) Halle, Cladophlebis oblonga Halle, C. mendozaensis (Geinitz) Frengüelli, Archangelskya protoloxsoma (Kurtz) Herbst, Otozamites albosaxatilis Herbst, O. ameghinoi Kurtz, O. bechei Brongniart, O. hislopii (Oldham) Feistmantel, Ptilophyllum acutifolium Morris, Alicurana artabei Herbst & Gnaedinger (=Kurtziana brandmayri Frengüelli), A. nestarensis Herbst & Gnaedinger (=Kurtziana cacheutensis (Kurtz) Frengüelli), Nilssonia taeniopteroides Halle & Elatocladus planus (Feistmantel) Seward.
Como se mencionó en la introducción, la flora fósil de la Formación Nestares comparte varios taxones con las paleofloras de las formaciones El Freno y Piedra del Águila, ambas consideradas como Eojurásicas. Todas ellas se componen de especies cuyos biocrones se restringen al Triásico Superior-Jurásico Inferior, algunas son exclusivas para el Jurásico Temprano de Gondwana, mientras que otros taxones tienen un biocrón más amplio y cosmopolita dentro de Gondwana. La información provista por Zavattieri et al. (2008) y por Spalletti et al. (2010) se oponen a las consideraciones dadas por Morel et al. (2013), quienes concluyeron que la edad de la Formación Nestares es Sinemuriana por su contenido megaflorístico que comparte especies con las unidades mencionadas (Artabe et al., 2005; Quattrocchio et al., 2007; Spalletti et al., 2007; Morel et al., 2013) y por las relaciones estratigráficas con la Formación Piedra del Águila (datada radimétricamente al Sinemuriano). Morel et al. (2013) sostienen que la localidad tipo de Formación Nestares aflora en el sector noroccidental del Macizo Nordpatagónico, mientras que la Formación Piedra del Águila corresponde al sur de la Cuenca Neuquina (Spalletti et al., 2010), por lo cual, ambas unidades no tienen relaciones geológicas o estratigráficas que las vinculen. Sin embargo, el contenido palinológico de la Formación Nestares constituye hasta ahora, la referencia cronológica y paleoambiental más confiable indicando que esta secuencia sería tan joven como Toarciano tardío, lo cual diferenciaría en forma neta la edad de esta unidad con respecto a las otras unidades mencionadas (Zavattieri et al., 2008) y que ya fue analizado por Spalletti et al. (2010) para la Formación Piedra del Águila.
AGRADECIMIENTOS
La asociación de plantas fósiles que forman parte de esta contribución fueron coleccionadas en 1996 por una de las autoras (AMZ), participante de un proyecto liderado por el Dr. Rodolfo Coria y financiado por “The Dinosaur Society” (1996-1997). Esta contribución fue parcialmente financiada por la Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT-FONCyT) Argentina (PICT 2011-2546, AMZ) y por la Secretaría General de Ciencia y Técnica, SGCYT-UNNE (PI 2015-2018, Q005-2014) y CONICET (PIP 2014-2016. 112 201301 00317) a SCG. Agradecemos al Dr Rodolfo A. Coria, director del Museo “Cármen Funes”, Plaza Huincul, Provincia del Neuquén por facilitar el acceso para estudiar el material depositado en dicha institución. También extendemos nuestra gratitud al Lic. Flavio Bellardini, Curador encargado de la Colección Paleontológica del mencionado Museo por su constante colaboración y asistencia con los materiales y con el repositorio. También hacemos extensivo nuestro agradecimiento al Prof. Dr Steve McLoughlin (Swedish Museum of Natural History) Estocolmo por facilitarnos la foto original del ejemplar de Ginkgophytopsis truncata del Cretácico Inferior de Maryborough Basin para comparar con el material argentino. Las autoras agradecen a los revisores Dres I. Escapa y E. Morel por la lectura crítica del manuscrito y las sugerencias realizadas al mismo.
BIBLIOGRAFÍA
1. Anderson J.M. & H.M. Anderson. 1989. Palaeoflora of southern Africa: Molteno Formation (Triassic), v. 2: Gymnosperms (excluding Dicroidium). Balkema, Rotterdam. 567 pp. [ Links ]
2. Anderson, J.M. & H.M. Anderson. 2003. Heyday of the gymnosperms: systematics and biodiversity of the Late Triassic Molteno fructifications. Strelitzia 15. National Botanical Institute, Pretoria, 398 pp. [ Links ]
3. Arrondo, O.G. & B. Petriella. 1980. Alicurá, una nueva localidad plantífera liásica de la provincia de Neuquén, Argentina. Ameghiniana 17: 200–215.
4. Arrondo, O.G., A.E. Artabe, E.M. Morel & B. Petriella. 1984. Floras Fósiles Mesozoicas. En: V.A. Ramos. (Ed.), Geología y Recursos Naturales de la Provincia de Río Negro. 9º Congreso Geológico Argentino (San Carlos de Bariloche), Relatorio, pp. 367–372.
5. Arrondo, O., L. Spalletti, E.M. Morel & D. Ganuza. 1991. The sedimentological and paleobotanical characteristics of an Upper Triassic–Lower Liassic basin in northwestern Patagonia (Argentina). En: H. Ulbrich & A.C. Rocha-Campos (eds.), 7th Gondwana Symposium Proceedings, Instituto de Geociencias, Universidade de Sao Paulo (Sao Paulo), 714: 517– 532.
6. Artabe, A.E. 1982. Presencia de Podozamites elongatus (Morris) Feistmantel (Coniferales, Podozamitaceae) en la tafoflora de Alicurá (Provincia de Neuquén, Argentina). Ameghiniana 19: 361–365.
7. Artabe, A.E., D.G. Ganuza, L.A. Spalletti, A. Zúñiga & E.M. Morel. 2005. Revisión de la paleoflora del Cerro La Brea (Jurásico Temprano), provincia de Mendoza, Argentina. Ameghiniana 42: 429–442.
8. Artabe, A.E., E.M. Morel, D.G. Ganuza, A.M. Zavattieri & L.A. Spalletti. 2007. La paleoflora triásica de Potrerillos, Provincia de Mendoza, Argentina. Ameghiniana 44: 279–301.
9. Baldoni, A. 1987. Megafloras del Jurásico inferior de la Patagonia extraandina. Con algunas consideraciones sobre Venezuela y Colombia. En: W. Volkheimer (ed.): Bioestratigrafía de los Sistemas regionales del Jurásico y Cretácico de América del Sur. 1. Comité Sudamericano Jurásico y Cretácico, 159‑199. [ Links ]
10. Barone-Nugent, E.D., S. McLoughlin & A.N. Drinnan. 2003. New species of Rochipteris from the Upper Triassic of Australia. Review of Palaeobotany and Palynology 123: 273–287.
11. Bomfleur, B., E.L. Taylor, T.N. Taylor, R. Serbet, M. Krings & H. Kerp. 2011. Systematics and paleoecology of a new peltaspermalean seed fern from the Triassic polar vegetation of Gondwana. International Journal of Plant Sciences 172: 807–835.
12. Bomfleur, B., A.L. Decombeix, A.B. Schwendemann, I.H. Escapa, E.L. Taylor, T.N. Taylor & S. McLoughlin. 2014. Habit and ecology of the Petriellales, an unusual group of seed plants from the Triassic of Gondwana. International Journal of Plant Sciences, 175: 1062–1075.
13. Charrier, R., L. Pinto & M.P. Rodríguez. 2007. Tectonostratigraphic evolution of the Andean Orogeny in Chile. En: T. Moreno & W. Gibbons (eds), The Geology of Chile. The Geological Society 21–114. London.
14. Douglas J. G. 1969. The Mesozoic floras of Victoria. Parts 1 and 2. Memoirs of the Gelogical Survey of Victoria 28. 310 p. [ Links ]
15. Frengüelli, J. 1946. Contribuciones al conocimiento de la flora del Gondwana superior en la Argentina. 33 Ginkgoales de los estratos de Potrerillos en la Precordillera de Mendoza. Notas del Museo de La Plata, Paleontología 87: 299–325.
16. Gnaedinger, S. & R. Herbst. 1998. La Flora Triásica del Grupo El Tranquilo, Provincia de Santa Cruz (Patagonia). Parte V. Pteridophylla. Ameghiniana 35: 53–65.
17. Gnaedinger, S. & R. Herbst. 2004. Pteridophylla triásicas del Norte de Chile. Géneros Dejerseya Herbst, Linguifollium (Arber) Retallack y Yabeiella Ôishi. Revista del Museo Argentino de Ciencias Naturales, nueva serie 6: 49–59.
18. Gnaedinger, S. & A.M. Zavattieri. 2015. Paleoflora de la Formación Llantenes (Triásico Superior), provincia de Mendoza, Argentina. Nuevos registros. XVI Simposio Argentino de Paleobotánica y Palinología. La Plata. Ameghiniana 52, Suplemento Resúmenes, p. 34. [ Links ]
19. Gulisano, C.A. 1992. Paleogeographic evolution of west-central Argentina. En: G.E.G. Westermann (ed.), The Jurassic of Circum-Pacific, 150-151. Cambridge University Press. [ Links ]
20. Gulisano, C.A. & A.R. Gutiérrez Pleimling. 1994. The Jurassic of the Neuquén Basin. Field Guide, A – Neuquén Province. IV Jurassic stratigraphy of the Neuquén Basin. Asociación Geológica Argentina, Serie E 2: 21–35.
21. Herbst, R. 1977. Sobre Marattiales (Filicopsidae) triásicas de Argentina y Australia. Parte II. Los géneros Danaeopsis y Rienitsia. Ameghiniana 14: 19–32.
22. Herbst, R. & A. Troncoso. 2012. La flora Triásica de la Quebrada Doña Inés Chica, Región de Atacama, Chile. GAEA Journal of Geosciences 8: 55–66.
23. Herbst, R. & S. Gnaedinger. 2002. Kurtziana (Pteridospermae? incertae sedis) y Alicurana nov. gen. (Cycadopsida) del Triásico y Jurásico temprano de Argentina. Ameghiniana 39: 331–341.
24. Herbst, R., A. Troncoso & S. Gnaedinger. 2001. Rochipteris nov. gen., hojas incertae sedis (=Chiropteris pro parte) del Triásico Superior de Argentina y Chile. Ameghiniana 38: 257–269.
25. Holmes, W.B.K. & H.M. Anderson. 2005. The Middle Triassic megafossil flora of the Basin Creek Formation, Nymboida Coal Measures, New South Wales, Australia. 5. The genera Lepidopteris, Kurtziana, Rochipteris and Walkomiopteris. Proceedings of the Linnean Society of New South Wales 126: 39. [ Links ]
26. Jones, D.G. & N.J de Jersey. 1947. The flora of the Ipswich Coal Measures morphology and floral succession. Department of Geology, University of Queensland 3: 1-88. [ Links ]
27. Legarreta, L. & C.A. Gulisano. 1989. Análisis estratigráfico secuencial de la Cuenca Neuquina (Triásico Superior - Terciario Inferior). En: Chebli, G.A. & Spalletti, L.A. (eds)., Cuencas sedimentarias Argentinas. Serie Correlación Geológica 6: 221–243.
28. McLoughlin, S., A.P. Tosolini & A.N. Drinnan. 2000. Revision of an Early Cretaceous macroflora from the Maryborough Formation, Maryborough Basin, Queensland, Australia. Memoirs of the Queensland Museum 45: 483–503.
29. McNeill, J., F.R. Barrie, W.R. Buck, V. Demoulin, W. Greuter, D.L. Hawksworth, P.S. Herendeen, S. Knapp, K. Marhold, J. Prado, W.F. Prud’homme van Reine, G.F. Smith, J.H. Wiersema, N.J. & Turland (eds.). 2012. International Code of Nomenclature for algae, fungi, and plants (Melbourne Code): adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011. Koeltz Scientific Books, Königstein, 208 p.
30. Meyen, S.V. 1987. Fundamentals of Palaeobotany. Chapman and Hall, London. 432 p. [ Links ]
31. Morel, E.M., A.E. Artabe, D.G. Ganuza & A. Zúñiga. 2011. La paleoflora Triásica del Cerro Cacheuta, Provincia de Mendoza, Argentina. Petriellales, Cycadales, Ginkgoales, Voltziales, Coniferales, Gnetales y Gimnospermas Incertae Sedis. Ameghiniana 48: 520–540.
32. Morel, E. M., D.G. Ganuza, A.E. Artabe & L.A. Spalletti. 2013. Revisión de la paleoflora de la Formación Nestares (Jurásico Temprano), Provincia del Neuquén y Río Negro, Argentina. Ameghiniana 50: 493–508.
33. Petriella, B. & O.G. Arrondo. 1982. El género Kurtziana Frengüelli: su morfología y vinculaciones. Ameghiniana 19: 209–215.
34. Quattrocchio, M.E., M.A. Martínez & W. Volkheimer. 2007. Las floras jurásicas de la Argentina. Asociación Paleontológica Argentina. Publicación Especial 11 Ameghiniana 50º aniversario: 87–100.
35. Retallack, G.J. 1980. Middle Triassic megafossil plants and trace fossils from Tank Gully, Canterbury, New Zealand. Journal of the Royal Society of New Zealand 10: 31–63.
36. Rosenfeld, U. & W. Volkheimer. 1980. Turbidite und andere Rhythmite im tieferen Jura des Neuquén-Beckens (Argentinien). Turbidites and other rhythmic sediments in the Lower Jurassic of the Neuquén Basin (Argentine). Neues Jahrbuch für Geologie und Palaöntologie Abhandlungen (Stuttgart) 159 (3): 379–421.
37. Spalletti, L.A. 1996. Lower Jurassic anastomosing fluvial deposits from the cratonic margin of an Andean Back – Arc basin, Argentina. En: A.C. Riccardi (Ed.), Advances in Jurassic Research. Transtec Publications, Switzerland 1–2: 265–274.
38. Spalletti, L.A., J.C. Merodio & S.D. Matheos. 1992. Composición, procedencia y marco tectónico de sedimentitas eojurásicas (Formación Nestares) del extremo sudeste de la Cuenca Neuquina. Revista de la Asociación Geológica Argentina 47: 277–286.
39. Spalletti, L.A., E.M. Morel, J.R. Franzese, A.E. Artabe, D.G. Ganuza, & A. Zúñiga. 2007. Contribución al conocimiento sedimentológico y paleobotánico de la Formación El Freno (Jurásico Temprano) en el valle superior del río Atuel, Mendoza, Argentina. Ameghiniana 44: 367–386.
40. Spalletti, L.A., J.R. Franzese, E.M. Morel, L. D´elia, A. Zúñiga & C. Fanning. 2010. Consideraciones acerca de la sedimentología, paleobotánica y geocronología de la Formación Piedra del Águila (Jurásico Inferior, Neuquén, República Argentina). Revista de la Asociación Geológica Argentina 66: 305–313.
41. Taylor, T.N., G.M. del Fueyo & E.L. Taylor. 1994. Permineralized seed fern cupules from the Triassic of Antarctica: implications for cupule and carpel evolution. American Journal of Botany 81: 666–677.
42. Troncoso, A., R. Herbst & J. Muñoz Bravo. 2004. La flora fósil de la Formación Huimpil-Llafquentué, Triásico Superior, IX Región, Chile. Revista del Museo Argentino de Ciencias Naturales, nueva serie 6: 313–319.
43. Walkom, A.B. 1918. Mesozoic floras of Queensland. Part 2. The flora of the Maryborough (marine) series. Geological Survey of Queensland Publication 262: 1–21.
44. Zavattieri, A.M. & W. Volkheimer. 2003. Palynostratigraphy and paleoenvironments of Early Jurassic strata (Nestares Formation) in northern Patagonia, Argentina. Part 1. Terrestrial species. Ameghiniana 40: 545–558.
45. Zavattieri, A.M., U. Rosenfeld & W. Volkheimer. 2008. Palynofacies analysis and sedimentary environment of Early Jurassic coastal sediments at the southern border of the Neuquén Basin, Argentina. Journal of South American Earth Sciences 25: 227–245.
Recibido: 28-VI-2017
Aceptado: 25-IX-2017

