











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkINTRODUCCIÓN
Las garzas (Pelecaniformes, Ardeidae) se ali mentan principalmente de presas como peces, pe queños mamíferos, crustáceos, insectos y reptiles (Beltzer et al., 2005). Estas aves son importantes consumidores de los humedales y, por lo tanto, contribuyen a funciones importantes del ecosis tema, como acelerar los ciclos de nutrientes en las áreas de alimentación y regular las poblaciones de peces (Miranda & Collazo, 1997; Michelutti et al., 2010). Tienen adaptaciones morfológicas y fi siológicas relacionadas con la vida en los ambien tes acuáticos, como alargamiento relativo de las patas y picos largos y puntiagudos que utilizan para atrapar a sus presas (Beltzer et al., 2005). Las características de estas aves reflejan los dife rentes hábitos de alimentación y adaptaciones a las diversas condiciones ecológicas. Las especies con mayor tamaño corporal y del pico pueden capturar e ingerir presas más grandes (Beltzer et al., 2005). Se ha demostrado que existe relación entre la estructura de los diferentes segmentos del tubo digestivo (e.g., lengua, estómago), el tipo de ingesta de alimentos y los hábitos de las aves (Sabry, 2015; Al- Ahmady Al-Zahaby, 2016). Si bien, existen antecedentes referidos a la descrip ción de la anatomía macroscópica e histológica del tubo digestivo de garzas (Beltzer et al., 1987; De Carlo et al., 1988; Mosso et al., 1994; Rosa de Montaner et al., 1997; L-Ahmady Al-Zahaby, 2016; Mildren, 2020), en general, son escasos los trabajos sobre estas aves (Sabry, 2015).
El Hocó colorado Tigrisoma lineatum (Boddaert, 1783) se distribuye desde el sur de México al norte y centro de Argentina y es con siderada una especie rara o poco común en todo su rango de distribución (Martínez-Vilalta et al., 2020). Si bien su alimentación es poco conocida, se sabe que incluye peces (Martínez-Vilalta et al., 2020), insectos acuáticos, anfibios, crustáceos, serpientes acuáticas (Beltzer, 1990; Kushlan & Hancock, 2005), lagartos y pequeños mamíferos (Sovrano et al., 2019). El objetivo de este traba jo fue describir macroscópica e histológicamente los diferentes segmentos del tubo digestivo de Tigrisoma lineatum e interpretarlas en rela ción con su biología alimentaria (Beltzer, 1990; Sovrano et al., 2019).
MATERIAL Y MÉTODOS
El material biológico empleado en este estu dio corresponde a muestras preparadas de los distintos segmentos del tubo digestivo de adultos del Hocó colorado (Tigrisoma lineatum) pertene cientes a la colección del Instituto de Limnología (UNL-CONICET) de la provincia de Santa Fe (00786-00790, 00931-00935). Este material fue obtenido de los humedales del río Paraná Medio durante las décadas de 1980 y 1990. Las capturas fueron efectuadas con fines científicos y contaron con la autorización del Ministerio de Ambiente y Ganadería de la provincia de Santa Fe. Al mo mento de la captura, se obtuvieron medidas mor fométricas de los ejemplares (n=10) como longi tud total (distancia entre la cabeza y el extremo distal de la cola) y culmen (desde la base hasta el extremo distal) con un calibre (± 0,1 mm) y con una regla metálica (± 0,1 mm) en base a los crite rios de Baldwin et al. (1931). Los tubos digestivos de las aves colectadas fueron estudiados median te observación directa (macroscópica), con mi croscopio estereoscópico en fresco y fotografiados (n=10). Se obtuvieron medidas del tubo digestivo total (desde pico y la cavidad bucofaríngea hasta la cloaca) y de cada tramo con compás de pun ta seca y regla metálica (± 0,1 mm). Para lograr su conservación fueron fijados en formol 10% (n= 10) y conservados en etanol. Para las obser vaciones histológicas se utilizaron seis de estas muestras y se siguieron las técnicas usuales de fijación, deshidratación e inclusión en parafina. Se realizaron cortes de 3 a 6 μm de grosor para a su montaje definitivo y su posterior observación en el microscopio. Las muestras se examinaron y fotografiaron utilizando estereomicroscopio (Nikon 2500). Se realizaron tinciones mediante la técnica de corriente con hematoxilina-eosina (H-E) según el criterio de Celani de Basi et al. (1984). Se calculó el coeficiente intestinal (RI) que refleja la relación entre la longitud del tubo digestivo (pico-cloaca) y la longitud del ave (pico a base de cola).
Los valores se reportan como media ± desvia ción estándar (SD).
RESULTADOS
El tubo digestivo comienza con el pico y la cavidad bucofaríngea y continúa en un largo esó fago distensible. El esófago conduce el alimento a un estómago glandular y luego al estómago mus cular. El divertículo pilórico comunica a este últi mo con el primer tramo del intestino delgado. El límite del intestino delgado es un ciego pequeño y simple. El recto es corto y termina en la cloa ca, que se abre al exterior por la abertura cloacal (Fig. 1). No se hallaron variaciones intraespecífi cas en las estructuras descriptas. La longitud del tubo digestivo fue de 1827 ± 3,88 mm en prome dio y la longitud total de los individuos fue de 700 ± 2,02 mm.
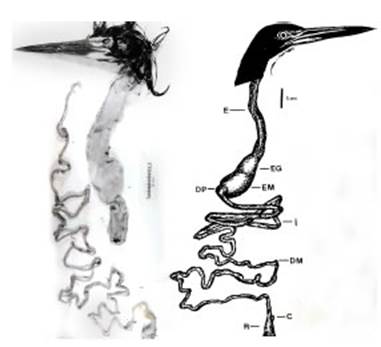
Fig. 1 Tubo digestivo de Tigrisoma lineatum. Abreviaturas: C, ciego; DM, divertículo de Meckel; DP, divertículo pilórico; E, esófago; EG, estómago glandu lar; EM, estómago muscular; I, intestino; R, recto.
Los ejemplares de T. lineatum poseen un pico rectiforme y longirrostro con un extremo pun tiagudo (Fig. 1), con valor promedio de culmen de 140 ± 2,35 mm. La lengua es estrecha y con forma de pirámide triangular, aguzada y lisa. La mucosa lingual tiene un epitelio plano estratifi cado queratinizado, con igual desarrollo en am bas caras hacia el extremo de la lengua, en tanto que, con menor desarrollo en la cara inferior en la base de la lengua. Las papilas están más desa rrolladas en la cara dorsal de la base lingual. El corion de tejido conectivo es denso con presencia de glándulas tubulosas compuestas y numerosas. El soporte es cartilaginoso y se observan tres partes en la base, rodeadas por fibras musculares estriadas longitudinales y oblicuas que disminu yen hacia el ápice. En el ápice está presente una única porción constituida por la prolongación del cartílago endogloso y escasas fibras musculares.
El esófago tiene paredes extensibles y posee una longitud promedio de 230 ± 4,5 mm. Se ob servó que las paredes son extensibles puesto que se pliegan cuando el esófago estaba vacío y alcan za una capacidad máxima de almacenamiento, sin dañarse, cuando contenían presas. La mucosa posee un epitelio plano estratificado no querati nizado en toda su extensión con numerosas glán dulas mucosas tubuloalveolares, que se abren a la luz del órgano mediante conductos simples. La lámina de tejido conectivo (corion) es gruesa con vasos sanguíneos y determina los pliegues en el órgano vacío. La capa muscular de la mucosa pre senta fibras longitudinales con escaso desarrollo. La túnica muscular tiene mayor desarrollo con fibras circulares internas. La adventicia es una fina capa de tejido conectivo.
El estómago tiene forma de saco y es alar gado, está formado por el estómago glandular o proventrículo, el estómago muscular o ventrículo y el divertículo pilórico (Fig. 2).

Fig. 2 Estómago de Tigrisoma lineatum. Abreviaturas: E, esófago; EG, estómago glandular; EM, estómago muscular.
El estómago glandular se distingue externa mente por su color levemente más oscuro que se corresponde con la presencia de las glándulas secretoras. En un corte sagital, se observan el pa quete de estas glándulas en paredes que oscilan entre los 2 mm (estómago repleto con 80 cc) y 6 mm (estómago vacío, valores obtenidos en un estómago con un centímetro cúbico de conteni do estomacal) conformando una faja de unos 25 mm de ancho dispuestas de manera trasversal siguiendo el sentido longitudinal del tubo diges tivo. La túnica mucosa tiene pliegues longitu dinales y epitelio cilíndrico simple con glándulas tubuloalveolares simples (glándulas proventricu lares) profusas en el corion, separadas por cordones de tejido conectivo y, en las que se observan los conductos abiertos a la luz del órgano (Fig. 3). La submucosa es poco visible. La túnica muscu lar está bien desarrollada y se observa como una capa de fibras longitudinales delgadas externa y otra de fibras circulares interna, más amplia. La adventicia o serosa es una capa de tejido conjun tivo laxo (Fig. 3). No se observa el istmo (separa ción entre el estómago glandular y el estómago muscular).
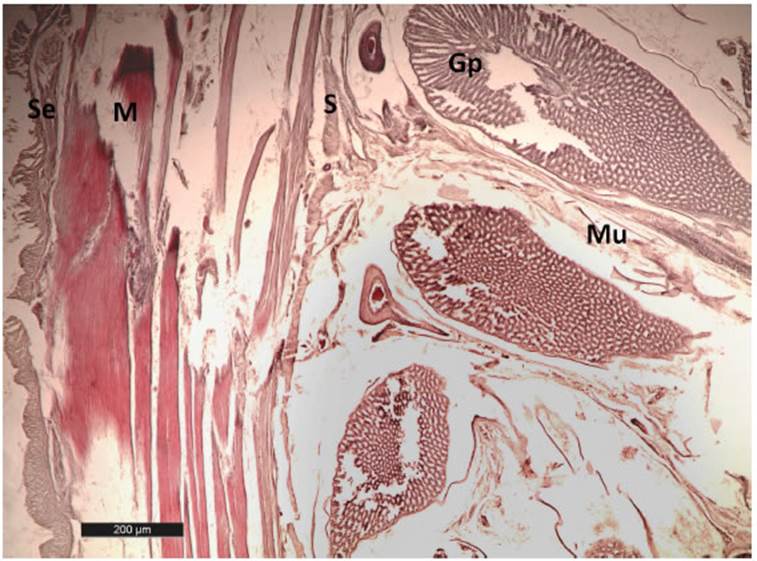
Fig. 3 Corte histológico del estómago glandular (proventrículo) de Tigrisoma lineatum. Abreviaturas: Gp, glán dula proventricular; M, muscular; Mu, mucosa; S, submucosa; Se, serosa. H-E 40X.
El estómago muscular presenta paredes rela tivamente delgadas de 2 mm en estómago reple tos y de 4 mm en estado vacío. La túnica mucosa tiene pliegues con epitelio cilíndrico simple con glándulas tubulares que se abren a la luz. La tú nica muscular es gruesa y posee fibras oblicuas internas, una capa con fibras circulares que la ro dea, bien desarrollada y, una capa de fibras longi tudinales externa. La túnica adventicia presenta las mismas características con respecto al estó mago glandular (Fig. 4). El divertículo pilórico se diferencia por su aspecto globular con pliegues, sirve de conexión entre el estómago muscular y el primer tramo del intestino delgado (unión ventrículo-duodeno). La mucosa tiene glándulas tubulares simples. La capa muscular está desa rrollada.

Fig. 4 Corte transversal del estómago muscular (ventrículo) de Tigrisoma lineatum. Abreviaturas: M, muscular; Mu, mucosa; Se, serosa. H-E 40X.
La longitud total promedio del intestino es de 1390 ± 20 mm. El intestino delgado tiene una longitud de 1386 ± 20 mm con aproximadamente 20 asas y un diámetro que oscila entre 2,5 y 4 mm. La primera porción del intestino (duodeno) es relativamente larga y consta de una extensa asa que contiene al páncreas, sin diferenciar se morfológicamente de la parte posterior del intestino (yeyuno e íleon). La mucosa presenta numerosas vellosidades finas con epitelio cilín drico simple alto y células caliciformes y glándu las tubulosas simples formadas por invaginación del epitelio (criptas de Lieberkhün). El corion de tejido conectivo laxo se ubica entre las mencio nadas criptas. La lámina muscular de la mucosa es delgada con fibras musculares lisas. La túnica muscular consta de una capa de fibras circula res interna desarrollada y una capa longitudinal externa delgada. La serosa presenta las mismas características con respecto a las presentes en el estómago (Fig. 5).

Fig. 5 Corte histológico de la primera porción del intestino (duodeno) de Tigrisoma lineatum. Abreviaturas: ME, muscular externa; MI, muscular interna; Mu, mucosa; Se, serosa. H-E 40X.
En el límite entre el intestino delgado y el grueso está presente un único y rudimentario ciego, de 2 ± 0,2 mm de longitud. Este presenta una capa muscular gruesa y abundante infiltra ción linfática. El recto presenta una longitud de 40 ± 2 mm y un diámetro de 5 mm. La mucosa posee un epitelio cilíndrico simple con vellosida des más gruesas y bajas que el intestino delgado. La lámina muscular es una capa gruesa con capa interna circular desarrollada y una capa externa longitudinal con menor desarrollo.
El divertículo de Meckel se observa aproxi madamente a mitad del recorrido del intestino, presentando una longitud de 2 ± 0,2mm. La mu cosa posee un epitelio glandular con abundante infiltración linfática. La capa muscular se obser va con predominio de fibras circulares y con no table tejido conectivo en la serosa.
El valor del coeficiente intestinal (RI) fue de 2,61 ± 0,27.
DISCUSIÓN Y CONCLUSIONES
Nuestra descripción del tubo digestivo de Tigrisoma lineatum es similar a descripciones efectuadas para otras especies de garzas en los humedales del río Paraná (Garcita bueyera Bulbucus ibis, Beltzer et al., 1987; Garza blan ca Ardea alba y Garcita blanca Egretta thula, De Carlo et al., 1988; Garza bruja Nycticorax nyctico rax, Mosso et al., 1994; Garcita azulada Butorides striata, Rosa de Montaner et al., 1997).
El pico, la lengua y la cavidad bucofaríngea funcionan para obtener e ingerir las presas. El desarrollo de la lengua alargada, delgada y con su extremo puntiagudo en garzas podría estar relacionado con el ajuste de presas pequeñas en el momento de la deglución (Mosso et al., 1994). El diseño del pico y lengua incrementa la eficacia durante las capturas mediante la implementa ción de ajustes para los distintos tipos y tamaños de presas (versatilidad trófica), lo que se eviden cia en el amplio rango de ítems alimentarios ha llados en estudios sobre la dieta de Tigrisoma li neatum (Beltzer, 1990; Kushlan & Hancock 2005; Sovrano et al., 2019). La presencia de glándulas linguales tubulares contribuye humedeciendo la cavidad bucofaríngea, que facilita el tránsito de las presas (Samar et al., 2017).
El largo y extensible esófago posee funciones de transporte y le permite actuar como órgano de almacenamiento de los alimentos cuando el estómago está repleto o cuando las presas son voluminosas (Mosso et al., 1994), lo que acuerda con el hallazgo de presas abundantes o grandes durante la disección. En este sentido, los grandes pliegues de la mucosa dejan una luz en el centro del esófago que, ante la ausencia del buche, per miten almacenar el alimento a lo largo del esófa go (McLelland, 1989).
El estómago glandular tiene un considerable desarrollo con abundantes glándulas. El estóma go muscular, en cambio, presenta escaso desa rrollo de las capas musculares comparado con el estómago grande de especies granívoras y que en su interior puede contener gastrolitos, que ayu dan a la digestión mecánica (Zanuzzi & Barbeito, 2014). Las dimensiones del estómago glandular y la secreción de las glándulas pueden relacionar se con la ingesta de peces (Zanuzzi & Barbeito, 2014) u otras presas enteras, lo que requiere una importante digestión química. En esta especie, como en otras aves acuáticas, no es posible dis tinguir macroscópicamente la región del istmo (Zanuzzi & Barbeito, 2014). El estómago mus cular actúa como cámara receptora de los ali mentos impregnados de las secreciones gástricas que continúan el proceso de digestión química y que es necesario para efectuar una compleja di gestión en aves carnívoras (Abumandour, 2014; AbdElnaeem et al., 2019).
En algunas aves acuáticas como cormoranes (Phalacrocoracidae), patos (Anatidae) y gansos (Anseriformes) existe un estómago pilórico con grandes pliegues que funcionan como filtro para retener los alimentos (Zanuzzi & Barbeito, 2014). De manera similar, en T. lineatum se observó un engrosamiento de las paredes musculares en las proximidades del divertículo, que actuaría como un esfínter de retención y evitando el paso al in testino de piezas no digeribles (e.g., exoesquele tos). Estas características coinciden con lo des cripto para otras especies de garzas (Beltzer et al., 1987; Mosso et al., 1994).
El divertículo de Meckel es un vestigio de la vida embrionaria, que en el adulto se transfor ma en un órgano linfoide (Alvarado Pinedo & Barbeito, 2014). Este órgano pertenece a los tejidos linfoides asociados al tubo digestivo y cumple la función de reforzar el sistema inmune del ave (Oláh & Vervelde, 2008). El grado de desarrollo de los ciegos ha sido relacionado con la dieta, con ciegos muy desarrollados en especies omnívoros y herbívoras, ciegos vestigiales no funcionales en especies granívoras, en su mayoría vestigiales y no glandulares, pero con una variación extrema en el grado de desarrollo en las especies insectí voras y ciegos vestigiales o ausentes en especies piscívoras (Naik & Dominic, 1962; McLelland, 1989). El ciego único y relativamente pequeño de garzas piscívoras, fue observado por Clench & Mathias (1995) como un rasgo característico dentro de esta familia. Un estudio reciente en el que se compararon los ciegos de siete especies de garzas, encontró que estos órganos se hallan de manera singular y son pequeños en especies predominantemente piscívoras (Gran garza azul Ardea herodias, Gran garza blanca A. h. occi dentalis) (Mildren, 2020). En T. lineatum se des cribió un único ciego, corto y poco desarrollado con infiltración linfoide que, por lo dicho ante riormente para otras especies de la familia se re laciona con su dieta y puede tener una función inmunológica según las observaciones realizadas en este trabajo.
El coeficiente intestinal obtenido presenta un valor intermedio con respecto al de las especies piscívoras, con coeficientes elevados en Cormorán neotropical Nannopterum brasilianum (RI=3,10) (Beltzer et al., 1987) y aquellos carnívoros no pis cívoros como Bulbulcus ibis (RI=1,96) (Beltzer et al., 1987), ubicándose esta especie entre las garzas que poseen un amplio espectro trófico en las que si bien los peces constituyen una fracción importante de la dieta, incorporan además otros taxa como reptiles, pequeños mamíferos, anfibios e insectos (Beltzer, 1990; Kushlan & Hanconck, 2005; Salvador & Fariña, 2018; Sovrano et al., 2019).
Este trabajo es la primera descripción ma croscópica e histológica del tracto digestivo de T. lineatum. Esta información es útil para esta blecer generalizaciones y relaciones con aspectos ecológicos como la dieta o también, identificar características específicas de adaptaciones mor fofuncionales y parentescos filogenéticos entre especies.

