










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Asociación Argentina de Sedimentología
Print version ISSN 1853-6360
Rev. Asoc. Argent. Sedimentol. vol.5 no.2 La Plata Dec. 1998
ARTÍCULOS
Estructuras biogénicas en el Cenozoico tardío de Mar del Plata (Argentina) atribuibles a grandes mamíferos 1
Marcelo A. Zárate *, María Susana Bargo +, Sergio F. Vizcaíno +, Alejandro Dondas # y Orlando Scaglia #
* CONICET/IANIGLA-CRICYT, CC 330, 5500 Mendoza, República Argentina.
+ Departamento Científico Paleontología Vertebrados, Museo de la Plata, Paseo del Bosque s/n, 1900 La Plata, Buenos Aires, República Argentina.
# Museo Municipal de Ciencias Naturales "Lorenzo Scaglia", Plaza España, 7600 Mar del Plata, Buenos Aires, República Argentina
1 Contribución presentada a la Primera Reunión de Icnología del Mercosur y Tercera Reunión Argentina de Icnología. Mar del Plata, 1998.
Recibido: 1º de diciembre de 1998.
Aceptado: 20 de agosto de 1999.
Resumen
Las estructuras biogénicas atribuibles a mamíferos grandes son muy frecuentes en los depósitos del Cenozoico tardío de Mar del Plata. Son estructuras cerradas, discordantes en relación con las unidades estratigráficas hospedantes; presentan sección transversal subcircular a ligeramente elíptica con diámetros variables entre 0,75 y 1,80 m. Los rellenos sedimentarios están usualmente compuestos por arenas muy finas y limos, ya sea estratificados o macizos, que alternan con niveles de arcillas laminadas. Las estructuras, interpretadas como cuevas y galerías, se hallan excavadas en sustratos pelíticos o arenosos finos, ya sea de planicies de inundación o interfluvios. La distribución estratigráfica abarca desde el Plioceno hasta quizás el Pleistoceno tardío final (Máximo Glacial?, Tardiglacial?). Los principales criterios de reconocimiento son las relaciones estratigráficas, dimensiones y contornos cerrados. Los rellenos sedimentarios y los contenidos faunísticos son criterios útiles que deben emplearse en relación contextual. Se las ha atribuido a mamíferos del Orden Xenarthra, en especial armadillos de gran tamaño como Pampatherium (Pampatheriidae), Eutatus (Dasypodidae) y Propraopus (Dasypodidae). Análisis anatómicos, alométricos y biomecánicos efectuados en milodóntidos pleistocenos (Scelidotherium, Glossotherium y Lestodon) sugieren que estos mamíferos fósiles deben considerarse también como probables constructores de estas estructuras. Algunas huellas encontradas en túneles desprovistos de relleno sedimentario, se adaptan a los dos dedos más desarrollados de las manos de los Scelidotherium y Glossotherium. Se infiere que esta fauna fosorial de mamíferos grandes debe haber desempeñado un papel destacado como agente de erosión, al generar volúmenes importantes de material sedimentario. Al tratarse de estructuras estratigráficamente transgresivas, el valor como indicadores paleoambientales así como su acotamiento cronológico están limitados, aunque pueden ser útiles para comprender la historia diagenética de los depósitos.
Palabras clave: Argentina; Buenos Aires; Cenozoico tardío; Estructuras biogénicas; Grandes mamíferos.
Extended abstract
The occurrence of mammals caves and burrows is a notable feature of the Late Cenozoic deposits of the Mar del Plata area (Fig. 1). Although these structures have been known for more than a hundred years, few studies have been devoted to their description and interpretation. This paper is focused on the morphological characteristics, the stratigraphic setting and the field criteria of recognition of the biogenic structures attributed to big mammals. We discuss the associated fauna and their probable builders and evaluate their importance as stratigraphic and sedimentological indicators.
The structures possess discrete boundaries and subcircular to slightly elliptical cross-sections. Longitudinal sections are also very frequent, but not usually reported, possibly because they have been misinterpreted as features of fluvial deposition. Their maximum width (horizontal axis) which usually exceeds the height slightly, varies between 0.75 m and 2 m (Fig. 2).
Some structures show flat floors and arched roofs (Fig. 3f). Although few structures have been measured (n=42) they tend to fall into two sizes ranges. The maximum width of most ranges from 80 cm to 130 cm whereas many of the remaining structures are approximately 180 cm in width (Fig. 2). The major size variation might reflect different species of diggers; size variations between 15-30 cm are possibly caused by measurement of width along a plane oblique to the longitudinal axis of the structure (due to difficulties in recognizing the true axis); those between 5 cm and 10 cm are attributable to measurement error caused by boundaries that are difficult to distinguish. Recently, several tunnels, reaching up to 1.8 m in width and 40 m in length and devoid of sedimentary filling, have been discovered in the Mar del Plata area. The tunnels are interpreted as animal burrows.
The studied biogenic structures were formed in very fine sandstones or siltstones (loess-like sediments) and are located either in floodplain or interfluvial paleoenvironments. The tunnels are filled with thinly stratified fine sands and silts alternating with laminated clay layers. Scutes of Dasypodidae (Zárate, 1989) and disarticulated remains of rodents have occasionally been recovered from the sedimentary filling.
Temporally, the structures are found from the Pliocene to the late Pleistocene, and possibly extended into Late Glacial Maximum-Late Glacial times. The main criteria for field recognition of these structures are their stratigraphic relationships, sizes and closed boundaries. Sedimentary fillings and associated faunal remains are further criteria that may be applied once the contextual framework has been established based on the main criteria. Frenguelli (1921) attributed these structures to Glyptodontidae (Glyptodon, Doedicurus, Neuryurus). More recently, Imbellone & Teruggi (1988), Imbellone et al. (1990) and Quintana (1992) attributed them to large armadillos such as Pampatherium (Pampatheriidae), Eutatus (Dasypodidae) and Propraopus (Dasypodidae). Anatomical, allometric, and biomechanical analysis recently performed on Pleistocene milodontids (Scelidotherium, Glossotherium, Lestodon) indicate that their limbs were well prepared for digging (Bargo et al., submitted). Also, the diameters of the structures coincide with or slightly exceed the inferred diameters of Scelidotherium and Glossotherium, and supports the interpretation that the structures were excavated by these mylodontids. Furthermore, the scratchs found on the walls and roof of a burrow devoid of sedimentary filling in Mar del Plata and originally attributed to large armadillos by Quintana (1992), are better explained as having been formed by the two more developed unguals digits of Scelidotherium and Glossotherium.
The paleoenvironmental implications of these biogenic structures remain limited, as the ecological reasons for their construction have not been fully elucidated. The structures may be useful in the interpretation of the diagenetic history of the deposits. The burrows and caves may have been at least 5-6 m deep based on the shallowest empty burrows found in Mar del Plata, and thus traverse different stratigraphic and age units. This transgression of litho- and chronostratigraphic units renders problematic the determination of their time of origin and relative abundance at any stratigraphic level. Based on the size and abundance of the structures, we infer that the Pleistocene fossorial fauna of large mammals must have played a significant role as potential agents of erosion, generating substantial volume of sediments.
Key words: Argentina; Buenos Aires; Late Cenozoic; Biogenic structures; Big mammals.
INTRODUCCIÓN
La ocurrencia de cuevas y galerías de mamíferos es una característica de los depósitos del Cenozoico tardío de la Provincia de Buenos Aires, señalada por diversos autores desde comienzos del siglo XX (Ameghino, 1908; Frenguelli, 1928; Kraglievich, 1952). A pesar de que su existencia es conocida desde hace largo tiempo, se han efectuado muy pocos estudios específicos. Recientemente se investifaron las cuevas del roedor fósil Actenomys (Genise, 1989; Scognamillo, 1993) muy frecuentes en los afloramientos del Plioceno de Chapadmalal (Fig. 1). Las estructuras subcirculares de mayores dimensiones (mayores a 0.70 m) se han atribuido a mamíferos grandes, aunque no se demostraron los criterios empleados para sostener tal interpretación. Los escasos antecedentes existentes sobre estas estructuras conocidas con el nombre de crotovinas, término que también incluye a las de pequeñas dimensiones, se refieren a las características y mecanismos de depositación de los rellenos sedimentarios (Imbellone & Teruggi, 1988; Imbellone et al., 1990). También se ha descripto la morfología de una galería vacía encontrada en Mar del Plata (Quintana, 1992).
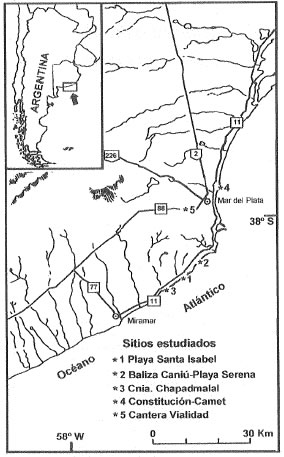
Figura 1. Mapa de ubicación de los sitios estudiados.
Figure 1. Map showing localities of study.
Los objetivos de esta contribución son analizar la morfología, distribución estratigráfica y génesis de las estructuras atribuibles a mamíferos grandes, proponer criterios de reconocimiento, discutir la asignación faunística y probables constructores y evaluar su importancia como indicadores estratigráficos y sedimentológicos.
CARACTERÍSTICAS GENERALES
Morfología.
Las mediciones de los diámetros máximos horizontales y verticales se efectuaron en 42 estructuras (Fig. 2) de los sitios Playa Santa Isabel, Baliza Caniú-Playa Serena, tramo Constitución-Camet, sección Colonia Chapadmalal, y Cantera de Vialidad (Fig. 1). Presentan límites discretos y sección transversal subcircular a ligeramente elíptica, de acuerdo con la orientación (perpendicular u oblicua) de la exposición (Figs. 3a, b y e); en varios casos se reconocieron pisos planos y techos y paredes abovedados (Fig. 3f). Son ligeramente más anchas que altas; los diámetros máximos (ejes horizontales) oscilan entre 75 cm y 200 cm (Fig. 2). Si bien el número de cuevas es todavía escaso, la distribución de frecuencias muestra una tendencia a agruparse en dos tamaños principales. Así, el mayor número de estructuras presenta un diámetro horizontal máximo variable entre 100 cm y 130 cm con una moda secundaria en alrededor de 170 cm-180 cm. Estas diferencias en tamaño tal vez reflejen distintos animales cavadores, mientras que las variaciones menores (15 cm-20 cm) podrían ser consecuencia de la orientación de las secciones medidas y errores en la medición (estimados en 5 cm-10 cm) por imprecisión en la ubicación de los límites. Aunque también son frecuentes, no hay referencias de exposiciones longitudinales (Fig. 3d), quizás debido a la dificultad para reconocerlas como tales, ya que si los rellenos son estratificados, es muy probable que en los levantamientos de campo se las interprete como estructuras fluviales de canal o lentes palustres. También son frecuentes las estructuras con rellenos sedimentarios parcialmente erosionados, lo que permite identificarlas como galerías (Figs. 3c y e).
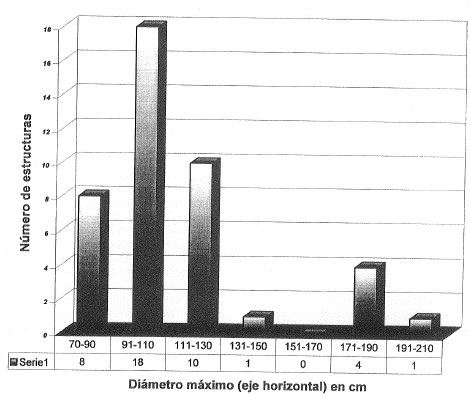
Figura 2. Histograma de número de estructuras en función de los diámetros máximos (eje horizontal).
Figure 2. Histogram of the distribution of structures by maximum width.

Figura 3. Estructuras biogénicas de mamíferos grandes en Playa Santa Isabel (Formación Vorohué, Kraglievich 1952; Aloformación Punta San Andrés, Miembro inferior, Zárate 1989). (ps) paleosuelo, (ts) tosca. a) vista general de la sección y estructura biogénica; b) detalle de a), erosión diferencial del relleno; c) pared de estructura erosionada en afloramiento; d) sección longitudinal; e) estructura exhumada por erosión marina de sección subcircular; f) estructura de piso plano y forma abovedada con relleno sedimentario de arenas muy finas y limos estratificados.
Figure 3. Biogenic structures of big mammals at Playa Santa Isabel (Vorohué Formation, Kraglievich, 1952; Punta San Andrés Alloformation, lower member, Zárate 1989) (ps) paleosol, (ts) “tosca” (calcrete); a) general view of the stratigraphic section and a biogenic structure; b) close up of a), differential erosion of the sedimentary filling; c) wall remains from an eroded structure; e) subcircular structure exhumed by marine erosion; f) flat floor and arched roof structure with sedimentary filling made up of very fine sands and silts.
En los últimos años, varios túneles de grandes dimensiones (hasta 1,80 m de diámetro y 40 m de longitud), sin relleno sedimentario, han sido descubiertos en el área de Mar del Plata durante la construcción de obras civiles. Si bien hay reportes orales de cavidades vacías encontradas en la zona, la primera descripción morfológica efectuada corresponde a una paleocueva aparecida en la zona de Punta Mogotes en 1987 que incluye marcas atribuibles a rasguños en techo y paredes (Quintana, 1992). Con posterioridad, se han registrado por lo menos otros 5 túneles de características similares que aparecieron a profundidades de 5-6 m y hasta 12 m con respecto a la superficie actual.
Sustratos y ambientes.
Las estructuras biogénicas están excavadas en sustratos pelíticos o arenosos finos con moderado a alto grado de cohesión. Los ambientes sedimentarios donde se las encuentra son muy variados; incluyen planicies de inundación o bien interfluvios. Atraviesan tanto niveles de paleosuelos como materiales poco reorganizados por pedogénesis (Fig. 3). Hasta el momento no se ha observado una vinculación particular entre las estructuras y los paleoambientes. Sólo se ha registrado una mayor abundancia en algunos tramos de niveles estratigráficos correspondientes a planicies de inundación, pero ello puede responder a problemas de reconocimiento e identificación en los afloramientos.
Rellenos sedimentarios.
Imbellone & Teruggi (1988) e Imbellone et al. (1990), analizaron la sedimentología de los rellenos de tres estructuras-tipo del Pleistoceno ¨sensu lato¨ del área de La Plata y propusieron diferentes mecanismos de transporte y sedimentación para explicar la colmatación de las cavidades, tales como percolación de aguas circulantes, inundaciones, acción gravitatoria, acción eólica. Básicamente los rellenos sedimentarios están compuestos por arenas finas y limos estratificados o megascópicamente de aspecto macizo, que alternan con niveles de arcillas laminadas y depósitos de brechas intraformacionales. En un caso se exhumaron del relleno placas de dasipódidos (Playa Santa Isabel, Zárate, 1989). En otra, se encontraron restos desarticulados de roedores y estructuras de bioturbación de invertebrados. Las estructuras halladas en niveles del Plioceno (Chapadmalalense) pueden estar atravesadas por cuevas de roedores (Actenomys). Las acumulaciones de tosca suelen ser frecuentes en los contornos de las estructuras.
Distribución estratigráfica.
Las estructuras en cuestión presentan una amplia distribución estratigráfica que abarca desde el Plioceno hasta el Pleistoceno tardío. Existen variaciones en sus abundancias relativas tanto en sentido vertical como horizontal. Frenguelli (1928) señala que, en términos comparativos, las madrigueras grandes predominan en los depósitos del “Prebelgranense” (Marplatense, Ensenadense) en relación con el Preensenandense (Montehermosense-Chapadmalalense), asignando la dife-rencia a los cambios faunísticos ocurridos. Según Frenguelli (1928), en el “Prebelgranense” han desapare-cido la mayor parte de las formas de mamíferos pequeños, los que fueron reemplazados por formas grandes. Por su parte, Kraglievich (1952) menciona “la presencia de cuevas grandes en el nivel II de Vorohué (Marplatense) de hasta 1 metro de diámetro”, (Fig. 3) característica también señalada como rasgo peculiar en investigaciones estratigráficas más recientes (Zárate, 1989).
CRITERIOS DE RECONOCIMIENTO
Las estructuras son cerradas y discordantes en relación con los depósitos hospedantes ya que truncan los niveles estratigráficos. En sustratos de rocas calcáreas o compuestas por otras sales, que no componen los afloramientos en cuestión, la disolución puede generar cavidades aunque de morfología diferente a las estructuras cerradas que consideramos. Los materiales sedimentarios del Cenozoico tardío son de composición silicoclástica y los procesos sedimentarios que actuaron han originado estructuras de erosión de contornos abiertos. En estos materiales, la meteorización, en particular la formación de suelos, y la diagénesis, involucran principalmente procesos de oxidación, reducción, translocación de material fino, precipitación de calcáreo y sólo muy localizadamente disolución sobre las superficies de las costras de toscas.
Desde el punto de vista biológico, la ocurrencia de restos fósiles incluidos en el relleno sedimentario es un criterio de reconocimiento comúnmente empleado (Frenguelli, 1928; Kraglievich, 1952). Sin embargo, la sola presencia de restos, por el momento ocasionales, en las cuevas de grandes dimensiones no es evidencia suficiente para efectuar la asignación de éstas a la especie encontrada pues pueden registrarse allí por otros factores (e.g. haber sido introducidos por acción de aguas circulantes o predadores, o haber buscado refugio en un túnel fabricado por individuos de otras especies). Por el contrario en el caso de cuevas de pequeñas dimensiones del Plioceno este criterio ha sido de gran utilidad ya que la presencia de porcentajes muy elevados (75%) del género fósil Actenomys en su interior (Genise, 1989), así como el alto grado de articulación y preservación de los restos (Peña, 1997) reducirían considerablemente la posibilidad de que los restos de Actenomys hayan sido introducidos aleatoriamente durante el proceso de relleno o bien que las habitaran circunstancialmente. Asimismo, implícitamente, se ha utilizado el método analógico para comparar cualitativamente los patrones resultantes con los generados por la actividad de la fauna fosorial actual, con los cuales muestran similitud.
En ciertas situaciones, cuando las estructuras están parcialmente truncadas por superficies de erosión de canales fluviales, la interpretación no es tan clara. Aparecen como depósitos de geometría lenticular, pero las paredes laterales son sensiblemente cóncavas y los contornos muy regulares. En estos casos, se asemejan a las estructuras erosivas producto de escurrimiento encauzado.
Si se consideran los rellenos sedimentarios, las estructuras erosivas hídricas de los ambientes del Cenozoico tardío en cursos de orden menor y fuera de los ámbitos serranos, presentan material psefítico en la base (grava o gravilla residual compuesta por intraclastos y clastos de tosca) que indica la acción del escurrimiento encauzado. Las estructuras biogénicas exhiben rellenos sedimentarios generalmente estratificados o bien masivos sin material grueso asignable a carga de lecho, o bien incluyen fracción psefita de intraclastos angulares del material hospedante en el que están excavadas.
Los criterios de reconocimiento fundamentales son las relaciones estratigráficas, dimensiones y contornos; los rellenos sedimentarios y los eventuales contenidos faunísticos son criterios importantes que deben emplearse en relación contextual.
FAUNA ASOCIADA
En el caso de las estructuras de grandes dimensiones varias veces fueron atribuidas a mamíferos del Orden Xenarthra, un grupo altamente característico de la fauna sudamericana que durante el Plioceno y, especialmente el Pleistoceno, desarrollaron formas de gran tamaño en varios linajes. Frenguelli (1921, pag. 343) menciona entre ellos a los gliptodontes Glyptodon, Doedicurus, Neuryurus (Cingulata, Glyptodontidae) y señala que las cuevas “frecuentemente contienen todavía los restos del esqueleto del animal que los habitaba”. En el Balneario Centinela del Mar (Baliza del Malacara) Frenguelli (1928, 1955) señala la existencia de una cueva rellena con ceniza volcánica, que asigna al perezoso terrestre Scelidotherium (Tardigrada, Mylodontidae), al encontrar restos del mismo en su interior.
Kraglievich (1934) y Quintana (1992) descartan a los gliptodontes debido a la imposibilidad funcional de los gliptodontes para la excavación.
En trabajos recientes, estas estructuras se atribuyen solamente a armadillos de gran tamaño. Imbellone & Teruggi (1988), Imbellone et al. (1990), a partir de una comunicación personal de Eduardo Tonni, asignan las paleocuevas de La Plata tentativamente a Pampatherium (Pampatheriidae) y Eutatus (Dasypodidae). Quintana (1992) propone a Pampatherium, Eutatus o Propraopus (Dasypodidae) por comparación entre el diámetro corporal y el diámetro transverso de las cuevas. El hábito fosorial está ampliamente difundido entre los representantes actuales del grupo.
Excepto la mención de Frenguelli (1928, 1955) de la asociación entre una paleocueva y los perezosos fósiles, nunca se consideró a estos xenartros como probables contructores de las mismas.
Sin embargo, varios autores (Owen, 1842, 1851; Winge, 1941; Cuenca Anaya, 1995; White, 1997) destacaron que los perezosos milodóntidos presentaban en sus miembros adaptaciones para excavar, probablemente utilizadas para desenterrar alimento de origen vegetal.
Análisis anatómicos, alométricos y biomecánicos efectuados en milodóntidos pleistocénicos (Scelidotherium, Glossotherium y Lestodon) también indican que sus miembros estaban perfectamente diseñados para excavar (Bargo et al., enviado).
Algunas de las huellas encontradas en el túnel atribuido a armadillos por Quintana (1992) representan pares de surcos paralelos (Fig. 4b). Sin embargo, los surcos son demasiado grandes para ser atribuidos a los dasipódidos arriba mencionados. Aunque los pampaterios son de tamaño mayor que los dasipódidos, sus manos aún resultan demasiado pequeñas para esas huellas y presentan tres dedos bien especializados para cavar (Edmund, 1985), con lo que sería esperable encontrar trazas conformadas por tres surcos más o menos paralelos. Es sugestivo que el tamaño y forma de las huellas halladas en aquel túnel, se adapten a las manos de Scelidotherium y Glossotherium (Fig. 4a), que poseen los dígitos II y III más desarrollados con poderosas garras para cavar.
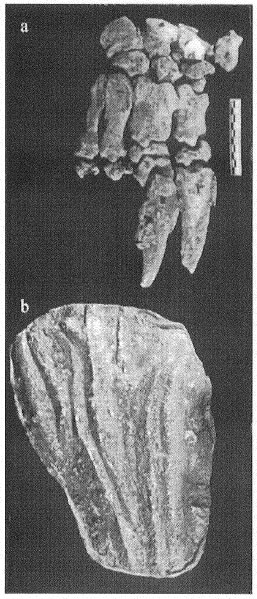
Figura 4. a) Autopodio derecho de Scelidotherium (MLP 3-574) y b) calco de las marcas atribuidas a rasguños hallada en el túnel descripto por Quintana (1992; MMP 2c).
Figure 4. a) Right manus of Scelidotherium (MLP 3-574), and b) cast of the tracks referred as scratches found in the tunnel described by Quintana (1992; MMP 2c).
Otra evidencia que apoyaría la interpretación de estos milodóntidos como constructores de algunas de las cuevas es el diámetro de las mismas, que en todo su recorrido coincide o supera el diámetro inferido para ambos perezosos (aproximadamente 1 metro). Los diámetros de los túneles construidos por mamíferos actuales se aproxima al diámetro corporal de su constructor (Hickman, 1990). Entre el resto de la fauna de mamíferos plio-pleistocenos no se encuentran animales cavadores de tamaño comparable al de estos milodóntidos.
La suma de la información analizada sugiere que los perezosos de la familia Mylodontidae deben ser agregados a la lista de los probables constructores de las grandes cuevas encontradas en el Plio-Pleistoceno de la Región Pampeana.
SIGNIFICADO ESTRATIGRÁFICO Y SEDIMENTOLÓGICO
El acotamiento cronológico de estas estructuras biogénicas, así como la interpretación de las variaciones de sus abundancias relativas, presenta varias limitaciones. En primer término, aún no es posible establecer cuál ha sido la profundidad de excavación con respecto a la superficie. Como orden de magnitud, a fin de efectuar inferencias, se podría suponer una profundidad cercana a los 5 -6 metros, partiendo de las cuevas y galerías vacías más superficiales, encontradas en Mar del Plata. En las secuencias aflorantes en Chapadmalal y Miramar, tales espesores involucran lapsos muy prolongados que pueden interesar niveles de distintas edades. Así, aunque las cuevas se encuentren excavadas, por ejemplo, en el Marplatense, podrían corresponder a un momento del Ensenadense. En consecuencia, la estimación de abundancias relativas por unidad estratigráfica debe ser reinterpretada debido al carácter transgresivo de las estructuras.
Por otro lado, en el caso de los túneles vacíos encontrados en Mar del Plata, a juzgar por el alto grado de preservación relativa que exhiben y la ausencia de relleno sedimentario, así como por el hecho de estar excavados parcialmente en depósitos eólicos del Pleistoceno tardío, es muy probable que se trate de estructuras relativamente recientes, generadas durante el último ciclo de glaciación, tal vez no más antiguas que el último máximo glacial o tardiglacial.
Otra limitación es el reconocimiento de las cuevas
en los afloramientos, pues está en función de la naturaleza de sus rellenos sedimentarios, los cuales eterminan su grado de visibilidad. En general, son registradas sólo aquellas estructuras con límites discretos, de manera que lo obtenido, en términos cuantitativos, son estimaciones mínimas.
Las variaciones en la abundancia relativa de fósiles de mamíferos grandes y pequeños debe considerarse en términos tafonómicos y por lo tanto en relación con las condiciones paleoambientales. Así, la dominancia de restos fósiles de especies pequeñas en el Plioceno, está relacionada con la preservación diferencial que ha ocurrido en el microambiente fosorial y con el proceso de sedimentación (Peña, 1997). En consecuencia, la interpretación de Frenguelli (1928) sobre el predominio de un determinado tipo de cueva no puede confirmarse por el momento, pues tal vez refleje una tendencia aparente.
Como estas estructuras son rasgos postdeposi-tacionales, estratigráficamente transgresivos en relación con las unidades hospedantes, no es posible asignarles valor paleoambiental a partir del análisis de las características de los niveles estratigráficos hospedantes. Para reconstruir condiciones paleoambientales sería necesario, por un lado, vincularlas con una determinada paleosuperficie, pero se desconocen las entradas y si éstas se ubicaban en la superficie del suelo, en barrancos naturales o en ambos. Por el otro, sería necesario comprender los requerimientos ecológicos de la fauna que las excavó. Este último aspecto resulta sumamente complejo y aún no ha sido desarrollado convenientemente. Debido a que estas cuevas son producto de condiciones posteriores a la sedimentación, podrían ser indicadores de la historia diagenética de los depósitos.
El papel desempeñado por esta fauna fosorial como agentes potenciales de erosión y sedimentación es un aspecto aún no evaluado. En ambientes actuales, los sedimentos acumulados en la entrada de las cuevas, generan localizaciones puntuales con alta disponibilidad potencial de material clástico que puede ser redepositado durante las lluvias o bien por el viento, tanto en la superficie como en el interior de las cuevas. En el caso de la fauna fosorial de mamíferos grandes que habría habitado la llanura pampeana, se debe pensar en volúmenes muy importantes de material removido y disponible en la entrada de las cuevas, que excedería enormemente las estimaciones basadas en pequeños animales cavadores actuales. Este material clástico, generado por bioerosión, podría haber constituido una fuente importante de aprovisionamiento de los sedimentos, luego redepositados en áreas de canal, planicies de inundación, o interfluvios, sin necesidad de apelar a procesos naturales abióticos. Así también, considerando la abundante y variada fauna del Cenozoico tardío y las numerosas evidencias de rastros de actividad, es muy probable que numerosas estructuras sedimentarias en forma de lente, o cubetas irregulares, de dimensiones inferiores a los 2 metros, generalmente interpretadas como rasgos erosivos fluviales, representen cuevas colapsadas o truncadas.
CONCLUSIONES
La interpretación de las estructuras biogénicas atribuidas a mamíferos grandes debe efectuarse sobre la base de criterios independientes y en relación contextual, lo que ha permitido incluir a los perezosos de la familia Mylodontidae en la lista de probables constructores de las cuevas de grandes dimensiones del Cenozoico tardío pampeano. Si bien por el momento no es posible identificar materiales generados por la fauna y estimar su participación en los registros sedimentarios, es útil introducir en nuestros modelos y reconstrucciones paleoambientales la acción de la fauna como agente geomorfológico. Las especulaciones y cuestionamientos que hemos planteado reflejan por un lado el desconocimiento que todavía tenemos del papel desempeñado por la megafauna en los sistemas ecológicos pleistocenos de la región pampeana. Por otro lado, ponen de relieve la necesidad de considerar, en las interpretaciones sedimentológicas y estratigráficas la actividad de la fauna como un agente geomorfológico significativo en la evolución del paisaje, lo que supone profundizar la interacción de geólogos y paleontólogos para el logro de objetivos comunes.
Agradecimientos
Los autores desean expresar su agradecimiento al Dr. E. Tonni, por la lectura crítica del manuscrito, al Dr. G. De Iuliis por la lectura del Extended Abstract y a uno de los revisores, el Dr. J. Genisse, por las recomendaciones efectuadas. Maruca Riggs nos asistió durante la redacción del manuscrito. Este trabajo fue financiado parcialmente por los proyectos PICT 1961 (Agencia Nacional de Promoción Científica y Tecnológica), Exa 10 de la Universidad Nacional de Mar del Plata y N242 del Programa de Incentivos de la Universidad Nacional de La Plata.
REFERENCIAS BIBLIOGRÁFICAS
1. AMEGHINO, F., 1908. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapadmalal. Museo de Historia Natural de Buenos Aires 7(3): 343-428. [ Links ]
2. BARGO M. S., S. F. VIZCAÍNO, F. M. ARCHUBY & R. E. BLANCO, Enviado. Limb bone proportions, strength and digging in some Lujanian (Late Pleistocene-Early Holocene) mylodontid ground sloths (Mammalia, Xenarthra). Journal of Vertebrate Paleontology. [ Links ]
3. CUENCA ANAYA, J., 1995. El aparato locomotor de los escelidoterios (Edentata, Mammalia) y su paleobiología. Colección “Estudis” 6. Adjuntament de Valencia. 452 pp.
4. EDMUND, G., 1985. The fossil giant armadillos of North America (Pampatheriinae, Xenarthra = Edentata). En G. G. Montgomery (Ed.), The evolution and ecology of armadillos, sloths and vermilinguas: Smithsonian Institution Press: 83-93. Washington D.C. [ Links ]
5. FRENGUELLI, J., 1921. Los terrenos de la costa atlántica en los alrededores de Miramar (Provincia de Buenos Aires) y sus correlaciones. Boletín de la Academia Nacional de Ciencias en Córdoba XXIV: 325-485. [ Links ]
6. FRENGUELLI, J., 1928. Observaciones geológicas en la región costanera sur de la Provincia de Buenos Aires. Universidad Nacional del Litoral, Facultad de Ciencias de la Educación, Anales 3: 101-130. [ Links ]
7. FRENGUELLI, J., 1955. Loess y limos pampeanos. MEN-Universidad Nacional de la Plata, Facultad de Ciencias Naturales y Museo de la Plata. Serie Técnica y Didáctica Nro 7. 88 pp. La Plata. [ Links ]
8. GENISE, J., 1989. Las cuevas de Actenomys (Rodentia: Octodontidae) de la Formación Chapadmalal (Plioceno superior) de Mar del Plata y Miramar (Provincia de Buenos Aires). Ameghiniana 26(1-2): 33-42. [ Links ]
9. HICKMAN, G. C., 1990. Adaptativeness of tunnel system features in subterraneal mammal burrows. En E. Nevo & O. Reig (Eds.), Evolution of subterranean mammals at the organismal and molecular levels. Proceedings of the Fifth International Theriological Congress: 185-210. Roma. [ Links ]
10. IMBELLONE, P. & M. TERUGGI, 1988. Sedimentación crotovínica en secuencias cuaternarias bonaerenses. II Reunión Argentina de Sedimentología, Actas: 125-129. Buenos Aires. [ Links ]
11. IMBELLONE, P., TERUGGI, M. & L. MORMENEO, 1990. Crotovinas en sedimentos cuaternarios del partido de la Plata. En M. Zárate (Ed.) Simposio Internacional sobre Loess. Características, cronología y significado paleoclimático del loess: 166-172. Mar del Plata. [ Links ]
12. KRAGLIEVICH, L., 1934. La antigüedad pliocena de las faunas de Monte Hermoso y Chapadmalal, deducidas de su comparación con las que le precedieron y sucedieron. Imprenta El Siglo Ilustrado, Montevideo: 17‑136. [ Links ]
13. KRAGLIEVICH, L., 1952. El perfil geológico de Chapadmalal y Miramar, Prov. De Buenos Aires. Revista Museo Municipal Ciencias Naturales y Tradicionalista 1(1):8-37. Mar del Plata. [ Links ]
14. OWEN, R., 1842. Description of the skeleton of an extinct gigantic sloth, Mylodon robustus, Owen, with observations on the osteology, natural affinities, and probable habits of the megatherioid quadruped in general. R. & J. E. Taylor, London, 176 pp. [ Links ]
15. OWEN, R., 1851. On the Megatherium (Megatherium americanum, Blumenbach). I. Preliminary observations on the exogenous processes of vertebrae. Philosophical Transactions of the Royal Society of London 141(2): 719-764. [ Links ]
16. PEÑA, M. I., 1997. Tafonomía de restos óseos y condiciones ambientales del Plioceno tardío, Playa San Carlos (Pdo. de Gral Puerredón). Tesis de Grado de Licenciatura en Ciencias Biológicas. Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata. 37 pp. (inédito). [ Links ]
17. QUINTANA, C. A., 1992. Estructura interna de una paleocueva, posiblemente de un Dasypodidae (Mammalia, Edentata) del Pleistoceno de Mar del Plata (Provincia de Buenos Aires, Argentina). Ameghiniana 29(1): 87-91. [ Links ]
18. SCOGNAMILLO, D., 1993. Estructura de las cuevas de Actenomys (Rodentia, Octodontidae) de la Aloformación Playa San Carlos, Plioceno tardío (Barranca de los Lobos, Pdo. de Gral Pueyrredón): significado paleoecológico y estratigráfico. Tesis de Grado de Licenciatura en Ciencias Biológicas. Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata. (inédito). [ Links ]
19. WHITE, J. L., 1997. Locomotor adaptations in Miocene Xenarthrans. En R. F. Kay, R. H. Madden, R. L. Cifelli & J. J. Flynn (Eds.), Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press: 246-264. [ Links ]
20. WINGE, H., 1941. Edentates (Edentata). En S. Jensen, R. Spärck & H. Volsoe (Eds.), The interrelationships of the Mammalia genera: 319-341. Reitzels Forlag, Copenhagen. [ Links ]
21. ZÁRATE, M. A., 1989. Estratigrafía y geología del Cenozoico tardío aflorante en los acantilados marinos comprendidos entre Playa San Carlos y el arroyo Chapadmalal, Partido de General Pueyrredón, provincia de Buenos Aires. Tesis Doctoral. Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata. 220 pp. (inédito). [ Links ]

