










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Asociación Argentina de Sedimentología
Print version ISSN 1853-6360
Rev. Asoc. Argent. Sedimentol. vol.10 no.2 La Plata Aug./Dec. 2003
ARTÍCULOS
Integración de facies y biofacies en un análisis secuencial en plataformas clásticas Devónicas del sur de Bolivia y noroeste Argentino
Luis A. Alvarez1, Alejandra Dalenz-Farjat1, Roberto M. Hernández1 y Luis Albariño2
1XR srl. Exploracionistas Regionales. Parque Belgrano, Primera Etapa, Manzana N Casa 14,4400 Salta.
2Repsol YPF. Esmeralda 255, Piso 10, Of. 1001. C1035ABE Buenos Aires.
luis@xrexplorer.com , alejandra@xrexplorer.com , hernandezr@xrexplorer.com, lmalbarinom@repsolypf.com
Recibido: 11 de agosto de 2003
Aceptado: 02 de diciembre de 2003
Resumen: Se exponen los resultados obtenidos del análisis de facies, asociaciones de facies, ambientes y estudio de biofacies en la cuenca silúrico-devónica del subandino sur de Bolivia y del noroeste de Argentina, analizada en su tramo devónico. Se expone la bioestratigrafía utilizada en esta cuenca, tanto en base a palinomorfos como fauna bentónica. Se caracterizan los ambientes y subambientes en una plataforma silicoclástica marina desarrollados en 19 Secuencias Genéticas, entre el Lochkoviano y el Frasniano. Se plantea un modelo de distribución de facies y de ambientes. Finalmente se estudian las biofacies desarrolladas durante etapas de contracción y de expansión de esta cuenca.
Palabras Claves: Asociación de Facies; Ambientes Silicoclásticos; Biofacies; Devónico; Argentina; Bolivia.
EXTENDED ABSTRACT
Integration of facies and biofacies in a sequence analysis in detrital platforms from Devonian of southern Bolivia and northwestern Argentina
A silurian-devonian basin of southern Bolivia and northwestern Argentina (Figs. 1, 2) is interpreted as a backarc basin developed in a noncontracted margin, like a Java system (Fig. 3). In this basin, 19 Genetical Sequences are described in a continuous sedimentation from Ludlowian to Frasnian (Fig. 8). In this paper we analize the Devonian interval. Data from facies analysis, facies associations, environments and biofacies of this basin are presented (Figs. 4, 5).
The biostratigraphy of the basin is based in palynomorphs and benthic fauna. Palynological analysis is based in world wide associations and First Occurrence Biozone for each association. Chitinozoans, acritarchs, spores, and others are considered to determine the age of each sample. Benthic fauna are divided in two types, specialized and non-specialized. The former is considered an important fauna to date. This fauna is calibrated with other basins outside Gondwana (Table 1).
Facies and environmental conclusions are placed in an eustatic curve designed for this period. 21 facies associations are described and exposed (Figs. 4, 6, 7). Environmental determinations in a marine siliciclastic platform developed in 19 Genetical Sequences from Lochkovian to Frasnian are characterized (Fig. 8). Seven marine environments are described and placed in a model where facies associations and environments present a logical distribution (Fig. 5).
Biofacies are studied during contracted and expanded periods. During contractions, in normal regressions of Pragian and Emsian age, biofacies are more diversified and frequent than biofacies developed in the same conditions during Eifelian expansions of the basin (Fig. 10). During forced regressions, biofacies are less diversified and poorly distributed in space (Fig. 11).
In this platform type, development of benthic fauna is a function of available space and eustatic changes. During a second order eustatic fall, a forzed regression could be determinated in the development of the benthic fauna.
During expanded periods, biofacies developed in normal regressions during Eifelian and Giventian are less diversified and less frequent than biofacies developed during similar conditions in the Coger Devonian (Fig. 12). This process is controlled by boreal benthic fauna until the Eifelian. We propose a biofacies distribution model for this siliciclastic ramp (Fig. 13 and Table 2).
Key Words: Facies association; Siliciclastic Environments; Biofacies; Devonian; Argentina; Bolivia.
INTRODUCCION
Se presentan las asociaciones de facies sedimentarias desarrolladas durante la depositación de los cortejos sedimentarios como consecuencia de las variaciones eustáticas, con o sin influencia tectónica, durante el Silúrico superior a Devónico superior (Ludlowiano - Frasniano) en el noroeste de Argentina y en el sur de Bolivia (Fig. 1). Esta disposición de facies y ambientes se enmarca en líneas-tiempo que son definidas mediante la paleontología y la palinología. Se estudiaron 17 perfiles de superficie y 6 perfiles eléctricos de pozos (Fig. 2), donde se definieron líneas-tiempo. El reconocimiento de Secuencias Estratigráficas Genéticas en el sentido de Galloway (1989), la definición de facies, procesos y ambientes sedimentarios ubicados dentro de las líneas-tiempo mencionadas, permitió establecer un modelo de cuenca sedimentaria (Albariño et al., 2002). En el presente trabajo, se presenta el detalle de las asociaciones de facies, ambientes definidos en una rampa silicoclástica y las biofacies que se desarrollaron durante las etapas de contracción y de expansión que sufrió esta cuenca durante cerca de 57 millones de años.
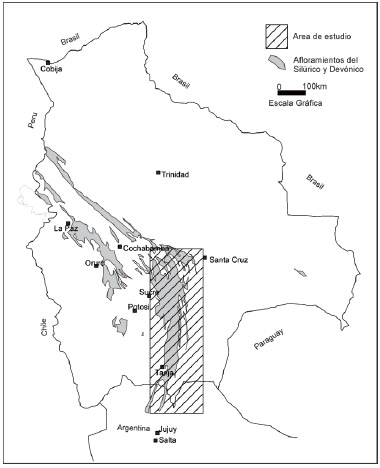
Figura 1: Mapa de afloramientos Siluro-Devónicos del sur boliviano y noroeste argentino. A partir del mapa geológico de Bolivia publicado y mapas del noroeste argentino de XR s.r.l.
Figure 1: Map of Silurian and Devonian outcrops of Bolivia and NW Argentina. From published maps. Information from NW Argentina is from XR. s.r.l.

Figura 2: Mapa de ubicación de las columnas levantadas en este trabajo y otras localidades y pozos mencionados en Bolivia y Noroeste Argentino.
Figure 2: Map of stratigraphic columns made by the authors and other places and wells referred in Bolivia and Northwestern Argentina.
METODOLOGIA
Se identificaron 21 asociaciones de facies en el campo, y se definieron ambientes sedimentarios marinos. Este análisis fue enmarcado en líneas tiempo. Las observaciones fueron integradas en correlaciones este - oeste y norte - sur tanto con datos de superficie como de subsuelo, lo que permitió definir eventos regresivos en la cuenca, tanto en condiciones normales como forzadas, y eventos expansivos. Se definieron 19 Secuencias Genéticas y se estudiaron sus biofacies. En el análisis paleoecológico se determinaron algunos conceptos claves:
En este trabajo se considera el concepto de biofacies según Reading (1986) y Arche (1992), como una facies o cuerpo de roca con características específicas, basadas en el color, estratificación, composición, textura, contenido fosilífero y tipo de estructuras sedimentarias y donde priman las condiciones biológicas de la fauna. Cada biofacies fue caracterizada con los siguientes parámetros paleoecológicos:
Se estudiaron las biofacies para definir el tipo de comunidad o asociación. Se denominan comunidades autóctonas aquellas que no sufrieron ningún desgaste, ni transporte y que mantienen rasgos en posiciones de vida. Las asociaciones parautóctonas son aquellas que habrían sufrido remoción por efecto de las corrientes hidrodinámicas pero sin sufrir transporte ni selección.
Las asociaciones alóctonas son aquellas que fueron acumuladas hidrodinámicamente y cuyas relaciones y posiciones de la fauna dentro de la asociación son casuales.
Se hizo el análisis de las biofacies, calculando la diversidad, es decir la medida cuantitativa adimensional de la existencia de un taxón a nivel de familias, géneros y especies, en una comunidad o en una asociación dada. Esta medida se calcula en función al desarrollo matemático de Shannon-Weaver empleada por Sanders (1968), Sanchez (1984), Sanchez et al. (1991) y Dalenz-Farjat (2000). Los resultados de estos cálculos se ubican en valores que fueron analizados previamente para la cuenca silúrica-devónica de Bolivia, comparando con diversidades de otras cuencas (Dalenz-Farjat, 2000) a fin de establecer parámetros cualitativos de los valores calculados. Se definieron las composiciones de las biofacies en base a porcentajes de representación de un taxón dentro de la totalidad de individuos y especies de una comunidad y asociación. Se definieron los tipos tróficos de la fauna constituyente de cada biofacies en base a las clasificaciones de Walker y Bambach (1974) y Bambach (1983).
CARACTERIZACION DE LA CUENCA
Si bien es controvertido aún definir el marco tectónico que dio origen a la cuenca, las evidencias registradas hasta la fecha para el sector central, sur y sudoeste, resumidas en Dalenz et al. (2002), indican para el margen central del Gondwana, un borde occidental rígido y sobreelevado a partir de una etapa de convergencia o proto-subducción que se activa notablemente en el Cámbrico superior - Ordovícico inferior. El cual es conocido como evento D2 en Sierras Pampeanas (Rapela et al., 1992) y Fase Irúyica (Salfity et al., 1984) en Cordillera Oriental, dando inicio al magmatismo Famatiniano (Aceñolaza & Toselli, 1973), que continúa durante el Ordovícico hasta el Devónico superior con las fases Oclóyica y Chánica respectivamente (Salfity et al., 1984). En la Puna, el magmatismo Famatiniano se desarrolla a lo largo de dos fajas submeridianas, evidenciando ambas en su etapa temprana filiación oceánica, con características de dorsal oceánica y relacionadas a zonas de subducción con la generación de arcos volcánicos (Rapela et al., 1992). Las características petrológicas, geoquímicas e isotópicas de los plutones (Grupo G2 y post D2, Fig. 3 de Rapela et al., 1992) indican un sensible aumento del componente cortical en la fuente y marcada semejanza con granitoides colisionales. Ello sustenta diversos modelos de colisión continente-continente (Coira et al., 1982; Ramos et al., 1986; Ramos, 1988; Dalla Salda et al., 1990 en Rapela et al., 1992) o arco de isla-continente para el intervalo Ordovícico-Devónico superior.

Figura 3: Sistemas Arco-Fosas en margenes continentales, según Dickinson y Seely (1979). Notar que la cuenca de retroarco puede formarse tanto en márgenes con acortamiento tipo"andino" como sin acortamiento, tipo "Java".
Figure 3: Continental-Margin Arc-Trench Systems, after Dickinson and Seely (1979). A backarc basin can be deve-lopped in a contracted margin, as an "andean" system and noncontracted margin, as a "Java" system.
Para esta parte occidental del Gondwana durante el Silúrico-Devónico, se sostiene como hipótesis una cuenca de perfil este - oeste asimétrico, que se ha desarrollado sobre un basamento cratonizado (cuenca de retroarco sin acortamiento ver Dalenz et al. (2002) y Selley (1998), capítulo 8 y pg. 385), (Fig. 3) constituido por el cratón de Guaporé, el macizo de Arequipa-Huarina, los terrenos Chiquitanos, los cratones Pampeano y Río de la Plata y la acreción de terrenos dislocados de Laurentia (Precordillera), en tiempos del Paleozoico temprano (Suárez Soruco, 1999; Suárez Soruco et al., 2000). Presenta un borde oriental de bajo relieve con escaso aporte de sedimentos y una subsidencia muy lenta debido a la rigidez de la corteza con un espesor de 85 km en el este de Bolivia y más de 50 km al este de Argentina (Stewart y Watts, 1997) y un borde occidental con alto relieve, de cuyas áreas proviene el mayor volumen de sedimentos depositados en la cuenca, con una subsidencia más acentuada debido probablemente a un menor espesor cortical. La escasa a nula influencia tectónica sinsedimentaria estaría demostrada por la ausencia de una faja plegada intra Silúrico superior-Devónico, y por el modelo estratigráfico de cuenca propuesto en base a las observaciones de campo y sísmicas (Albariño et al., 2002), lo que permite poner en discusión el modelo tectogenético de antepaís (Dalenz et al., 2002).
BIOESTRATIGRAFIA
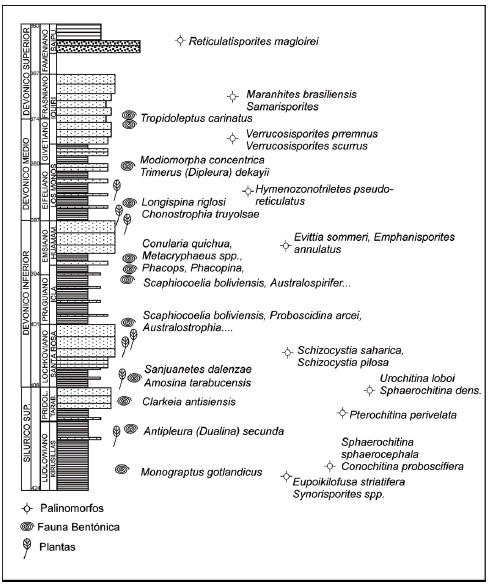
La bioestratigrafía que se desarrolló en esta cuenca se basa en asociaciones de fauna y flora calibradas en la cuenca silúrico-devónica bolivianaargentina en los sectores interandino, andino y subandino. Las asociaciones empleadas en la bioestratigrafía de esta cuenca están constituidas tanto por esporas, quitinozoarios, acritarcas, como por fauna bentónica boreal. Este trabajo retoma las conclusiones bioestratigráficas a las que arribaron numerosos especialistas tanto de empresas petroleras estatales como YPFB e YPF, como de privadas, después de décadas de investigación. En este trabajo se presenta una apretada síntesis de esas ideas que fueron publicadas en Albariño et al. (2002) adicionando grupos de mayor resolución bioestratigráfica (Tabla 1). A través de la palinología, la correlación cronológica de las series estudiadas en Bolivia y regiones adyacentes fue realizada tomando en cuenta el registro de palinomorfos comunes y especies claves a nivel mundial entre las Provincias del Continente de las Viejas Areniscas Rojas, la Región Ardeno-Renana, Gondwana, hasta las actuales China, Rusia, Australia y la Antártida, fundamentalmente para las esporas del Silúrico y Devónico, según Richardson y McGregor (1986); Streel et al. (1987) y Albariño et al. (2002). Asimismo se reconocieron eventos morfológicos importantes en la sucesión de esporas a nivel mundial y "primeras apariciones" de ciertas taxa reflejando la evolución aparentemente sincrónica de los mismos (Albariño et al., 2002). A través de la paleontología de invertebrados, las dataciones empleadas en la calibración del Devónico tipo (en cuencas no gondwánicas) se determinan sobre la base de la presencia de conodontes, graptolites y amonites, que a su vez se encuentran calibrados por dataciones radiométricas de 2 niveles volcánicos en Europa. En la cuenca silúrica-devónica de Bolivia y del noroeste Argentino no hay conodontes, ni graptolites y son muy escasos los amonites (goniatites).
Tabla 1: Columna generalizada del Siluro-Devónico en el area de Icla - Tarabuco, con controles de edades en base a asociaciones palinológicas y paleontológicas válidas para el area de trabajo.
Table 1: Generalized column of Silurian - Devonian from Icla - Tarabuco area, with palynozones and paleontological associations in the studied area.
Fauna No Especializada: El Devónico tipo a nivel mundial tanto en Alemania, República Checa, Francia y Bélgica se desarrolló en paleolatitudes muy bajas, en aguas templadas, en facies calcáreas. Mientras que en Gondwana las facies depositadas son silicoclásticas, la fauna desarrollada en estos mares es de temperaturas frías y de latitudes altas a muy altas. En este panorama, la fauna bentónica del hemisferio sur se caracteriza por ser de largos biocrones, aguas frías y por lo tanto es poco diversificada y poco especializada, es decir de hábitos oportunistas. Las anteriores condiciones expuestas hacen que esta fauna sea dominante en la cuenca y se asocie entre sí siguiendo controles ambientales (biofacies), cortando líneas tiempo. Por lo tanto, esta clase de asociaciones no es propicia para utilizarla en dataciones.
Fauna Especializada: Se define así a aquellos grupos de fauna que generaron hábitos muy específicos, por lo cual se desarrollaron muy rápidamente en determinadas áreas pero que, por ser tan especializadas, fueron muy frágiles a cualquier cambio tanto climático y geográfico, como genético, sufriendo la extinción de cada especie de manera violenta. Estos grupos muy especializados sufrieron severas extinciones a nivel específico y genérico, condiciones ideales para caracterizar fauna de biocrones cortos.
En consecuencia, la datación en las secuencias estudiadas se realiza a través de la fauna bentónica no especializada a la que se adiciona por un lado fauna boreal especializada que invade la cuenca gondwánica desde provincias norteñas o del margen norte del Gondwana en eventos transgresivos relativamente sincrónicos, y por otro lado, fauna especializada que se desarrolló en el Gondwana durante las progradaciones de la cuenca. Se establece una tabla cronoestratigráfica generalizada, con la cual se propusieron las líneastiempo clave entre el Ludlowiano y el Frasniano en esta parte del Gondwana, en base a quitinozoos, graptolites, esporas y fauna bentónica previamente calibrada (Tabla 1).
DESCRIPCIÓN DE FACIES
Características de las asociaciones de facies siluro-devónicas en el área de estudio
Se dan a conocer 21 asociaciones de facies con las cuales se trabajó esta parte de la cuenca Silúrico- Devónica (Figs. 4 y 5). Además se presentan las facies que fueron originalmente definidas por Starck et al., 1992 y adaptadas al proyecto.

Figura 4: Resumen de conjunto de asociaciones de facies y subambientes sedimentarios estudiados en la Cuenca Siluro Devónica del Subandino del sur de Bolivia y del noroeste Argentino. Se incluyen las asociaciones de facies continentales y mixtas, modificado de Starck (1995).
Figure 4: Facies associations and sedimentary environments studied in Silurian - Devonian basin in Southern Bolivia and Northwestern Argentina. Continental and mixed continental facies are included, modified from Starck (1995).

Figura 5: Conjunto de asociaciones de facies devónicas en la plataforma marina (según Elliott (1986).
Figure 5: Devonian facies associations in marine platform, from Elliott (1986).
Asociación de facies D1: Pelitas laminadas.
Pelitas negras micáceas, desde arcilitas a fangolitas muy finas, laminadas con algunos nódulos de hierro. La laminación paralela se encuentra tanto en la base de los bancos como en los techos. Eventualmente se encuentran algunos bancos de arenas muy finas grises, intercaladas con la facies pelítica. Estos bancos de arenas suelen tener pocos centímetros de espesor y pueden presentar internamente laminación ondulítica y pequeños hummockies. Esta asociación presenta en todos los casos un predominio de material pelítico sobre el arenoso, con proporciones que varían de 80 % a 20 % y de 90 % a 10 %.
Interpretación: Esta asociación de facies se depositó en la parte distal de la plataforma interna, donde la sedimentación se produce casi exclusivamente por decantación. Esta asociación de facies corresponde a los Cortejos de Nivel Alto (HST) y Transgresivo (TST) en el interior de la cuenca. La energía hidrodinámica en esta parte de la cuenca es mucho menor, con lo cual los estratos son de granulometría muy fina y laminados. Se registran eventuales tormentas que depositaron delgados niveles de arena muy fina intercalados. La sedimentación se produce por debajo del nivel de base del oleaje, inclusive en tiempo de tormentas.
Asociación de facies D2: Intercalación de pelitas laminadas y arenas muy finas.
Intercalación de pelitas negras con laminación paralela y delgados bancos de arena muy fina gris con acción de olas en el tope de los bancos de arena y desarrollo interno de estratificación tipo hummocky. En la fracción fina suelen encontrarse nódulos de hie-rro. En esta asociación se tendría una relación de arenas y pelitas de 50 % de pelitas laminadas, 25 % de arenas con acción de olas y 25 % restante de arenas con hummockies. En algunos bancos de arenas muy finas intercalados, se encontraron pequeños deslizamientos sinsedimentarios. Esta característica es muy poco frecuente. El espesor máximo de los bancos de arena puede llegar a los 80 cm.
Interpretación: Esta asociación de facies corresponde a una sedimentación de plataforma interior, algo más somera que D1, pero igualmente relacionada con una energía hidrodinámica muy reducida, asociada a tormentas esporádicas que removilizaron material arenoso al interior de la cuenca. Acción de ola y estratificación de tipo hummocky determina una mayor energía hidrodinámica en la plataforma, en comparación con la asociación D1. Durante la depositación de esta asociación de facies, se registran eventos en donde la acción de olas ha afectado los depósitos de interior de plataforma en los períodos de somerización. La asociacion de facies D2a corresponde a la intercalación de pelitas laminadas y arenas muy finas laminadas.
Asociación de facies D3: Intercalación de limolitas laminadas y arenas muy finas con hummockies.
Asociación de facies que se caracteriza por ser grano y estrato creciente, desde limolitas laminadas negras a grises oscuras que pasan a arenas muy finas a finas con acción de olas y estratificación cruzada de tipo hummocky. Las arenas finas cuspidales de cada parasecuencia finalizan con hummockies y también con acción de olas en el techo de cada banco. Esta asociación de facies se compone de una proporción limolitas-arenas del 50 %.
Interpretación: Esta asociación corresponde a una sedimentación de plataforma interior proximal a frente de costa distal, donde aún se mantienen constantes procesos de decantación de material limoso, pero que se ven interrumpidos por tormentas que removilizan el sedimento, con aportes de arenas muy finas. Esta sedimentación se ubicaría por encima del nivel de base del oleaje durante las tormentas. Durante las tormentas, el material es sometido a cierto flujo de energía hidrodinámica con lo cual se observan depósitos con acción de olas y hummockies.
Las asociaciones de facies D3a y D3b corresponden a variantes de arenas muy finas laminadas y arenas muy finas vaques, respectivamente.
Asociación de facies D4: Vaques fangolíticas a limolíticas con arenas muy finas con hummockies.
Esta es una asociación grano y estrato creciente que se inicia en la base con vaques masivas intensamente bioturbadas, de matrix tanto fangolítica como limolítica. Las vaques varían de grises a gris oscuras de acuerdo al porcentaje de pelitas en la matrix. En el tope de las vaques, se intercalan arenas con estratificación entrecruzada tipo hummocky y con acción de olas. Igualmente hacia el tope, las arenas pasan a ser finas y con laminación paralela, muy micáceas y grises.
Interpretación: Esta asociación de facies corresponde a una sedimentación en el frente de costa, probablemente en el frente de costa distal. En períodos de buen tiempo, la fauna epibentónica produce una fuerte bioturbación del material sedimentado tanto al desplazarse como al alimentarse. Durante los períodos de tormentas, se produce un aporte de material arenoso de granulometría muy fina a fina con un proceso de sedimentación más rápida. En el siguiente período de buen tiempo, nuevamente la fauna retrabaja el material, mezclando tanto la fracción pelítica autóctona y depositada principalmente por decantación, con el material psamítico transportado por las tormentas. El resultado es una mezcla de fracciones tanto pelíticas como arenosas, definidas como vaques de matrix fangolítica a limolítica. La energía hidrodinámica aumenta en esta asociación pues pasa de vaques bioturbadas a arenas con predominio de acción de olas, hummockies y culmina en arenas laminadas.
Las asociaciones de facies D4a y D4b corresponden a variantes de vaques limolíticas a arenas muy finas tabulares con hummockies (D4a) y vaques limolíticas y arenas muy finas con laminación paralela (D4b).
Asociación de facies D5: Arenas finas con hummockies y arenas tabulares bioturbadas.
Asociación de facies que se caracteriza por arenas muy finas a finas, castañas, muy micáceas, en bancos de 20 a 30 cm de espesor, de bases netas, con estratificación tipo hummocky, pasando a arenas tabulares que pueden presentar cierta laminación en la base y que pasan a ser masivas y muy bioturbadas en el techo en espesores que alcanzan el metro. Estas arenas presentan muy buena selección y son arenas más limpias que aquellas descritas en las asociaciones D4, D4a y D4b.
Interpretación: Esta asociación de facies se depositó en el frente de costa, dominado por olas y tormentas episódicas. La depositación de esta asociación se ubica entre el nivel de base del oleaje durante buen tiempo y el nivel de base del oleaje durante mal tiempo. Las arenas muy finas a finas son depositadas durante buen tiempo y son retrabajadas durante las tormentas, con hummockies y laminaciones. El proceso pierde energía hacia el techo cuando se desarrollan bioturbaciones. La asociación de facies D5a corresponde a arenas finas laminadas y arenas tabulares bioturbadas.
Asociación de facies D6: Tempestitas.
Se caracteriza por delgados bancos tabulares a lenticulares de material psamítico, de arenas finas hasta medias, con clastos pelíticos y de arenas muy finas. Internamente estos bancos delgados pueden presentar nódulos de hierro retrabajados junto con clastos blandos y las arenas más finas. Estos bancos se encuentran en el techo de las asociaciones D3 y D4. En algunos casos, pueden encontrarse algunos fragmentos de fauna bentónica incorporada a la tempestita como un clasto más que sufrió transporte en el proceso de formación.
Interpretación: Esta asociación se forma en el frente de costa donde se depositan niveles de material transportados por las tormentas que erodan las superficies de litología más fina, transportando clastos de pelitas de la playa frontal al frente de costa a frente de costa distal. Estos niveles de tempestitas se encuentran depositados sobre varias asociaciones de facies, pues responden a eventos episódicos como son las tormentas que retrabajan material en la playa frontal hacia el frente de costa y eventualmente hacia la plataforma interna proximal.
Las asociaciones de facies D6a y D6b corresponden a variantes de arenas finas con slumps y tempestitas (D6a) y arenas finas laminadas con tempestitas (D6b).
Asociación de facies D7: Arenas con estratificación entrecruzada de bajo ángulo y arenas con desplazamientos.
Esta asociación está constituida por arenas muy finas a finas micáceas, grises, con estratificación entrecruzada de bajo ángulo y una eventual acción de olas, en espesores de medio metro como máximo, de bases netas y topes algo ondulosos. Por encima de estas arenas se encuentran arenas finas en bancos de más de un metro a metro y medio de espesor, con desplazamientos sinsedimentarios, donde la granulometría involucrada es de muy fina a fina.
Interpretación: Esta asociación corresponde a una depositación en el frente de costa en el nivel de batido de las olas, donde se produce mejor selección mineralógica en las arenas, con estratificación entrecruzada de bajo ángulo. La sedimentación se produce en un nivel de mayor energía hidrodinámica con respecto a las asociaciones D1, D2, D3, D4 y D5. Las asociaciones de facies D7a y D7b corresponden a variantes de arenas laminadas que culminan en arenas con deslizamientos (D7a) y arenas con estratificación entrecruzada de bajo ángulo (D7b).
sociación de facies D8: Arenas finas laminadas y arenas con estratificación entrecruzada de bajo ángulo.
Arenas finas a finas medias, en bancos tabulares, cuarcíticas, con laminación paralela alcanzando el metro y medio de espesor. La asociación de facies culmina con bancos con estratificación entrecruzada de bajo ángulo. Eventualmente se registraron arenas de grano medio laminadas en el techo de la asociación.
Interpretación: Esta asociación de facies se produce en la playa frontal, donde el proceso principal es el batido del oleaje, que produce la laminación paralela de alto régimen de flujo en el frente de la costa. Este nivel de energía hidrodinámica produce una buena selección mineralógica de las arenas, por lo cual se encuentran muy bien seleccionadas, son predominantemente cuarcíticas y con una distribución micácea laminada.
Las asociaciones de facies D8a, D8b y D8c corresponden a variantes de arenas finas laminadas tabulares (D8a), arenas tabulares medianas laminadas (D8b) y arenas tabulares bioturbadas y con estratificación entrecruzada (D8c).
Se adaptaron las asociaciones de facies para el borde sur y oeste de la cuenca definidas por Starck (1995):
Facies P2: Pelitas grises y verdosas, hasta rojizas. Menos laminadas que las anteriores, pueden contener detritos vegetales.
Facies A2: Areniscas finas a medianas, en tonalidades grises. Con laminación ondulítica dada por ondulitas de corriente.
Facies A4: Areniscas finas a medianas, gris verdosas, en bancos tabulares a lenticulares. Con entrecruzamiento tangencial (sigmoides?). Generalmente en bancos amalgamados.
Facies A5: Areniscas gruesas hasta conglomerádicas. Con clastos dispersos de hasta 2 cm, con tendencia a concentrarse en los techos de las capas (por aventamiento). Colores amarillentos hasta rojizos. Capas amalgamadas con bases erosivas. Entrecruzamientos planares y en artesas.
Facies C1: Conglomerado fino, arenoso. Gris amarillento hasta rojizo. Oligomíctico, con clastos bien redondeados de cuarzo y cuarcitas de 1cm de diámetro promedio. Bancos amalgamados con estructura interna entrecruzada en artesa (Fig. 4).
AMBIENTES SEDIMENTARIOS
Del análisis de este conjunto de asociaciones de facies se ha determinado que el medio sedimentario en el que se depositaron las sedimentitas devónicas del sur de Bolivia y del noroeste de Argentina corresponde a un ambiente marino de playa y de plataforma continental, con dominio de acción de olas y episodios de tormentas con menor influencia de mareas. La combinación de estas asociaciones de facies permitió el reconocimiento de procesos sedimentarios y definir subambientes, los que han sido diferenciados de la siguiente manera (Fig. 5):
Subambiente Continental y Mixto: Este subambiente se ha desarrollado en una planicie aluvial y está constituido por una asociación de facies integrada por facies P2, A2, A4, A5 y C1. Ha sido descripto en borde de cuenca, registrado entre el Lochkoviano y el Emsiano en los perfiles de Abra de Zenta, Alarache y Lipeo y en el Pragiano del perfil de Sella.
Ambiente Sedimentario de Playa: con el subambiente de Playa Frontal, Frente de Costa Proximal y Frente de Costa Distal.
Playa Frontal: los límites de depositación de este subambiente lo constituyen los límites de marea alta y baja o playa media. Los materiales que se depositan en esta zona son arenosos y las proporciones de pelitas son pequeñas. Las capas presentan laminación cruzada planar de bajo ángulo. Estas buzan en dirección al mar y representan verdaderas superficies de acreción. Es un área de acumulación de valvas (conchillas) muertas y parcialmente bioturbado. En la zona de trabajo se ha reconocido en muy escasos lugares definido por las asociaciones de facies D7, D8a y D8b.
Frente de Costa: Este subambiente permanece siempre bajo el agua; se halla delimitado entre el límite infe-rior de marea baja y el punto donde el oleaje deja de ejercer su acción sobre el fondo, en periodos de buen tiempo. Si bien dominan las facies arenosas también participan las pelitas. Son comunes las ondulitas de olas alineadas paralelas a la costa de distinta escala y por incluir abundante fauna, son intensamente bioturbados. Se ha subdividido en: Proximal, dominio de psamitas con ondulitas, definido por asociaciones de facies: D5, D5a, D7, D7a, D7b, subordinada D4 y D4a y Distal, dominio de pelitas bioturbadas, definido por asociaciones de facies: D3, D3a, D3b, D4, D4a, D4b, y muy subordinadas D2 y D5. Este subambiente, en períodos de mar agitado, podría incluir asociaciones de facies D6, D6a y D6b (Tempestitas).
Ambiente Sedimentario de Plataforma Continental: con el subambiente de la Zona de Transición y Plataforma Interna.
Zona de Transición: Los límites de depositación de este subambiente no son muy precisos; se podría tomar como referencia del límite superior, al punto donde el oleaje deja de ejercer su acción en el fondo en períodos de buen tiempo, y el límite inferior hasta donde se registran eventos (acción de oleaje en el fondo) producidos con mar agitado y en tiempos de tormentas. Dominan las facies pelíticas, macizas y laminadas, sobre las psamíticas, con bioturbaciones. Estas últimas suelen ser finas a medianas con ondulitas simétricas y asimétricas. Se han reconocido las asociaciones de facies: D2, D2a, D3, D3a, subordinadas, D1, D4, D4a y en períodos de tormentas con D6, D6a y D6b.
Plataforma Interna: Este es un subambiente de interior de cuenca, desarrollado dentro de la plataforma continental. Dominan las facies pelíticas laminadas con procesos de decantación, suelen intercalar niveles ferruginosos con concreciones de óxido de hierro o sulfuros de hierro (pirita), escasamente bioturbadas. Intercalan delgados niveles lenticulares de arenas muy finas macizas a laminadas. Presenta asociaciones de facies D1, D1a y subordinada D2. Con episodios de mal tiempo (tormentas) pueden incluir escasos lentes arenosos con asociación de facies D6 y D6a.
MODELO DE DISTRIBUCION DE FACIES EN LA CUENCA SILURICO - DEVONICA
A partir de las observaciones estratigráficas y bioestratigráficas de campo en las columnas estudiadas, se han interpretado ciclos grano y estrato crecientes más o menos regulares. La mayor dificultad fue definir límites de cortejos sedimentarios, principalmente entre el HST (progradación de nivel alto) y el LST o SMST (nivel bajo). En cambio fue posible observar intervalos progradantes con grandes espesores a partir de la máxima inundación y asignados a "eventos regresivos", sobre el que yacen intervalos de poco espesor (en general grano decrecientes) definidos como "eventos transgresivos".
Para ejemplificar el modelo se han elegido dos etapas representativas en la evolución de la cuenca. Una de contracción durante el Pragiano (SG V, Fig. 8) con una distribución de facies de planicie aluvial y costera desplazadas hacia el interior de la cuenca (Fig. 6). La otra es de expansión en el Eifeliano (SG XII) que genera una inundación regional y el consiguiente desplazamiento de facies de interior de cuenca (plataforma interna) hacia el continente (Fig. 7). Los puntos de control elegidos son: el perfil de Alarache (frontera Bolivia - Argentina) ubicado en una posición proximal en el sector sudoeste de la cuenca y el perfil de Sobo-Sobo (Bolivia) situado a más de 300 km en dirección nor-noreste en cercanías al centro de la cuenca (Fig. 2).

Figura 6: Modelo de distribución de facies. Etapa de Contracción.
Figure 6: Facies distribution model. Contraction period

Figura 7: Modelo de distribución de facies. Etapa de Expansión.
Figure 7: Facies distribution model. Expansion period.

Figura 8: Curvas eustáticas de 2° y 3° orden adaptadas de Johnson et al. (1985) para el Siluro-Devónico del Subandino sur de Bolivia y del Noroeste Argentino. Los números romanos representan las 19 Secuencias Genéticas definidas en la cuenca.
Figure 8: 2d. and 3th. order eustatic curves from Johnson et al. (1985) referred to the Silurian-Devonian of Bolivian and Northwestern Argentinian Subandean. Roman numbers are 19 Genetic Sequences defined in the basin.
Durante la etapa de contracción, el desplazamiento de la línea de costa hacia el interior de la cuenca ("evento regresivo") está evidenciado por la progradación del conjunto de facies continental-mixta (C-M) en la misma dirección, invadiendo varios kilómetros dentro de la planicie costera. Esto genera el desplazamiento y distribución (principalmente por acción de olas y corrientes marinas en la planicie costera) de las arenas constituidas por el conjunto de facies de frente de costa proximal y distal (SFP y SFD) hacia el interior de la plataforma. El conjunto de facies pelíticas representativas de la plataforma interna (OFS) está restringido a las áreas de depocentros de la cuenca, como se puede observar en la Secuencia Genética V de la figura 6. En la etapa de expansión de la cuenca, como la ocurrida durante el Eifeliano, se produce una inundación generalizada, desplazando la línea de costa hacia el continente. Esto se manifiesta en la Secuencia Genética XII con una distribución del conjunto de facies de offshore que alcanza en su máxima inundación las proximidades del perfil de Alarache; desarrollando durante el evento regresivo (progradación de nivel alto) facies de frente de costa distal en su mayor parte y de frente de costa proximal antes del inicio del evento transgresivo con el que termina el ciclo de esta secuencia (Fig. 7).
CICLICIDAD Y CURVA EUSTATICA EN LA CUENCA
De las observaciones realizadas se interpreta que la cuenca se inicia con una etapa de expansión (máximo nivel de base) en el Silúrico superior (Ludlowiano - Pridoliano), con escasos registros de campo en el área estudiada, a la que le sigue una de contracción (mínimo nivel de base) que se extiende durante el Devónico inferior (Lochkoviano - Emsiano) y coincide con un periodo de tiempo en el que se registran los mayores espesores y volúmenes de arenas distribuidos tanto en la zona litoral como en el interior de la cuenca. Posteriormente, una nueva expansión genera un dominio de depósitos pelíticos en el Devónico medio que se extiende desde el Eifeliano hasta el Givetiano superior, a partir del cual se observa nuevamente una progradación de facies arenosas hacia el interior de la cuenca coincidente con otra disminución del nivel de base, que se extiende hasta el Frasniano superior (Devónico superior basal).
Esta ciclicidad coincide en general, con la curva eustática de 2do. Orden empleada por Johnson et al. (1985) para el Paleozoico (Fig. 8). De esta curva se deduce que la mayoría de los reservorios más productivos se encuentran en el Lochkoviano, Pragiano y Emsiano (Devónico inferior) bajo la nominación formal de Santa Rosa y Huamampampa en el primer descenso de nivel de base (curva eustática de segundo orden); y en el Givetiano superior a Frasniano (Devónico medio a superior) como Formación Iquiri durante la segunda etapa de contracción de la cuenca. En las zonas de borde el diacronismo de las unidades formales es más acentuado aun, debido al dominio de facies arenosas proximales depositadas a través del tiempo (Fig. 9). Si bien hay que tener en cuenta las anomalías producidas en distintos sitios de la cuenca, como por ejemplo en los Yacimientos Camiri-Guayruy -Tucán (cuenca devónica de Bolivia), donde se han descripto 14 niveles arenosos petrolíferos de edad givetiana y las "Areniscas Sararenda" de edad Givetiano Inferior (Albariño et al., 2002), durante el período de máxima expansión (curva de Johnson et al., 1985). Otro ejemplo de estas importantes variaciones de facies se observan en los yacimientos Ramos y Aguaragüe (cuenca devónica de Argentina). Allí la Formación Icla presenta una edad algo mayor (Pridoliano - Lochkoviano) pero coincidente con la curva eustática de Johnson et al. (1985). Mientras que en la localidad tipo, donde dicha unidad litoestratigráfica (pelitas de plataforma externa en interior de cuenca) tiene una edad pragiana, coincide con el período de máxima contracción de la cuenca, de acuerdo al modelo.

Figura 9: Cuadro de Correlación y Equivalencias entre Unidades Cronoestratigráficas con Unidades Litoestratigráficas.
Figure 9: Correlation Chart and Equivalences between Chronostratigraphical with lithostratigraphical units.
De los perfiles y pozos analizados la mayor y mejor calidad de información obtenida es del Devónico inferior y medio donde se describieron 19 Secuencias Genéticas en el rango de los ciclos eustáticos de 3er. Orden (Fig. 8). Algunos de estos límites pudieron ser calibrados con mayor precisión porque coinciden con límites de pisos definidos por macrofauna y/o palinología. El rigor con que se definieron las secuencias dentro del intervalo de cada uno de los pisos es menor debido a la imposibilidad de disponer de líneas tiempo de mayor detalle, las que podrán ser revisadas y mejor acotadas en el futuro.
BIOFACIES
Se estudiaron y procesaron más de 70 biofacies de 10 columnas estratigráficas de superficie en sus tramos devónicos (Fig. 2 y Tabla 2). Inicialmente cada biofacies fue caracterizada independiente de su posición en cada columna y en cada Secuencia Genética, como así también de cada ciclo depositacional regresivo y transgresivo. Posteriormente, se ubicó cada biofacies dentro de cada Secuencia Genética definida y se analizaron los patrones generales de distribución de esta fauna en los ciclos regresivos/transgresivos en la plataforma silicoclástica, anteriormente definida (Albariño et al., 2002; Dalenz et al., 2002). Se dividió a las observaciones en biofacies devónicas que se desarrollaron en Secuencias de nivel alto o durante la expansión de la cuenca y biofacies devónicas que se desarrollaron en Secuencias de nivel bajo o durante la contracción.
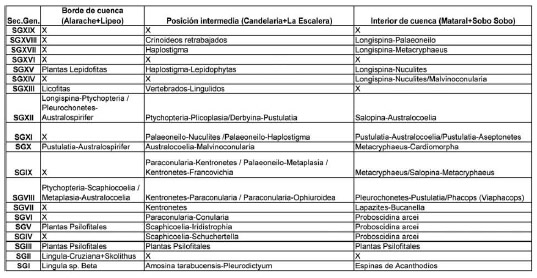
Tabla 2: Principales biofacies caracterizadas en la cuenca silúrica-devónica en borde de cuenca, posición intermedia e interior de cuenca.
Table 2: Most important biofacies from silurian-devonian basin in three positions: border external basin, middle and interior basin.
Biofacies devónicas en secuencias desarrolladas durante etapas de contracción: Durante las etapas de contracción se identificaron regresiones normales y forzadas.
Durante las regresiones normales correspondientes a un nivel eustático bajo de 2° orden del Devónico inferior, la fauna bentónica se diversificó y colonizó la plataforma marina fini-lochkoviana y pragiana a emsiana durante el desarrollo de los ciclos regresivos y en menor medida del still stand de los mismos (Fig. 10). La fauna desarrollada en la máxima inundación de cada Secuencia suele presentar diversidades bajas a muy bajas y puede sufrir numerosas condensaciones. Además, se puede afirmar que la fauna se distribuye sobre todo desde el frente de costa hasta la zona de transición y el offshore. Como posible efecto de la reducción del espacio disponible, producto de la progradación de la plataforma durante regresiones normales, la fauna va separándose en subáreas de composiciones y diversidades particulares, sin por ello llegar a la especiación alopátrica. Es decir, que en niveles del ciclo regresivo de Secuencias contemporáneas, las características de las biofacies pueden ser variables, debido a esta reducción del espacio disponible.

Figura 10: Distribución de las biofacies en dos Secuencias desarrolladas durante etapas de contracción donde se produce una regresión normal. La posición de Candelaria es del interior oeste de la cuenca y Zenta es el borde sur.
Figure 10: Biofacies distribution in two Sequences developped during contractions corresponding to normal regression. Candelaria is situated in the center west of the basin, and Zenta is at the southern border.
De acuerdo a la distribución de biofacies en secuencias regresivas normales lochkovianas a pragianas, tanto en posiciones de borde como Abra de Zenta y Alarache, así como en posiciones intermedias, Candelaria - La Escalera y en posiciones de interior como Sobo Sobo, se observan niveles de plantas fragmentadas y mal conservadas, con diversidad nula, en el highstand de la Secuencia regresiva.
En posiciones intermedias de la cuenca como Candelaria - La Escalera, se encuentra una biofacies alóctona formada por una concentración de plantas psilofitales fragmentadas de diversidad nula en el highstand del ciclo regresivo. En posiciones de interior como Sobo Sobo, se han estudiado cinco biofacies formadas todas por plantas psilofitales con una buena conservación en niveles concentrados y amalgamados co-rrespondientes al highstand y probablemente al still stand del ciclo regresivo. La presencia de fauna bentónica en el Lochkoviano tardío se concentra en niveles superiores del ciclo regresivo progradante, con el desarrollo de fauna epibentónica y aeróbica, dominada por braquiópodos que difícilmente se conservaron de la erosión del transgresivo del techo de esta Secuencia Genética. Un ejemplo de biofacies preservada está en Candelaria - La Escalera y en Presto - El Peral (obs. pers. ADF). En Candelaria - La Escalera, se ha definido la comunidad de Scaphiocoelia - Schuchertella en el fin de la progradación (still stand?) (techo de la Formación Santa Rosa) con una diversidad media (D = 1,56), compuesta por 98 % de braquiópodos como Scaphiocoelia boliviensis Whitfield, Australospirifer hawkinsi (M. y S.) entre otros, con tipos tróficos epibentónicos y suspensívoros. Suelen asociarse a esta biofacies, artejos de crinoideos sueltos. En El Peral (al norte de La Escalera) se encontró un nivel de fauna preservada en una playa muy somera, con fauna comparable (Racheboeuf, 1992). Esta es la primera colonización importante que experimenta la plataforma devónica con el desarrollo y la diversificación de la fauna bentónica, durante regresiones "normales" durante el Lochkoviano.
Entre el Pragiano medio y el Emsiano terminal, en un nivel eustático bajo de 2° orden se produce una mayor reducción y contracción de la cuenca, producida ya sea por algún movimiento de levantamiento cortical apenas distinguible (Dalenz et al., 2002), o bien por un cambio en el clima con un efecto en el volumen de material aportado a la cuenca, o bien por una caída de nivel de base de tercer orden. El producto de esta reducción de espacio en la cuenca fueron dos regresiones forzadas (Albariño et al., 2002; Dalenz et al., 2002) cuyo efecto en el bentos se traduce en fauna bentónica de diversidades muy bajas en comparación con las calculadas en Secuencias precedentes de nivel bajo de 2° orden afectadas por regresiones normales en los mismos puntos de la cuenca. En Candelaria - La Escalera se tienen tres biofacies. La primera es una biofacies alóctona, formada por fauna en nódulos calcareos de Paraconularia aff. ulrichana (Clarke), de diversidad muy baja. El segundo nivel es una biofacies parautóctona a alóctona de Conularia - Paraconularia, de paleobatimetrías de frente de costa distal(?). El tercer nivel es una biofacies parautóctona a alóctona de Australocoelia de baja diversidad. Las tres biofacies se distribuyen en el ciclo regresivo que mantiene paleobatimetrías de offshore en este punto de la cuenca. Dos de los tres horizontes tienen características alóctonas, habiendo sido transportados cuenca adentro. En Mataral se observa la biofacies de Proboscidina que se repite en tres niveles. Esta biofacies presenta rasgos de "feedback" tafonómico, con "time averaging", pues las conchillas se habrían desarrollado sobre niveles de otras conchillas presentando una mala conservación, en varias "capas" o generaciones de Proboscidina en bancos de 5 a 10 cm. Estos bancos están relacionados a niveles de condensación cuenca adentro como es Mataral para esta Secuencia. En Villagranado se tiene una biofacies que se repite en dos niveles. Esta biofacies está formada por bivalvos endobentónicos detritívoros profundos como Palaeoneilo que no parecen haber sido transportados ni haber sufrido condensación. Esta biofacies estaría marcando una paleobatimetría de frente de costa distal a offshore, siendo parautóctona.
La distribución de biofacies en esta secuencia, responde a un control paleobatimétrico, pues mientras en el interior de la cuenca se tienen biofacies de paleobatimetrías más profundas dominadas por fauna endobentónica y detritívora como el género Palaeoneilo y otras afectadas por condensación y "timeaveraging", otras responden a paleobatimetrías de frente de costa distal en puntos con desarrollo de bahías como las biofacies de Candelaria anteriormente descriptas. Además se advierten muy bajas diversidades, aún menores que en secuencias previas, indicios de retrabajo en la fauna que está concentrada en nódulos y posiciones de interior con condensaciones ya registradas en otras secuencias.
Además, se advierte el predominio de niveles donde las biofacies son retrabajadas, acumuladas mecánicamente en niveles alóctonos en posiciones de interior de cuenca que aún se mantuvieron inundados durante el desarrollo de estas regresiones (Fig. 11). Se observa la clara migración de la línea de costa cuenca adentro, con una abundante concentración de plantas fragmentadas (licofitas y lepidofitas) mejor documentada en Secuencias desarrolladas en regresiones forzadas que en normales. Igualmente, en niveles de fauna autóctona a parautóctona, se observa el desarrollo de fauna poco comparable entre puntos de batimetrías similares en la cuenca afectada por regresiones forzadas.

Figura 11: Distribución de biofacies en una Secuencia desarrollada durante una contracción donde se produce una regresión forzada.
Figure 11: Biofacies distribution in a Sequence developped du-ring contraction corresponding to a forced regression.
Esto puede ser interpretado como un efecto de la mayor reducción del espacio disponible que se produce entre una regresión forzada y otra normal. La fauna se presentaría entonces "compartimentada" en las porciones de plataforma que permanecieron inundadas durante las regresiones forzadas. Biofacies devónicas desarrolladas en el Transgresivo durante etapas de contracción: Uno de los horizontes de mayor dispersión y de mejor reconocimiento en esta cuenca es un delgado horizonte ubicado en la base del transgresivo de la Secuencia Genética IV (Albariño et al., 2002), o el techo de la Formación Santa Rosa y la base de la Formación Icla en Icla, Presto y El Peral entre otros. La edad de este transgresivo fue analizada a partir de la fauna acumulada en este horizonte. A pesar de presentar rasgos claramente erosivos en su base, este transgresivo es considerado a nivel de la cuenca, como una línea isócrona, con fauna removida del fin de ciclo regresivo precedente, pero también con fauna autóctona. En Mataral, la base del transgresivo presenta un horizonte de unos 15 cm de espesor, formado por conchillas de Proboscidina arcei (Ulrich, 1892), con una base de conchillas fragmentadas, por encima de las cuales se observan acumulaciones removidas, poco transportadas, valvas articuladas pero con una mejor preservación hacia el techo del banco de estudio. La última capa de conchillas conserva la mineralogía de las valvas primarias, sin rastro de erosión o desgaste. Esta coquina, pudo haberse formado en un fondo bentónico blando, que fue "enriquecido" por conchillas duras, erodadas por la base del transgresivo a arenas preservadas del fin de la progradación de la misma Secuencia. Estas conchillas fragmentadas, van a "tapizar" el fondo bentónico blando, cambiando las características del espectro ecológico, permitiendo la colonización de fauna, en este caso la misma especie epibentónica. El tener fauna erodada, pero no exclusivamente, haría pensar en un resago transgresivo (sensu Kidwell (1991) y Van Wagoner et al., 1990) con una retroalimentación tafonómica (Kidwell, 1991), en tiempos de diversidades muy bajas, donde P. arcei era dominante en la plataforma de fines del Lochkoviano y logra adaptarse y sobrevivir a principios del Pragiano. En Candelaria - La Escalera, la fauna más vieja presente en el transgresivo es de edad lochkoviana terminal, por lo tanto el evento transgresivo tuvo que ser por lo menos postlochkoviano. Alguna fauna autóctona desarrollada ya sea en la base del transgresivo como en la máxima inundación sería pragiana como Iridistrophia cf. aliciae?, Scaphiocoelia boliviensis y P. arcei del Pragiano basal a partir de la asociación palinomórfica de Angochitina aff. comosa del Pragiano basal de Europa, según Le Herissé en Racheboeuf (1993). El estudio de la composición de las biofacies desarrolladas en este transgresivo tanto en puntos del borde de esta cuenca (Abra de Zenta, Lipeo, Alarache, en el noroeste argentino) como del centro (Mataral, Sobo Sobo en el centro de Bolivia), indica que la composición faunística de la base del transgresivo depende de la composición y de las características de la fauna desarrollada en los niveles progradantes que luego fueron erodados por la base de este transgresivo (Fig. 10).
Biofacies devónicas en Secuencias desarrolladas durante etapas de expansión: Durante el Devónico medio se registra un aumento de nivel de base de 2°orden, que permite la incursión de fauna boreal al Gondwana occidental, como los braquiópodos chonétidos Longispina, Chonostrophia, Anoplia y Montsenetes (Racheboeuf e Isaacson, 1993). Este es un evento biológico reconocido en el Gondwana a través de otros grupos faunísticos, como los cefalópodos nautiloideos (Crick, 1990), goniatites (que podría corresponder al "evento Chotec" de Chlupac, 1994), blastoideos (Waters, 1990), phylocáridos (Hannibal et al., 1993) y bivalvos (Dalenz Farjat, 2000). En la cuenca analizada las biofacies eifelianas son de menor diversidad (D = 1,33 de promedio, Dalenz Farjat, 2000) que las biofacies emsianas y pragianas de nivel bajo producidas durante regresiones normales pero no así las producidas durante regresiones forzadas. Composicionalmente, son biofacies distintas, con géneros claramente boreales, que no se registraron antes en la cuenca. La fauna es más diversa en la parte inferior a media de los ciclos regresivos y no en el techo de los mismos como en el caso de las Secuencias emsianas en etapas de contracción de la cuenca, probablemente porque la fauna eifeliana es fauna de paleobatimetrías mayores y niveles menores de energía (Fig. 12). Suelen encontrarse tipos tróficos endobentónicos y detritívoros acompañando a los epibentónicos y suspensívoros como en Bala Puca se tiene la biofacies de Palaeoneilo pondeana y Australospirifer - Longispina. Las composiciones faunísticas son distintas a las emsianas gondwánicas epibentónicas, aeróbicas y con individuos adaptados a arenas de energía alta a media. Mientras en el Emsiano existe un predominio de espiriféridos, centronélidos y chonétidos del tipo Pleurochonetes y Kentronetes, en el Eifeliano existe un predominio de chonetidos pequeños del tipo Longispina, Anoplia, Montsenetes y Chonostrophia, algunos bivalvos como Palaeoneilo constricta son frecuentes, notando una clara disminución de espiriferidos.
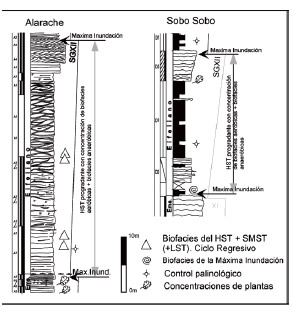
Figura 12: Distribución de biofacies en la Secuencia Genética XII en Alarache o borde sur de la cuenca y Sobo Sobo o interior. Notar que las biofacies se concentran en la parte inferior y media del highstand regresivo.
Figure 12: Biofacies distribution in Genetical Sequence XII in Alarache, southern border of the basin, and Sobo Sobo or center of the basin. Biofacies are concentrated at the bottom of the regressive highstand.
Modelo de distribución de biofacies en la cuenca (Fig.13): Las biofacies del Devónico inferior desarrolladas durante un nivel bajo de 2° orden, son más diversificadas y más frecuentes que las biofacies del Devónico medio desarrolladas en niveles altos de 2° orden, siempre que ambas comparaciones se hagan en regresiones normales (44 biofacies en total en la Secuencia VIII en todos los puntos estudiados del área frente a 18 biofacies de la Secuencia XII). Las biofacies del Devónico inferior se encuentran en el ciclo regresivo de mayor progradación de litofacies de borde. La composición de la fauna se empobrece y la diversidad disminuye tanto en biofacies del still stand por posible compartimentación del reducido espacio disponible de la zona litoral o de borde (?) y en la máxima inundación donde parece que se acentúan las condiciones anóxicas y poco propicias para la colonización de fauna aeróbica.

Figura 13: Dos Secuencias Genéticas tipo, donde se ilustra la ubicación y tipo de biofacies devónicas que con mayor frecuencia fueron registradas en el area de trabajo. Ambas están definidas por superficies de máxima inundación.
Figure 13: Two Genetical Sequences to illustrate the place and the type of devonian biofacies more frequently registered in the studied area. Those Sequences are defined with maximum flooding surfaces.
La base de los transgresivos es erosiva y su composición faunística dependerá de la composición y las características ecológicas que mantenga el final del ciclo progradante precedente. Las biofacies del Devónico medio desarrolladas durante los niveles altos del mar, presentan biofacies menos diversas que las del Devónico inferior en procesos regresivos normales, composicionalmente formadas con influencia boreal, desarrolladas en paleobatimetrías de frente de costa distal a offshore y con presencia de fauna endobentónica y detritívora además de la fauna epibentónica. La fauna se desarrolla en la parte basal y media de los ciclos regresivos. Esta variación de fauna entre el Devónico inferior y medio tiene que ver con un control de ingreso de fauna boreal favorecida por una subida de nivel de base de 2° orden que se registra en todo el Gondwana a partir del Eifeliano. Las biofacies desarrolladas en regresiones normales durante etapas de expansión o de contracción de la cuenca, son más numerosas y más frecuentes por Secuencia progradante que aquellas desarrolladas en regresiones forzadas (varían entre 44 a 18 biofacies en las Secuencias analizadas en regresiones normales y de 19 a 5 en las Secuencias de nivel bajo en regresiones forzadas), con una mayor distribución areal. En condiciones de regresión normal, las biofacies aeróbicas están formadas por comunidades parautóctonas a autóctonas, desarrolladas en el ciclo regresivo y el still stand y en menor medida en la máxima inundación. En este tipo de regresiones, se observa una compartimentación incipiente de la plataforma, lo cual hace que las biofacies puedan ser algo más homogéneas entre sí, en composición, valores de diversidad y tipos tróficos. Las biofacies de nivel bajo de 2° orden producidas durante regresiones forzadas, son las menos numerosas por Secuencia y tienen la menor distribución areal debido a la contracción avanzada de la cuenca. La fauna se presenta retrabajada en mayor proporción que en las biofacies de nivel bajo de 2º orden desarrolladas durante regresiones normales. Las asociaciones son casi todas alóctonas y se advierten numerosos niveles de nódulos en las máximas inundaciones en posiciones de interior de cuenca.
Se advierte una alta frecuencia de fragmentos de plantas a lo largo de todas las Secuencias, como prueba de la migración de la línea de costa cuenca adentro. Se considera avanzada la compartimentación de la plataforma, con biofacies contemporáneas muy heterogéneas entre sí, tanto en composición como en diversidad. En plataformas silicoclásticas tipo 2, donde la pendiente parece reducida como sería el caso de la cuenca analizada, el desarrollo de la fauna bentónica parece estar muy condicionado a las variaciones del espacio disponible en primera instancia, y posteriormente a los efectos de las variaciones eustáticas de 2° orden. Por otro lado, se puede interpretar que una regresión forzada durante un nivel bajo de 2° orden (etapa de contracción) en este tipo de plataformas, puede tener un efecto mayor y determinante en la limitación del desarrollo de la fauna, mucho más que si se produce en una etapa de expansión de la cuenca.
CONCLUSIONES
Se ha establecido un modelo de distribución de facies, en una plataforma silicoclástica marina, dominada por olas, en un lapso de tiempo desde el Ludlowiano al Frasniano. Esta parte sur y suroeste de la cuenca analizada, desarrolla ambientes sedimentarios de plataforma sin alcanzar el talud. La depositación en esta plataforma estuvo controlada por variaciones eustáticas y por variaciones de volumen de material aportado, sufriendo al menos tres eventos de regresiones forzadas que determinaron la depositación de arenas cuenca adentro. En esta cuenca las biofacies estuvieron fuertemente controladas por estos procesos ambientales y eustáticos. Este trabajo es el resultado de la integración del estudio de bioestratigrafía, facies, ambientes y biofacies en un contexto de análisis secuencial, lo que ha permitido comprender la dinámica de esta cuenca. Con este modelo se han replanteado estrategias exploratorias en el Devónico del área, conociendo nuevas distribuciones de reservorios y rocas sello.
Agradecimientos: Este trabajo constituye parte del estudio de la cuenca Silúrico-Devónica del subandino sur boliviano y noroeste argentino elaborado por las empresas XR s.r.l. y Pluspetrol entre los años 1997 y 2000. Los autores agradecen a Pluspetrol que les permitió publicar datos de su propiedad. Igualmente agradecen a Cristina Vistalli (Repsol - YPF), Daniel Poiré (Universidad Nacional de La Plata), Ricardo Gomez Omil (Pionner) y a todos los colegas que de una u otra manera aportaron ideas a la discusión del Silúrico- Devónico.
REFERENCIAS BIBLIOGRAFICAS
1. Albariño, L., A. Dalenz Farjat, L. Alvarez, R. Hernández y M. Perez Leyton, 2002. Las Secuencias Sedimentarias del Devónico en el Subandino Sur y el Chaco. Bolivia y Argentina. Quinto Congreso Exploración y Desarrollo de Hidrocarburos. CD Trabajos Técnicos. Mar del Plata. [ Links ]
2. Aceñolaza, F. G. y A. J. Toselli, 1973. Consideraciones estratigráficas y tectónicas sobre el Paleozoico Inferior del Noroeste Argentino. Segundo Congreso Latinoamericano Geológico, Actas 2:755-783. Caracas. [ Links ]
3. Arche, A., 1992. Sedimentología (Nuevas Tendencias). Consejo Superior de Investigación Científica. Volumenes I y II, 543 pp. y 528 pp. Madrid. [ Links ]
4. Bambach, R. K., 1983. Ecospace utilization and guilds in marine communities through the Phanerozoic. En M. J. S. Tevesz, y P.L. McCall (Eds.), Biotic interactions in Recent and Fossil benthic communities. Topics in Geology. Plenum Press.:719-146. [ Links ]
5. Coira, B., J. Davidson, C. Mpodozis y V. Ramos, 1982. Tectonic and magmatic evolution of the Andes of northern Argentina and Chile. Earth Sciences Review 18:303-332. [ Links ]
6. Crick, R. E., 1990. Cambro-Devonian biogeography of Nautiloid cephalopods. En W. S. McKerrow y C. R. Scotese (Eds.), Palaeozoic, Palaeogeography and Biogeography. Geologic Society Memoir 12:147-161. [ Links ]
7. Dalenz Farjat, A., 2000. Taxonomía, Paleoecología y Paleogeografía de Moluscos Bivalvos del Siluro-Devónico del Altiplano, Cordillera Oriental, Interandino y Subandino de Bolivia. Tesis Doctoral, Universidad Nacional de Córdoba. Tomos I y II. 351 pp. (inédito). [ Links ]
8. Dalenz Farjat, A., L. A. Alvarez, R. M. Hernández y L. M. Albariño, 2002. Cuenca Siluro-Devónica del Sur de Bolivia y del Noroeste Argentino: algunas interpretaciones. Quinto Congreso de Hidrocarburos. CD Trabajos Técnicos. Mar del Plata [ Links ]
9. Dalla Salda, L., C. Cingolani y R. Varela, 1993. A pre - Carboniferous tectonic model in the evolution of South America. Comptes Rendus XII ICCP 1:371-384. [ Links ]
10. Dickinson, W. R. y D. R. Seely, 1979. Structure and Stratigraphy of Forearc Regions. The American Association Petroleum Geology Bulletin 63:2- 31. [ Links ]
11. Galloway, W.E., 1989. Genetic Stratigraphic Sequences in Basin Analysis I: Architecture and Genesis of Flooding - Surface Bounded Depositional Units. American Association of Petroleum Geologists 73:125-142. [ Links ]
12. Hannibal, J. T., R. M. Feldmann y W. D. I. Rolfe, 1993. Phyllocarid crustaceans from the Devonian of Bolivia. En R. Suárez-Soruco (Ed.), Fósiles & Facies de Bolivia, II, Invertebrados y Paleobotánica. Revista Técnica YPFB, 13-14 (1-4):59-69. Santa Cruz. [ Links ]
13.Johnson, J. G., G. Klapper y C. A. Sandberger, 1985. Devonian eustatic fluctuations in Euramerica. Bulletin of Geological Society of America 96:567-587. [ Links ]
14. Kidwell, S. M., 1991. Taphonomic Feedback (Live/Dead interactions) and the genesis of bioclastic beds: keys to reconstructing sedimentary dynamics. En G. Einsele, W. Ricken y A. Seilacher (Eds.), Cycles and Events in Stratigraphy. Springer-Verlag:268-282. Tubingen. [ Links ]
15. Racheboeuf, P., A. Le Hérissé, F. Paris, C. Babin, F. Guillocheau y M. Truyolls - Massoni, 1993. Le Dévonien de Bolivie: Biostratigraphie et chronostratigraphie. Comptes Rendus Académie des Sciences de Paris 317:795-602. [ Links ]
16. Racheboeuf, P. R. y P. E. Isaacson, 1993. Los chonetoideos (Braquiopodos) silúricos y devónicos de Bolivia. En R. Suárez-Soruco (Ed.), Fósiles y Facies de Bolivia II, Invertebrados y Paleobotánica. Revista Técnica YPFB 13/14:99-119. [ Links ]
17. Ramos, V., 1988. Proterozoico tardío-Paleozoico temprano de Sud America. Una historia de colisiones, Episodes 11:168-174. [ Links ]
18. Ramos V. A., T. E. Jordan, R. W. Allmendinger, C. Mpodozis, S. Kay, J. M. Cortes y M. Palma, 1986. Paleozoic terranes of the Central Argentine - Chilean Andes. Tectonics 5:855- 880. [ Links ]
19. Rapela, C. W., B. Coira, A. Toselli y J. Saavedra, 1992. El magmatismo del Paleozoico inferior en el Sudoeste de Gondwana. En J. C. Gutierrez- Marco, J. Saavedra y I. Rábano (Eds.), Paleozoico Inferior de Iberoamérica. Universidad de Extremadura 21-68. Mérida. [ Links ]
20. Reading, H. G., 1986. Sedimentary environments and facies. Blackwell Scientific Publications. Second Edition. 615 pp. [ Links ]
21. Richardson, J. B. y D. C. McGregor, 1986. Silurian and Devonian spores zones of the Old Red Sandstones Continent and adjacent region. Bulletin Geological Survey of Canada 364:1- 79. [ Links ]
22. Salfity, A. J., S. A. Gorustovich y M. C. Moya, 1984. Las fases diastróficas en los Andes del Norte Argentino. International Simposium of Central- Andean Tectonics and Relations with Natural Resources. Academia Nacional de Ciencias de Bolivia. La Paz. [ Links ]
23. Sanchez, T. M., 1984. Etude paléoécologique du Paléozoique Supérieur dans le Nord des Andes du Vénézuela. Esquisse Systématique des mollusques bivalves. En P. Racheboeuf y C. Babin (Eds.), Biostratigraphie du Paleozoique 2:1- 165. [ Links ]
24. Sanchez, T. M., B. Waisfeld y J. L. Benedetto, 1991. Lithofacies, taphonomy and brachiopod assemblages in the Silurian of western Argentina: a review of malvinokaffric realm communities. Journal of South American Earth Sciences 4:307-329. [ Links ]
25. Sanders, H. L., 1968. Marine benthic diversity: a comparative study. The American Naturalist. 102: 243-268. [ Links ]
26. Selley, R. C., 1998. Elements of Petroleum Geology. Academic Press. Second Edition. New York. [ Links ]
27. Starck, D., E. Gallardo y A. Schulz, 1992. El Siluro- Devónico de la Cuenca Subandina y su Potencial Exploratorio. Informe Interno YPF. Comisión Geológica N° 4. (inédito). [ Links ]
28. Starck, D., 1995. Silurian - Jurassic Stratigraphy and Basin Evolution of Northwestern Argentina. En A. J. Tankard, R. Suárez-Soruco y H. J. Welsink (Eds.), Petroleum Basins of South America. American Association of Petroleum Geologists, Memoir 62:251-267. [ Links ]
29. Stewart, J. y A. B. Watts, 1997. Gravity anomalies and spatial variations of flexural rigidity at mountain ranges. Journal of Geophysical Research 102:5327-5352. [ Links ]
30. Streel, M., K. Higgs, S. Loboziak, W. Riegel y Ph. Steemans, 1987. Spore Stratigraphy and Correlation with fauna and floras in the type marine Devonian of the Ardenne Rhenish regions. Review of Paleobotany, Palynology 50:211-229. [ Links ]
31. Suárez Soruco, R., 1999. Bolivia, Proterozoico Superior - Paleozoico Inferior. Fourth International Symposium of Andean Geodynamics. Gottingen. [ Links ]
32. Suárez Soruco, R., A. Dalenz Farjat, Legrand Smith, 2000. Subdivisión tectono-estratigráfica de las rocas del Ciclo Cordillerano de Bolivia. Catorce Congreso Geológico de Bolivia. CD Trabajos Técnicos. La Paz. [ Links ]
33. Van Wagoner, J., R. M. Mitchum, K. M., Campion y V. T., Rahmanian, 1990. Siliciclastic sequence stratigraphy in well logs, cores and outcrops: concepts for high resolution correlation of time and facies. American Association of Petroleum Geologists, N7, 55p. [ Links ]
34. Walker, K. R. y R. K., Bambach, 1974. Feeding by benthic invertebrates: classification and terminology for paleoecological analysis. Lethaia 7:67- 78. [ Links ]
35. Waters, J. A., 1990. The palaeobiogeography of the Blastoidea (Echinodermata). En W. S. McKerrow y C. R. Scotese (Eds.), Palaeozoic, Palaeogeography and Biogeography, Geologic Society Memoir 12:339-352. [ Links ] AAS

