










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo
On-line version ISSN 1853-8665
Rev. Fac. Cienc. Agrar., Univ. Nac. Cuyo vol.43 no.2 Mendoza July/Dec. 2011
ARTÍCULOS ORIGINALES
Influence of fertilization on the growth of radicchio "Rosso di Chioggia" cultivated in two different environments
Influencia de la fertilización en el crecimiento de radicchio "Rosso di Chioggia" cultivado en dos ambientes diferentes
Maria Flavia Filippini 1, Carlo Nicoletto 2, Paolo Sambo 2, Juan Bruno Cavagnaro 1 y Ferdinando Pimpini 2
1Department of Agricultural Engineering and Biological Sciences. Facultad de Ciencias Agrarias. UNCUYO. Alte. Brown 500. Chacras de Coria. Mendoza. Argentina. M5528AHB.
2Department of Environmental Agronomy and Crop Science. University of Padova. Viale dell'Università, 16. 35020 Legnaro. Italy. carlo.nicoletto@unipd.it
Recepción: 20/04/2011
Aceptación: 20/10/2011
ABSTRACT
This study evaluated the influence of fertilization on some growth indexes in radicchio "Rosso di Chioggia". The trial was conducted in Rovigo (Italy) for two years and in Mendoza (Argentina) for one year. Increasing doses of macronutrients were considered, identified as N0P0K0, N1P1K1, N2P2K2 and, only in Rovigo, N3P2K2. Some indexes related to growth analysis: relative growth rate (RGR), net assimilation rate (NAR), leaf area ratio (LAR), specific leaf area (SLA), leaf weigh ratio (LWR), crop growth ratio (CGR), leaf area index (LAI) and leaf area duration (LAD) were calculated during the growing cycle. At Mendoza CGR was strongly influenced by NAR from transplant until 776 growth degree days (GDD). After that moment, CGR was affected by LAI that increased markedly due to favourable environmental conditions. From 1052 to 1653 GDD the high increase of LAI determined a reduction in photosynthetic efficiency. At Rovigo the trends in index values showed differences between the two years considered, especially in LAR and SLA. In the second year CGR had always higher values; mainly due to higher LAI values, since NAR did not differ significantly during the years. In the second half of the growing cycle, CGR was instead strongly linked to a contraction of photosynthetic effectiveness, due to head formation. High LAI values showed a growing cycle extension to the detriment of early head formation. It was also observed that when LWR is around 0.35 - 0.40 g g-1, the plant reaches marketable maturation and is ready to be harvested. In both environments, no clear effects of fertilization were observed on the growth indexes. In any case highly fertilized plants showed higher growing activity especially at the beginning of the growing cycle.
Keywords: Cichorium intybus L.; Group rubifolium; Growth analysis; Fertilization.
RESUMEN
Este trabajo evalúa la influencia de la fertilización en radicchio tipo "Rosso de Chioggia" (precocidad media) sobre algunos índices de crecimiento. Se realizaron ensayos durante dos campañas en Rovigo (Italia) y una en Mendoza (Argentina), aplicándose dosis crecientes de NPK, identificadas como N0P0K0, N1P1K1, N2P2K2, en Mendoza y en Rovigo, además, N3P2K2. Durante el cultivo se calcularon índices de crecimiento como: relative growth rate (RGR), net assimilation rate (NAR), leaf area ratio (LAR), specific leaf area (SLA), leaf weigh ratio (LWR), crop growth ratio (CGR), leaf area index (LAI) and leaf area duration (LAD). En Mendoza, el CGR estuvo fuertemente influenciado por NAR desde el trasplante hasta alcanzar 776 grados días (GDD); desde 1052 a 1653 GDD el CGR fue afectado por el LAI el cual aumentó marcadamente debido a las condiciones ambientales favorables. Entre los 1052 y 1653 GDD el incremento del LAI determinó una reducción en la eficiencia fotosintética. En Rovigo, la tendencia de los índices fue disímil en los dos años, encontrándose respuestas diferentes en LAR y en SLA. En el segundo año, el CGR siempre arrojó valores más altos, mientras que NAR no difirió en ninguno de los años. En la segunda mitad del ciclo, CGR estuvo fuertemente asociado a una menor eficiencia fotosintética, debido a la formación de la cabeza. Valores elevados de LAI indicaron una extensión del ciclo, retrasando la formación de la cabeza. Las plantas alcanzaron la madurez comercial con LWR entre 0,35 - 0,40 g g-1. En ambos ambientes, no se observó claramente el efecto de la fertilización sobre los índices; si bien las dosis más altas mostraron mayor actividad de crecimiento en las etapas tempranas.
Palabras clave: Cichorium intybus L.; Grupo rubifolium; Análisis de crecimiento; Fertilización.
INTRODUCTION
The increasing commercial interest expressed in radicchio (Cichorium intybus L., group rubifolium) during the last years is justified by both leaf characteristics and their nutritional features. A continuous expansion of areas cultivated with this chicory has been observed since the 1980s not only in Italy, but also in other parts of Europe (e. g. France, Germany, Spain, Holland) and trans ocean countries (e. g. Japan, USA, Chile and Argentina). This relevant cultivation increase has not been followed by an equal research interest, especially from a physiological point of view, topics where this species presents many problems to be solved.
Up to now only fowering dynamics, its conditioning factors (5, 6, 20, 21) and qualitative properties (12, 13, 16, 17, 22, 24) has been considered. No information has been found concerning plant growth analysis, nutrients uptake rates and nutrients translocation inside the plant. Growth dynamics have to be studied to understand the physiological processes occurring in plants growing in different environmental and cultivation conditions. Many and conclusive results have been obtained using some indexes derived from growth analysis for the most important herbaceous species (e. g. maize, soybean, wheat, sugar beet, tomato, potato, lettuce, spinach and broccoli) (1, 4, 7, 10, 14, 23, 26, 28).
Experiments that aim to understand different crop management efficiency (e. g. irrigation, fertilization, climate parameters, etc.) through observation of some growth indexes like LAI, RGR, CGR, LAR, NAR, LWR, SLA, LAD are also frequent. Van Holsteijn (10) observed that lettuce plants cultivated in a greenhouse reached higher CGR values during head formation, then the CGR index decreased until the end of crop cycle. During head formation phase LAR decreased, while NAR and RGR did not display any correlation. Spinach dry matter as well as LAI, CGR, RGR and NAR increased after nitrogen application as ammonium nitrate (25). In an experiment with tomato, Graifenberg et al. (8) found that CGR of whole plant and leaves (including petioles) was the only parameter significantly correlated with N, P, K Ca and Mg uptake. Concerning the fruits, correlation was significant only with P and K and sometimes also with Mg. LAI, LAD and NAR showed no correlation with nutrients uptake.
When water stress was applied during the critical phase in onion (the beginning of bulb enlargement) a significant negative effect on leaves and bulb dry matter accumulation was obtained (27). Leaves RGR and NAR decreased during the period of water deficiency, but increased following re-watering because of higher leaves dry weight, though functional leaf area was unchanged. During water stress NAR values also decreased and the low bulb fresh weight achieved at the cycle end was due to a lower growth of the photo-assimilating system.
The above papers show that plant growth analysis and growth indexes derived from it can give useful information for better crop management in order to optimize yield. Since these aspects have never been considered for radicchio, this study was undertaken to obtain further physiological information for this type of chicory. The experiment evaluated the effect of nitrogen, phosphorus and potassium fertilization on growth indexes in radicchio "Rosso di Chioggia" cultivated under different environmental conditions.
MATERIALS AND METHODS
The trials were carried out on radicchio "Rosso di Chioggia" (medium precocity class) at Luján de Cuyo, Mendoza (35° 52' S, 68° 52' W) - Argentina - for one year and in Veneto, Rovigo (44° 49' N, 12° 32' E) - Italy - at the "Po di Tramontana" Horticultural Experimental Station for two years.
Soil physical and chemical characteristics are reported in table 1. Values showed a good N content and intermediate P and K at Mendoza. At Rovigo, instead, a low N content was observed in the first year and good in the second one, P and K contents were good.
Table 1. Chemical and physical characteristics of the soil at the beginning of the experiments.
Tabla 1. Características fisicas y químicas del suelo al inicio de los experimentos.

Different fertilization formulae were tested:
Mendoza:
0 kg ha-1 N + 0 kg ha-1 P2O5 + 0 kg ha-1 K2O (N0P0K0 – F0)
60 kg ha-1 N + 80 kg ha-1 P2O5 + 50 kg ha-1 K2O (N1P1K1 – F1)
120 kg ha-1 N + 160 kg ha-1 P2O5 + 100 kg ha-1 K2O (N2P2K2 – F2)
Rovigo:
0 kg ha-1 N + 0 kg ha-1 P2O5 + 0 kg ha-1 K2O (N0P0K0 – F0)
100 kg ha-1 N + 100 kg ha-1 P2O5 + 100 kg ha-1 K2O (N1P1K1 – F1)
200 kg ha-1 N + 200 kg ha-1 P2O5 + 200 kg ha-1 K2O (N2P2K2 – F2)
300 kg ha-1 N + 200 kg ha-1 P2O5 + 200 kg ha-1 K2O (N3P3K3 – F3)
Further information on crop management is reported in table 2.
Table 2. Crop management practices during the experiments.
Tabla 2. Manejo del cultivo realizado durante los experimentos.

In Mendoza, in addition to information presented in table 2, the total amount of P2O5 (triple super phosphate 46%) and half quantity of K2O (sulfate of potash 50-52%) was applied at transplanting. The remaining amount of this nutrient was given during crop cultivation together with the first distribution of N (urea 46%), and the rest of the N was applied 15 days later. Before transplanting 2 L ha-1 of Trefan (48% trifuralin) was distributed to control weeds. Furrow irrigation was applied after transplanting every 7-10 days. Because of abnormal high rainfall on that season two treatments with sulfur + 1 kg ha-1 of Rovral (50% iprodione) was applied to protect plants from powdery mildew and Botritis sp.
At Rovigo, in both years, all the P2O5 (perphosphate 18-20%), K2O (potash sulfate 50-52%) and 1/3 of the total N quantity (ammonium nitrate 26-27%) were distributed with basal fertilization. The remaining N was distributed locally on two occasions during crop cultivation. A soil de-infestation was carried out before transplanting with 30 kg ha-1 of Geofos D (4.75% diazinone). After transplanting, sprinkler irrigation was applied during three days with a total volume of 200-300 m3 ha-1.
During the first year of cultivation at Rovigo, in the first ten days of August, because of a technical problem involving the entire district water disposal system, all Experimental Station outfalls were fooded by seawater. This event caused a marked increase in electrical conductivity (EC) in the soil solution increasing from 15.7±0.1 mS cm-1 at 0.3-0.4 m depth to 17.2±0.1 mS cm-1 at 0.0-0.1 m depth. High EC levels determined an abnormal plant growth and a high mortality rate (over 70%) in some areas.
From transplant to commercial plant maturation, identifed by a compact head, growth analyses were done every 10-11 days. A variable number of plants were used at each sampling date, (50 plants at samplings 1 and 2; 20 plants at samplings 3 and 4, and 5 plants in the next samplings). Plants were harvested and weighed, removing rotten and dried leaves. Photosynthetic active leaves and leaves forming the compact head (not photosynthetic) were then counted.
In addition, for sampling dates near commercial maturation, after removing rotten leaves, those remaining were divided into: outer, intermediate (partially wrapped on head (first 3-4) and inner leaves forming the commercial head.
After reaching commercial maturation, samplings were stopped when plants started to present completely rotten outer leaves (end of June at Mendoza and end of October in both experiments at Rovigo). Leaf area was determined on photosynthetic active leaves with a leaf area meter (Delta-T Devices) and leaf dry matter by oven-drying at 65 °C.
These measurements were used to calculate LAI, RGR, CGR, LAR, NAR, LWR, SLA and LAD with the classic approach (11) given that samples were collected with a time gap ranging from 7 to 25 days.
Growth parameters were calculated according to Hunt (11):
All data were statistically processed by means of ANOVA. Data from each trial were processed separately because of different fertilization amounts applied in the two locations; crop cycle length and the consequent sampling number did not allow all data to be considered together.
Meteorological data
Máximum and mínimum temperatures, relative humidity (RH) and total rainfall recorded at each experimental meteorological station are reported in figures 1, 2 and figure 3.

Figure 1. (Mendoza - Argentina) Ten-day averages of maximum and minimum temperatures (°C) and relative humidity (RH %) and ten-day cumulative rainfall (mm), collected from 3rd period of January to 3rd period of May.
Figura 1. (Mendoza - Argentina) Promedio de 10 días de las temperaturas máximas y mínimas (°C) y de la humedad relativa (RH %), y lluvia acumulada de 10 días (mm), medidas desde el 3rd período de enero al 3rd período de mayo.

Figure 2. (Rovigo - Italy) First year experiment. Ten - day averages of maximum and minimum temperatures (°C) and relative humidity (RH %) and ten - day cumulative rainfall (mm), collected from 1st period of August to 3rd period of October.
Figura 2. (Rovigo - Italia) Primer año de experiencia. Promedio de 10 días de las temperaturas máximas y mínimas (°C) y de la humedad relativa (RH %), y lluvia acumulada de 10 días (mm), medidas desde el 1er período de agosto al 3er período de octubre.

Figure 3. (Rovigo - Italy) First year experiment. Ten-day averages of maximum and minimum temperatures (°C) and relative humidity (RH %) and ten-day cumulative rainfall (mm), collected from 1st period of August to 3rd period of October.
Figura 3. (Rovigo- Italia) Segundo año de experiencia. Promedio de 10 días de las temperaturas máximas y mínimas (°C) y de la humedad relativa (RH %), y lluvia acumulada de 10 días (mm), medidas desde el 1er período de agosto al 3er período de octubre.
A higher temperature range (At = 13 °C) was observed at Mendoza than Rovigo (At = 10°C). Máximum temperatures recorded at Mendoza during the season ranged between 30.8 and 13.3°C and mínimums between 17.5 and 0°C. At Rovigo máximum temperatures did not differ from Mendoza (30°C in the first year and 33°C in the second), mínimum valúes were higher than Mendoza ranging between 18 and 7°C in 1993 and 22°C and 8°C in 1994. A high variability was also observed for RH.
At Mendoza values ranged between 53 and 75%, while at Rovigo they were higher, with minimum around 75% and maximum up to 95% in both experiments. In both environments and years rainfall was distributed throughout the crop cycle, with similar frequency, but higher volumes at Rovigo. In this location, 200 mm per year were recorded, while at Mendoza rainfall was about 140 mm.
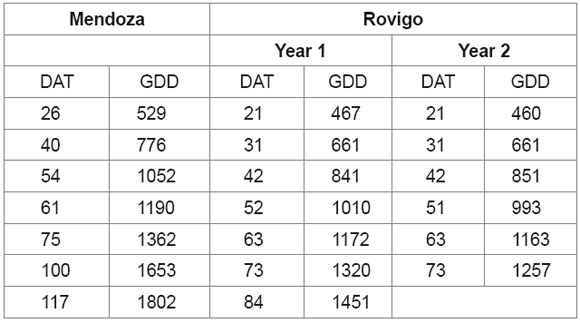
During the growing cycle, growing degree days (GDD) were considered in each location. For their calculation a base temperature of 2 °C was considered (Pimpini, personal information), the formula for radicchio being [(Tmin + Tmax)/2]-2. For clarity, both GDD and days after transplant (DAT) are reported in table 3.
Table 3. Growing degree days (GDD) and days after transplant (DAT) obtained during the growing cycle in each environment.
Tabla 3. Grados días (GDD) y días después de transplante (DAT) obtenidos durante el ciclo de cultivo en cada ambiente.
RESULTS
The infuences of fertilization and environment on some growth indexes and their possible correlations with yield are reported. Yield data, already published in a previous paper (19), are reported in table 4 to make results easier for the reader to understand. These data showed that increasing fertilization doses did not affect yield at Mendoza (Me), contrary to Rovigo in the first (Ro1) and second year (Ro2) where it increased.
Table 4. Fertilization infuence on total, marketable yields (t ha-1) and discard (%)(19).
Tabla 4. Infuencia de la fertilización sobre el rendimiento comercial total (t ha-1) y el porcentaje (%) de descarte (19).
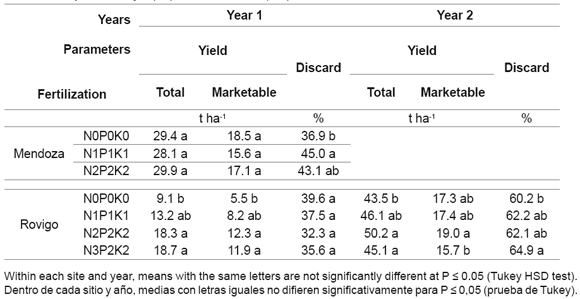
Within each site and year, means with the same letters are not significantly different at P ≤ 0.05 (Tukey HSD test). Dentro de cada sitio y año, medias con letras iguales no diferen significativamente para P ≤ 0,05 (prueba de Tukey).
Growth indexes in different environments and years
Growth indexes, that are taken in account for the first time in radicchio, showed that in both environments and years the growing cycle covered different periods, lasting for 1802, 1451 and 1257 growing degree days (GDD) in Me, Ro1 and Ro2, respectively (figure 4).

Figure 4. Main growth indexes calculated for the three experiments (Me = Mendoza; Ro1 = Rovigo first year; Ro2 = Rovigo second year). Figura 4. Principales índices de crecimiento calculados para los tres experimentos (Me = Mendoza; Ro1 = Rovigo primer año; Ro2 = Rovigo segundo año).
Figure 4 also shows differences in growth indexes, sometimes quite relevant, between environments and years. Concerning leaf area index (LAI), except for the first period of the growing cycle (529-1052 GDD), this index did not differ in the experimental areas. After that, Me values showed large increases compared with Ro1 and Ro2. Me reached its maximum value (8.77) at 1653 GDD. In Italy, Ro1 was always lower than Ro2. Maximum peaks reached at Rovigo were 1.75 at 1320 GDD and 3.13 at 1163 GDD, 80% and 65% lower than Me. The high LAI at Mendoza (5.0 and 2.8 times larger than Ro1 and Ro2 respectively) could be explained by the normal mild autumn conditions of this region, which also allowed a larger life cycle. Similarly to LAI, LAD did not differ in either location until 1190 GDD. After that LAD in Me showed large increases compared with Ro1 and Ro2, which always presented lower values (just 0.35 to 0.17 of that obtained in Me).
As expected LWR showed a decreasing trend during the cycle, since it represents the ratio of photosynthetic leaves biomass relative to total plant biomass. After 1052 GDD, Me expressed higher values than Ro1 and Ro2. In the latter location the first year (Ro1) LWR was higher than Ro2 during the whole growing cycle. At harvest time all experiments showed LWR of 0.38, although harvesting occurred at 1802 GDD in Me.
SLA increased with plant aging. In particular it ranged from 0.026 to 0.037 m2 g-1 in Me and from 0.014 to 0.024 m2 g-1 in Ro1. Instead, in Ro2, SLA decreased during the growing cycle from 0.025 to 0.019 m2 g-1.
LAR, a composite parameter partly determined by allocation (LWR) and leaf morphology (SLA), decreased in both environments. Me values were always higher than Ro1 and Ro2, ranging from 0.026 m2 g-1 at the beginning to 0.014 m2 g-1 at the end of the cycle. At Rovigo, higher values were obtained in Ro2 until 851 GDD, but after that date Ro1 overpass Ro2 until the end of the cycle.
NAR represent the leaves net CO2 assimilation rate. This index increased until 851 GDD at Rovigo and 1052 GDD at Mendoza, then decreased in both environments. During the early part of the cycle values at Rovigo were higher than at Mendoza. This index showed almost the same values in Ro1 and Ro2 until 1172 GDD. At the last sampling time Me expressed the lowest value of close to 0 g m-2 d-1, while Ro2 still showed quite a high index (2.25 g m-2 d-1).
Relative growth rate (RGR) presented, on the whole, a strong increase at the beginning of the growing cycle and then declined until the end of the cycle in both environments and years. The general trend of RGR was very similar to NAR showing a better association with the physiological component (NAR) than with the morphological one (LAR). Maximum values of RGR (0.11 g g-1 d-1) were obtained in Ro2 while maximums for Ro1 and Me were just around 0.09 g g-1 d-1. The larger values in Mendoza after 1000 GDD can be attributed to a larger LAR since NAR was similar to Rovigo.
CGR showed a similar trend in both locations and years, reaching a maximum around 1000 GDD and then decreasing with different slopes. Maximum values were similar in Me (14.62 g m-2 d-1) and Ro2 (12.4 g m-2 d-1), while in Ro1 it was less than a third of the others.
Growth indexes in relation to fertilization
Concerning the influence of fertilization, in Mendoza (figure 5) the lower level of fertilization N1P1K1 (F1) showed significantly higher LAI values compared with the control (F0) on several sampling dates. N2P2K2 (F2) also produced higher results than F0 at 1052, 1362 and 1653 GDD. In Rovigo1 and Rovigo2 high fertilization doses F2 and F3 that differed by the higher N content (300 kg ha-1), produced larger LAI than the control on all sampling dates except the first one.
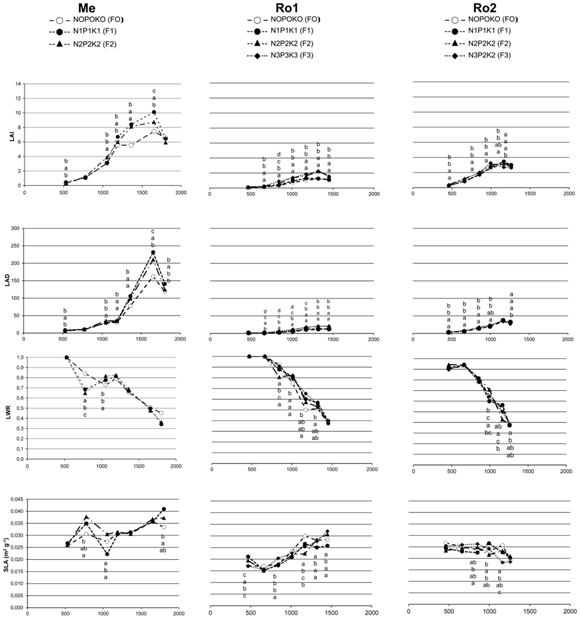
In each experiment, within growing degree days (GDD), means with the same letters are not significantly different at P ≤ 0.05 (Tukey HSD test). The letters position refers to the order of fertilization treatments reported in the figure.
En cada experimento, dentro del mismo grado día (GDD), medias con igual letra no son significativamente diferentes a P ≤ 0,05 (Tukey HSD test). La posición de las letras se refiere al orden de los tratamientos de fertilización reportados en la figura.
Figure 5. Fertilization influence on LAI, LAD, LWR and SLA in different years and environments (Me = Mendoza; Ro1 = Rovigo first year; Ro2 = Rovigo second year).
Figura 5. Influencia de la fertilización en LAI, LAD, LWR, y SLA en diferentes años y ambientes. ( Me = Mendoza; Ro1 = Rovigo primea año; Ro2 = Rovigo segundo año).
An increase in LAI duration (LAD), for most sampling dates in both locations and years, was observed for highly fertilized plots (figure 5). In particular, in Me, the higher values were obtained with F1. In Ro1, F2 and F3 significantly improved this index compared with F0 and F1 from 661 GDD to the end of growing cycle. In Ro2, instead, higher fertilization effectiveness with F2 and F3 were evident from the first sampling date until 1000 GDD. After 1257 GD), a substantial index decrease was determined by F3.
The ratio between photosynthetically active leaves and total plant leaves dry weight (LWR) was differently infuenced by fertilization formula. In Me fertilization affected this index just at 776 and 1052 GDD with a non-consistent trend. In Ro1, a beneficial effect of fertilization was found on just three dates (1010, 1172 and 1320 GDD). Similarly, the next year (Ro2) fertilization treatments showed different results only on the last three sampling dates and again with a non-consistent trend.
SLA (figure 5), that generally identifes leaf thickness, was less affected than the other growth indexes by fertilization doses during the growing cycle. In Me fertilization increased index values at 776 GDD as doses increased, but at the next sampling higher values were observed in F0 and F2 treatments and, at 1802 GDD, in F1. For all other GDD fertilization doses did not affect SLA. The results in Ro1 were very heterogeneous, with beneficial effects of fertilization just at 467 and 841 GDD. The next year (Ro2) fertilization only increased SLA on one date (993 GDD) compared to the control.
LAR, calculated as SLA x LWR (figure 6), showed a general decrease during the growing cycle, particularly marked in Ro2 and Me. In the latter location fertilization affected LAR only at 1052 GDD, with the highest value for F2. In Ro1 fertilization produced higher LAR at 467, 841 and 1172 GDD with higher values for F1 and F3 on different dates

In each experiment, within growing degree days (GDD), means with the same letters are not significantly different at P< 0.05 (Tukey HSD test). The letters position refers to the order of fertilization treatments reported in the figure.
En cada experimento, dentro del mismo grado día (GDD), medias con igual letra no son significativamente diferentes a P < 0,05 (Tukey HSD test). La posición de las letras se refiere al orden de los tratamientos de fertilización reportados en la figura.
Figure 6.Fertilization influence on LAR, NAR, RGR and CGR in different years and environments (Me = Mendoza; Ro1 = Rovigo first year; Ro2 = Rovigo second year).
Figura 6. Influencia de la fertilización en LAR, NAR, RGR, y CGR en diferentes años y ambientes. ( Me = Mendoza; Ro1 = Rovigo primea año; Ro2 = Rovigo segundo año).
Concerning dry matter per photosynthetically active leaf area unit and time unit (NAR), Me and Ro1 showed an increasing NAR until 1052 and 841 GDD, respectively (figure 6). This trend was not observed in Ro2. Regarding fertilization doses, Me showed higher values of NAR for F1 and F2 only at 1362 GDD. In Ro1 fertilization increased NAR on the first three dates compared to the control. However significant differences were obtained by different doses. After 1010 GDD the highest dose (F3) produced a significant decrease of the index with respect to the other treatments. In Ro2 fertilization treatments produced higher NAR values than the control on four of six dates. Again it is difficult to detect a defined pattern about which is the best fertilization formula since different formulae gave improvements with respect to the control on different dates.
The relative growth rate (RGR), calculated as the rate of increase of total dry weight per unit of plant dry weight, showed different trends in both locations and years (figure 6). In Me, F1 increases RGR relative to control at 529 and 1052 GDD, while at 776 and 1362 GDD F2 did the same. In Ro1, higher values of RGR were obtained by F2 at 467 GDD and by F3 at 661 GDD. At 1010 GDD, an RGR contraction was determined by F3 and, on the last sampling date, F2 and F3 drastically reduced the values. In Ro2 higher RGR were observed for fertilization treatments F2 and F3 at first sampling, but at 851 GDD and 1257 GDD the same fertilization treatments produced a decrease of RGR, indicating a non-consistent trend in the effect of fertilization on this index.
Crop growth rate (CGR), is shown in figure 6. CGR in Me was very high compared with Ro1 and Ro2. In Me, fertilization treatments, specially F1, produced higher values of CGR, but at the last sampling date (1802 GDD) fertilization drastically reduced the index. In Ro1 higher CGR were obtained by higher fertilization treatments (F2 and F3) on most dates. However, at the last sampling date (1451 GDD), a consistent contraction in values was determined by these treatments. A different trend was observed in Ro2, where F2 and F3 differed from other treatments only at 460 and 993 GDD. At 1163 GDD the higher value was found for F2, while the N content increase in F3 caused an index contraction. On the last sampling date F2 and F3 produced a similar decrease in CGR to Me and Ro1.
DISCUSSION
Concerning different environments we can see that crop growth rate (CGR) at Mendoza, calculated as the multiplication of photosynthetic plant effectiveness (NAR) and leaf area index (LAI) was strongly infuenced by NAR from transplanting but in the last part of the season it also seemed to be infuenced by the LAI increase. The increase in LAI may be due to favorable Argentinean environmental conditions (soil and climate). From 776 to 1190 GDD plants showed a strong growth rate with a 3-4 fold increase. In this period they formed leaves with high NAR (over 20% increase) and low thickness (over 20% decrease). From 1052 to 1653 GDD CGR decreased because the photosynthetic component of the index (NAR) decreases in parallel. At the beginning of this last part of the growing cycle head formation and enlarging started. This event is confirmed by some growth indexes trends that, at first, showed photosynthetic effectiveness (NAR) reduction and not proportional LAI expansion, with a consequent CGR decrease.
Thereafter they presented a continued and strong decrease of photosynthetically active leaves dry weight and those of total plant leaves ratio (LWR). But CGR of Me was still higher than Ro1 and Ro2 due to rise in values of LAI that occurred at that moment in Mendoza. They also displayed an obvious contraction of photosynthetic biomass and the rest of plant ratio (LAR) due to LWR. Moreover, LAR and NAR decrease led to an evident RGR contraction. The end of the growing cycle -which for this kind of radicchio takes place in autumn when heads are near marketable maturation and plant metabolic activities slow down- is also coincident with low temperatures during the season. Under these conditions it is not surprising that many growth indexes showed a decreasing trend. In particular, photosynthetic effectiveness reached very low values, causing similar behavior for RGR and CGR. Also SLA decreased and this event was probably due to high soil salinity from 0.0 to 0.4m deep. This result was also observed for Phaseolus vulgaris grown in high EC nutrient solution (2).
Considering fertilization treatments effectiveness on the mentioned growth indexes, no clear effects appeared although some trends have been observed. Fertilized plants, especially with F1, were able to intensely assimilate during the juvenile phase. After that moment fertilization treatments did not always differ during the growing cycle and, when they did, higher values appeared in an irregular way. At harvest high fertilization treatments caused outer leaves to fall earlier, as underlined by LAI and LAD values. In this last period, high fertilization also determined a strong CGR decrease due to photosynthetic effectiveness and LAI reduction. The unfertilized control showed a less drastic CGR decrease because of NAR reduction which was compensated, within certain limits, by LAI increase.
At Rovigo growth indexes values showed a not always similar trend in both trial years. In some cases there were evident differences, especially for LAR and SLA. CGR always showed higher values in 1994 and, at 841 and 1010 GDD, increases were 69.4 and 65.1% higher respectively, than 1993. This was almost exclusively due to higher LAI values, since NAR did not greatly differ between years. In the first year, lower values can be justifed by high EC of the circulating solution that, as reported for other species e. g. barley (15), sugar beet (18), water melon (3), bean (2) and some brassicaceae (9), can also determine drastic leaf area contraction.
In the second half of growing cycle CGR was instead strongly linked to photosynthetic effectiveness contraction due to head formation, which leaves are photosynthetically not active and in addition they are rich in anthocyannins increasing the metabolic cost of producing such kind of leaves.
Concerning the RGR trend, Ro1 and Ro2 were affected by photosynthetic biomass and the rest of plant ratio (LAR). In particular, an earlier and drastic LAR decrease occurred in Ro2 compared with Ro1, surely due to head formation. Leaf thickness (SLA) values showed a different trend, but this is justifed by the high salinity of soil circulating solution in 1993, as reported in materials and methods. Also in this environment there was no clear infuence of fertilization on growth indexes, but some results are worth commenting. F2 and F3 showed high growth rates, especially at the beginning of the growing cycle. Afterwards responses differed among treatments, but their effects were not continuous over sampling dates. This was probably due to the typical species heterogeneity that made representative plant sampling very difficult. At harvest, high fertilization anticipated outer leaves fall due to a lower tissues texture caused by high N content. This led to a precocious head marketable maturation.
Finally, considering yield aspects with relation to growth indexes, first of all it can be stated that the photosynthetically active leaves dry weight and total plant leaves ratio (LWR) is strongly linked with head marketable maturation; at this time LWR value was 0.38 g g-1 independently of location, trial year and growing cycle length, as verifed in Me. The low yield observed in Ro1, due, as already reported, to high soil EC, was also confirmed by some growth indexes like LAI, LAD and especially CGR. This last index was 75% lower at around 1000 GDD than in the other trials. LAI also confirmed plant responses during the growing cycle; indeed, in Me, index values, drastically higher than Ro1 and Ro2, were determined by higher photosynthetically active leaf number and size. In the former location maximum leaf number was 29 at 1362 GDD, while in the latter, this value did not exceed 20 leaves. The high yield in Ro2 was not due to high LAI, but it was justifed by leaf thickness increase (as indicated by SLA decrease), especially at the end of the growing cycle. Taking the average of the fertilizations, in fact, total and marketable plant weight were 626 and 357 g, respectively. This result is not verifed by the table 4 (page 119) data because this vegetable, new to the Argentinean environment, was not able to form marketable heads before temperatures froze (< 0 °C). In this condition, although harvesting lasted longer than 60 days, this chicory, characterized by high genetic heterogeneity, at the end of possible harvesting time still presented many plants in the rosette phase.
CONCLUSIONS
The results obtained in this experiment allow some conclusions to be drawn on growth indexes in relation to fertilization and yield aspects in radicchio. LWR allowed a critical value (0.35-0.40 g g-1) to be obtained for determining harvest time and this can be useful in areas where radicchio is introduced for the first time. It was also noticed that, in suboptimal environmental conditions, LAI reaches high values and this highlights a growing cycle extension to the detriment of compact head formation.
Concerning fertilizations with increasing doses of macronutrients, highly fertilized plants seemed to assimilate more intensely, especially during the juvenile phase, which was more or less extended in relation to the weather pattern. Sampling performed in the middle of the growing cycle gave results that were difficult to explain and did not allow particular effects to be attributed to fertilization treatments. At harvest, carried out at marketable head maturation, an earlier fall of outer leaves was observed with higher fertilization. This was basically due to high nitrogen content that determined a lower tissues texture. It should be stressed that, with this chicory, these considerations are valid only for this cultivar and precocity class.
1. Andrade, F. H. 1995. Analysis of growth and yield of maize, sunfower and soybean grown at Balcarce, Argentina. Field Crops Research, 41: 1-12. [ Links ]
2. Bayuelo-Jiménez, J. S.; Debouck, D. G.; Lynch, J. P. 2003. Growth, gas exchange, water relations, and ion composition of Phaseolus species grown under saline conditions. Field Crops Research, 80: 207-222. [ Links ]
3. Chartzoulakis, K. S. 1994. Photosynthesis, water relation and leaf growth of cucumber exposed to salt stress. Sci. Hort., 59: 27-35. [ Links ]
4. Gabrielle, B.; Denoroy, P.; Gosse, G.; Justes, E.; Andersen, M. N. 1998. A model of leaf area development and senescence for winter oilseed rape. Field Crops Research, 57: 209-222. [ Links ]
5. Gianquinto, G. 1997. Morphological and physiological aspects of phase transition in radicchio (Cichorium intybus L. var silvestre Bischoff): infuence of daylength and its interaction with low temperature. Sci. Hort., 71: 13-26. [ Links ]
6. Gianquinto, G.; Pimpini, F. 1995. Morphological and physiological aspects of phase transition in radicchio (Cichorium intybus L. var silvestre Bischoff): the infuence of temperature. Adv. Hort. Sci., 4: 192-199. [ Links ]
7. Gouyi, F. L. L. 1993. The characteristics and constraints of dry matter production in broccoli. Journal of Northwest Sci-Tech University of Agriculture and Forestry, 2: 11. [ Links ]
8. Graifenberg, A.; Giustiniani, L.; Temperini, O. 1987. Crescita e asportazione degli elementi nutritivi nel pomodoro da industria. Colture Protette, 3: 51-55. [ Links ]
9. He, T.; Cramer, G. R. 1993. Growth and ion accumulation of two rapid-cycling Brassica species differing salt tolerance. Plant and Soil, 153(1): 19-31. [ Links ]
10. Holsteijn, H. M. C. van. 1980. Growth of lettuce. II. Quantitative analysis of growth. Mededelingen Landbouwhogeschool Wageningen, 80(13): 1-24. [ Links ]
11. Hunt, R. 1990. Basic growth analysis - Plant Growth Analysis for Beginners. Unwin Hyman, London. [ Links ]
12. Innocenti, M.; Gallori, S.; Giaccherini, C.; Ieri, F.; Vincieri, F. F. ; Mulinacci, N. 2005. Evaluation of the phenolic content in the aerial parts of different varieties of Cichorium intybus L. J. Agric. Food Chem., 53: 6497-6502. [ Links ]
13. Lavelli, V. 2009. Antioxidant activity of minimally processed red chicory (Cichorium intybus L.) evaluated in xanthine oxidase-myeloperoxidase-and diaphorase-catalyzed reactions. J. Agric. Food Chem., 56: 7194-7200. [ Links ]
14. Maddonni, G. A.; Otegui, M. E. 1996. Leaf area, light interception, and crop development in maize. Field Crops Research, 48: 81-87. [ Links ]
15. Munns, R.; Greenway, H.; Delane, R.; Gibbs, J. 1982. Ion concentration and carbohydrate status of elongation leaves tissue of Hordeum vulgare growing at high external NaCl. II. Cause of growth reduction. J. Exp. Bot., 33: 574-583. [ Links ]
16. Nicoletto, C.; Pimpini, F. 2009. Infuence of the forcing process on some qualitative aspects in radicchio "Rosso di Treviso tardivo" (Cichorium intybus L., group rubifolium). 1. Nitrate, nitrite and organic nitrogen. Ital. J. Agron. / Riv. Agron., 4: 137-145. [ Links ]
17. Nicoletto, C.; Pimpini, F. 2010. Infuence of the forcing process on some qualitative aspects in radicchio "Rosso di Treviso tardivo" (Cichorium intybus L., group rubifolium). 2. Antioxidant capacity, phenols and ascorbic acid. Ital. J. Agron. / Riv. Agron., 5: 43-2. [ Links ]
18. Papp, J. C.; Ball, M.; Terry; N. 1983. A comparative study of the effects of NaCl salinity on respiration, photosynthesis and leaf extension growth in Beta vulgaris. Plant Cell. Environ., 6: 675-677. [ Links ]
19. Pimpini, F.; Filippini, M. F.; Sambo, P.; Gianquinto, G. 2000. Infuenza della concimazione sul contenuto di nitrato in due tipi di radicchio rosso. Riv. Agron., 34: 406-418. [ Links ]
20. Pimpini, F.; Nicoletto, C.; Varotto, S. 2007. Radicchio: efficacia del fotoperiodo e della vernalizzazione sull'induzione a fore. Colture Protette, 1: 50-63. [ Links ]
21. Pimpini, F.; Nicoletto, C. 2008. Effetto della vernalizzazione sull'induzione a fore in tipologie e classi diverse di radicchio (Cichorium intybus L. var silvestre Bischoff). Ital. J. Agron. / Riv. Agron., 2 Suppl.: 69-80. [ Links ]
22. Poli, F.; Sacchetti, G.; Tosi, B.; Fogagnolo, M.; Chillemi, G.; Lazzarin, R.; Bruni, A. 2002. Variation in the content of the main guaianolides and sugars Cichorium intybus var. "Rosso di Chioggia" selections during cultivation. Food Chemistry, 76: 139:147. [ Links ]
23. Rogers, H. H.; Cure, J. D.; Smith, J. M. 1986. Soybean growth and yield response to elevated carbon dioxide. Agriculture, Ecosystems and Environment, 16: 113-128. [ Links ]
24. Rossetto, M.; Lante, A.; Vanzani, P.; Spettoli, P.; Scarpa, M.; Rigo, A. 2005. Red cichories as potent scavengers of highly reactive radicals: a study on their phenolic composition and peroxyl radical trapping capacity and efficiency. J. Agric. Food Chem., 53: 8169-8175. [ Links ]
25. Santamaria, P.; Ventrella, D.; Magnifico, V.; De Boni, A.; Serio, F. 1993. Accrescimento, produzione e accumulo dei nitrati in spinacio (Spinacia oleracea L.) concimato con fertilizzante minerale. Riv. Agron., 27(4): 587-591. [ Links ]
26. Setiyono, T. D.; Weiss, A.; Specht, J. E.; Cassman, K. G.; Dobermann, A. 2008. Leaf area index simulation in soybean grown under near-optimal conditions. Field Crops Research, 108: 82-92. [ Links ]
27. Vega, J.; Lis, B. R. de; Cavagnaro, J. B. 1972. Rusticación a sequía en cebolla. II. Análisis del crecimiento. Phyton, 30(1/2): 89-107. [ Links ]
28. Yoshida, R.; Watanabe, S.; Fukuta, Y.; Kusaka, K.; Iwai, K.; Tamaka, T. 2003. Effect of 5-aminolevulinic acid on growth and nutrient uptake of leaf vegetables in alkaline soil. Proceedings of 30th annual meeting of Plant Growth Regulation Society of America. August 3-6, Vancouver, British Columbia, Canada. 30: 142-143. [ Links ]

