










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo
On-line version ISSN 1853-8665
Rev. Fac. Cienc. Agrar., Univ. Nac. Cuyo vol.45 no.2 Mendoza Dec. 2013
ARTÍCULOS ORIGINALES
Effect of colchicine and amiprophos-methyl on the production of in vitro doubled haploid onion plants and correlation assessment between ploidy level and stomatal size
Efecto de la colchicina y del amiprofos-metil en la producción in vitro de plantas dihaploides de cebolla y determinación de la correlación entre el nivel de ploidía y tamaño de los estomas
María Laura Foschi 1, Liliana Estela Martínez 2, MaríaTeresa Ponce 2, Claudio Rómulo Galmarini 1, 3, Borut Bohanec 4
1 Chair of Horticulture and Floriculture. Agricultural Faculty. National University of Cuyo. Alte. Brown 500. Chacras de Coria, Mendoza. Argentina. M5528AHB. mfoschi@fca.uncu.edu.ar
2 Chair of Plant Physiology. Agricultural Faculty. National University of Cuyo. Instituto de Biología Agrícola de Mendoza-Consejo Nacional de Investigaciones Científicas y Técnicas (IBAM-CONICET). Alte. Brown 500, Chacras de Coria, Mendoza. Argentina. M5528AHB.
3 Estación Experimental Agropecuaria INTA La Consulta, Mendoza, Argentina - Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
4 Biotechnical Faculty. University of Ljubljana, Jamnikarjeva 101, 1000 Ljubljana, Slovenia.
Originales: Recepción: 23/10/2012 - Aceptación: 19/08/2013
Abstract
Doubled haploid onion (Allium cepa L.) plants allow the production of completely homozygous lines for a later production of hybrids. The haploid plants are normally produced using in vitro gynogenesis. The obtained haploid plantlets must be treated with different agents for doubling chromosomes. It is necessary to adjust the concentration and the length of treatment of the doubling agent. In this case, the effect of 250 and 500 mg.L-1 colchicine and 15.2; 30 and 60 mg.L- 1 amiprophos-methyl during 24 and 48 h was assessed over the rate of onion haploid plantlets chromosome doubling. The best duplication treatment was 250 mg.L-1 colchicine for 48 h, which yielded 100% of doubled haploid plants. On the other hand, a positive correlation resulted from the ploidy level and stomatal size, and a negative correlation between the level of ploidy and stomatal density. Significant differences between the stomatal length, width and density in haploid and doubled haploid plantlets were observed. An economical and quick method to test ploidy level in onion plantlets is proposed through the measurement of stomatal size and density.
Keywords: Allium cepa L.; Antimitotic agents; Duplication; Gynogenesis; Hybrids
Resumen
La producción de plantas dihaploides de cebolla (Allium cepa L.), permite obtener líneas completamente homocigotas para luego producir híbridos. A través de la ginogénesis in vitro se ha logrado producir plantas haploides, las cuales deben ser sometidas a tratamientos de duplicación de sus cromosomas, siendo necesario ajustar la concentración del agente duplicador y la duración del tratamiento. En este trabajo se evaluó el efecto de la aplicación de 250 y 500 mg.L-1 de colchicina y 15,2; 30 y 60 mg.L-1 de amiprofos-metil durante 24 y 48 h, sobre la tasa de duplicación de cromosomas en plántulas haploides de cebolla. El mejor tratamiento de duplicación fue 250 mg.L- 1 de colchicina, durante 48 h, logrando un 100% de plantas dihaploides. Por otra parte, se obtuvo una correlación positiva entre el nivel de ploidía y el tamaño de estomas y una correlación negativa entre el nivel de ploidía y la densidad estomática, registrándose diferencias significativas entre los valores promedio de largo, ancho y densidad de estomas para plantas haploides y dihaploides. Se propone un método económico y rápido para verificar el nivel de ploidía de las plántulas de cebolla mediante la medición del tamaño y densidad estomática.
Palabras clave: Allium cepa L.; Agentes antimitóticos; Dduplicación; Ginogénesis; Híbridos
Abbreviations
APM: amiprophos-methyl
DAPI: 4',6-diamidino-2- phenylindole
DH: doubled haploid
DMSO: dimethyl sulfoxide
MS: Murashige and Skoog
Introduction
The development of homozygous lines is a fundamental step in the production of onion hybrids. However, one of the drawbacks of conventional improvement methods for this species is the prolonged time required for obtaining pure lines through self- pollination. In onion, more than three inbreeding generations cannot be done without high depression levels due to consanguinity. As a result, only partial homozygous lines can be obtained (1). The production of doubled haploid (DH) plants is a strategy used in a large number of species (17, 18, 26). It allows the production of complete homozygosis plants in a short period of time.
In onion, the production of DH is achieved through haploids induction via gynogenesis (in vitro cultivation of flowers, ovaries or non-pollinated ovaries) and followed by doubling the chromosomes number resulting in highly homogenous individuals with a high degree of genetic stability (2, 3, 12, 13). The production of complete homozygous plants in a shorter period constitutes a critical stage and is affected by: explant type, doubling substance, treatment length and treatment conditions (34). The most widely used doubling agent either in onions or other species is colchicine (3, 4, 14, 20), with an average doubling efficiency of 70% (21, 24, 25, 32). Less frequently, and with variable efficiency levels, trifluralin, oryzalin and amiprophosmethyl (APM) have been also used (4, 16, 22, 29).
Several studies about genome doubling in various species have been carried out. A protocol needs to be developed for 'Cobriza INTA' cultivar, there are several methods described in the literature for onion, but not for this cultivar. Another critical and laborious step in obtaining DH is the determination of ploidy level (8). The most frequently used methods have been chromosome counts in meristem root tips and the use of flow cytometry. The direct measurement of ploidy level has the disadvantage of being very laborious and time-consuming (10, 11, 30). Flow cytometry, allows the direct measurement of DNA quantity in nuclei from fresh leaves. Although it is a quick and hardly destructive technique, it measures nuclei in phases of high and low ploidy levels in a single plant; it also requires extremely costly equipment (9).
Measuring stomatal length, stomatal density and the number of chloroplasts in guard cells are indirect ways of determining ploidy levels. Such techniques are a quick, successful and harmless form of measurement techniques (6, 7) based on a highly positive correlation between the ploidy level and the stomatal length, and a highly negative correlation between the ploidy level and the stomatal density (23, 28, 37, 38, 39). These correlations have not been yet assessed in onions. In this work, an adjustment of an efficient method for chromosome number doubling in onion plantlets obtained via in vitro gynogenesis is proposed by using colchicine and APM. Also, it is proposed the use of measuring stomatal length and density for determining ploidy levels.
Materials and methods
Doubling conditions determination
Plant material
A total of 480 explants of 'Cobriza INTA' onion (19) from in vitro multiplication of a gynogenetic haploid embryo, according to the technique described by Kahane et al. (27), were used.
Doubling treatments
The effect of colchicine (Sigma) in two concentrations: 250 and 500 mg.L-1 (625 and 1250 μM), and APM (Fluka) in three concentrations 15.2 mg.L-1 30 mg.L-1 and 60 mg.L-1 (50, 100, and 200 μM) were tested in 2 exposition periods: 24 and 48 h. Nine replications were used for colchicine treatments, and 5 replications for APM treatments. An experimental unit of 6 plants per replication was tested.
The culture media was constituted by half strength macro and micronutrients of Murashige and Skoog (31) (MS), without growth regulators, adding 40 g.L-1 sucrose, jelled with 7.5 g.L-1 Bacto-Difco-Agar™ (Becton Dickinson S. R. L., Argentina) and adjusted to pH 5.8 (30, 34, 36). Colchicine was dissolved in water, whereas APM was dissolved in dimethyl sulfoxide (DMSO). Both substances were added to the sterile culture medium before solidification, by previously filtering with 0.22 μM Millipore filters.
Plantlets were cut alongside in half to be exposed to antimitotic agents (figures 1A, 1B, pág. 159). During and after treatment, the plantlets were exposed to a photoperiod of 16 h of 70 μmol m-2 h-1 light and to a temperature of 26 + 2°C. Once the treatments finished, the explants were placed into the same culture media, without growth regulators.

Figure 1. Conditioning of haploid plantlets in order to be exposed to antimitotic substances. Plantlet divided in half (A). Haploid plantlets on the solid culture medium during doubling treatments (B).
Figura 1. Acondicionamiento de las plántulas haploides para ser expuestas a las sustancias antimitóticas. Plantas divididas a la mitad (A). Plántulas haploides en medio de cultivo sólido durante el tratamiento de duplicación (B).
Ploidy analysis
The ploidy level was assessed two months after the doubling treatments were done, in order to assure that all of the budding leaves were originated from the cells treated with the doubling substances. All the samples were analyzed by flow cytometry (Model Partec PA, sofware Partec GMBH, Germany), belonging to the Institute of Floriculture - INTA, Castelar, Buenos Aires, Argentina, following Otto's methodology (33).
Assessed variables
In each treatment, the following variables were assessed: number of vitrescent plants (hyperhydric plants), survival percentage, the ploidy level of the plantlets and doubling efficiency (product between survival percentage and duplicate proportion of plantlets).
Experimental design and statistical analysis
The jars containing the plants were randomly distributed in culture chambers. The percentage of doubled plants without phytotoxicity symptoms was compared among treatments through the Chi-square test and proportions comparison was carried out for 95 % significance level (Statgraphics 4.0).
Correlations between ploidy level and stomatal size
Plant material
A total of 5 haploid genotypes and 5 diploid genotypes from 'Cobriza INTA' cultivar were used. The haploid plantlets were obtained through in vitro culture of unpollinated onion flowers, following the technique suggested by Ponce et al. (35). The diploid plantlets came from nectaries, placed at the bottom of the flowers, cultured in vitro and from dihaploid plantlets obtained through genome doubling treatments with antimitotic substances. The culture media was constituted by half strength macro and micronutrients of Murashige and Skoog (31), without growth regulators, adding 40 g.L-1 sucrose, jelled with 7.5 g.L-1 Bacto-Difco-Agar™ (Becton Dickinson S. R. L., Argentina) and pH 5.8. The plantlets were exposed to a photoperiod of 16 hours (70 μmol. m-2. h-1) and 26 + 2°C.
Ploidy level verification
The ploidy level of each plantlet was verified through chromosome counts in root meristems and also by flow cytometry. For direct chromosome countigs, the root tips were stained following Feulgen's technique; which consists in immersing root tips in a 0.1% colchicine solution, then the material was fixed in a 3:1 ethanol: glacial acetic acid solution, followed by the disgregation of proteins in HCl 1 N and rinse with 45% acetic acid. Finally, the material was stained with 1% hematoxylin in order to make microscope observations.
For flow cytometry, nuclear DNA content was stained with DAPI according to procedure modified by Otto (33). Nuclei of sample and standard species were released in 0.1 M citric acid containing 0.5% Tween 20. The suspension was filtered through a 30 μm nylon-mesh filter. A 3-4 volume of staining buffer containing 4 μg/ml DAPI in 0.4 M disodium hydrogen phosphate was added. Measurements were done on a Partec PAS flow cytometer using a linear scale. Samples were analyzed using an UV spectrum excited with a HBO lamp and emissions were measured through a GG 435 long-pass filter.
Stomatal size and density measurement
The length and width of 100 closed occlusive cells were measured on the epidermis of three completely developed leaves on in vitro plantlets. The stomatal density was calculated from the number of stomata per field. Fifteen fields were measured per plant. The number of stomata per visual field was then converted to stomata per mm2. In order to remove foliar epidermis, the 'peeling' technique described by D'Ambrogio de Argüeso (15) was used.
The images were captured using a ZEISS (Axioplan, West Germany) video camera connected to a microscope and a computer. The observations were made with a 20 x 1.6 zoom. The measurements of stomatal length, width and number per field were carried out with AxioVision 3.0 image software (figures 2A, 2B).
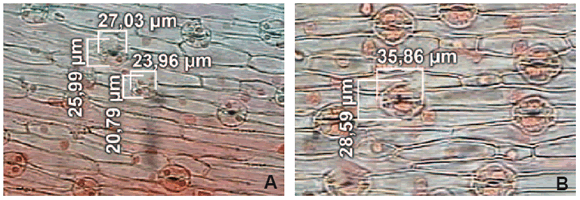
Figure 2. Measurement of stomatal length and width with AxioVision microscope in in vitro onion plantlets of 'Cobriza INTA' cultivar. Haploid (A) and diploid plantlets (B).
Figura 2. Medidas del largo y ancho de los estomas realizadas con microscopio AxioVision en plántulas in vitro del cultivar 'Cobriza INTA'. Plántulas haploides (A) y plántulas diploides (B).
Statistical analysis
Stomatal size, length, width and density were assessed. T-test was used to compare samples with different variances. Also, normal reliability intervals were used for ploidy levels (95% significance level). Correlation analysis between stomatal length, width and density and the ploidy level were carried out for 95% significance level (Statgraphics 4.0).
Results and discussion
Doubling experiment
Those plants treated with 250 mg.L-1 and 500 mg. L-1 colchicine for 48 h showed 42.6 and 44.4% vitrescence percentages, respectively (table 1).
Table 1. Percentage of vitrecence, survival and ploidy level in haploid plantlets of cv. 'Cobriza INTA' under the effect of different colchicine treatments.
Tabla 1. Porcentaje de vitrescencia, sobrevivencia y nivel de ploidía en plántulas haploides de cebolla cv. 'Cobriza INTA' con diferentes tratamientos de colchicina.

Vitrescence is a physiological disorder which, on certain occasions, appears on plants under in vitro conditions. This disorder has multiple causes, such as culture conditions, genotype, and presence of some growth regulators or other chemical substances like colchicine and APM (8). Similarly, all treatments with APM showed significantly superior vitrescence levels (23.3% to 30%) than that of the control (3.3%) (table 2). Nevertheless, the high vitrescence levels finally did not influence the efficiency of the treatment since many of these plants improved their condition and budded once they were transferred to a fresh media without antimitotics agents.
Table 2. Percentage of vitrecence, survival and ploidy level in haploid plantlets of cv. 'Cobriza INTA' under the effect of different APM treatments.
Tabla 2. Porcentaje de vitrescencia, sobrevivencia y nivel de ploidía en plántulas haploides de cebolla cv. 'Cobriza INTA' con diferentes tratamientos de APM.

The use of 500 mg.L-1 colchicine for 24 h showed the lowest doubling efficiency (12.9). In the remaining treatments, such variable diminished between 24.1 and 70.4 respect to the control. Treatment with 500 mg.L-1 colchicine for 24 h exposition, induced a high rate of mixoploids (65.4%), followed by treatment 250 mg. L-1 - 24 h (5.4%). In the remaining treatments statistically significant differences were not observed (table 1, page 160). The rate of mixoploids obtained from the use of APM ranged from 22.2% to 60.9 %. These proportions may indicate that the concentration was not enough. Although an increase of such concentration could raise the doubling rate, it would also increase phytotoxicity, diminishing the total efficiency of the treatment.
The variability of responses shown in the bibliography, as that obtained in this work, highlight the great dependence of the doubling efficacy on the genotype, thus justifying the need to determine the most adequade treatment in each case.
An inverse relationship between the doubling percentage and survival was recorded. Therefore, by using 500 mg.L-1 colchicine for 48 h, 100% doubling and the lowest survival rate were achieved (24.1%). By diminishing the concentration at 250 mg.L-1 with the same exposition duration, the doubling percentage was maintained, while the survival rate increased up to 70.4% (table 1, page 160). Regarding those treatments in which APM was used, the best one was 15.2 mg.L-1 for 48 h with 57.7% doubling and 70 % plantlet survival (table 2, page 160).
By comparing both antimitotic substances, the treatment with colchicine, 250 mg.L-1 during 48 h, proved to be the most doubling efficiency (70.4), due to the achievement of the highest doubling rate (100%) at the same survival percentage as the treatment with APM (15.2 mg.L-1 during 48 h), in which only 57.7% doubling was achieved and doubling efficiency was 40.4.
The doubling efficiency obtained with colchicine in this work is the highest one recorded for onion up to the present (3, 4, 21, 32). Concerning the use of APM, the doubling percentages obtained were similar to, and in most cases, higher than those reported in the bibliography (4, 21, 24, 26), but lower than those obtained with colchicine.
Correlation determination between ploidy level and stomatal size
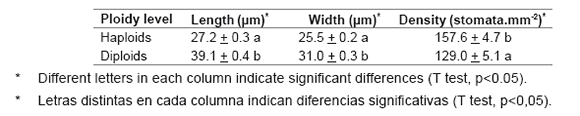
Differences in the stomatal length and density between haploid and diploid plants cultured in vitro were detected (P<0.05). The haploid plants had lower stomatal length and width, compared to diploid plants. The stomatal length and width of haploid plants were, 27.2 + 0.3 μm and 25.5 + 0.2 μm, respectively, compared to the diploid ones, which showed 39.1 + 0.4 μm and 31.0 + 0.3 μm respectively. In the same way, the number of stomata per mm2 also showed significant differences between haploid and diploid plants, being higher in haploid ones (table 3, page 162).
Table 3. Comparison of stomatal length, width (μm) and density (stomata.mm-2) among haploid and diploid 'Cobriza INTA' plantlets.
Tabla 3. Comparación del largo y ancho de estomas estomas (μm) y densidad (estomas. mm-2) entre plántulas haploides y diploides de cebolla cv. 'Cobriza INTA'.
These results agree with those obtained by several authors in other species with plants coming from in vitro culture as well as from field crops. Zhang et al. (40) assessed the stomatal length in in vitro cultured phlox plants and found a larger number of stomata in tetraploid plants than in diploid ones. Aryavand et al. (5) also concluded that stomatal size and number can be used to distinguish tetraploid plants from hexaploid ones in Aegilops neglecta. In other species, such as Vitis vinifera L. and wheat, significant differences were obtained in polyploid and diploid plants (28, 39).
A positive correlation (P<0.05), between the stomatal size and ploidy level was observed, which indicate that stomatal length (correlation coefficient: 0.62) and width (correlation coefficient: 0.41) can be directly related to the increase of ploidy level. On the other hand, a negative correlation between stomatal density and ploidy level (correlation coefficient: -0.25) was recorded, indicating that a decrease in stomatal number per mm2 resulted in an increase in ploidy level.
Based on these results, confidence intervals were carried out to determine ploidy level by stomatal density and size for a 95 % significance level. The ranges in which one invitro onion plant can be considered haploid or diploid are the following: for haploid plant: length (26.6 - 27.8 μm), width (24.9 - 25.9 μm), density (148.3 - 166.9 stomata. mm- 2) and for diploid plant: length (38.3 - 39.8 μm), width (30.4 - 31.5 μm) and density (118.8 - 139.2 stomata.mm-2). The stomatal measurement can be a useful and economic tool for ploidy level determination in onion.
Conclusions
The low percentage of spontaneous replication and stability of onion haploid plants confirms the need for an efficient protocol genome duplication using antimitotic substances. Generally, genome duplication of haploid plantlets increases with increased concentration and duration of treatment. In this paper the use of 250 mg.L-1 of colchicine for 48 h, in solid medium, was the most efficient as it produced the highest rate of genome duplication onion plantlets, consistent with the reduced phytotoxicity and high doubling efficiency.
By the results obtained, it can say that it is possible to determine the ploidy level of haploid and diploid in vitro onion plantlets, through stomata size and density. This confirms that the measurement of the stomata can be very important to determine different ploidy levels in most species. Therefore, it is proposed to measure the length, width and / or density of stomata onion plants from in vitro cultivation, as an alternative to determine ploidy important since it is a rapid, simple, inexpensive and effective procedure.
1. Acosta, A.; Gaviola, J.; Galmarini, C. R. 1993. Manual de producción de semillas hortícolas. In: Crnko, J. (ed.). La Consulta. Asoc. Coop. EEA La Consulta INTA. 83 p.
2. Alan, A.; Mutschler, M.; Brants, A.; Cobb, E.; Earle, E. 2003. Production of gynogenic plants from hybrids of Allium cepa L. and A. Roylei Stearn. Plant Sci. 165: 1201-1211. [ Links ]
3. Alan, A.; Brants, A.; Cobb, E.; Goldschmied, P.; Mutschler, M.; Earle, E. 2004. Fecund gynogenic lines from onion (Allium cepa L.) breeding materials. Plant Sci. 167:1055-1066. [ Links ]
4. Alan, A.; Mutschler, M.; Wansang, L.; Earle, E. 2007. Complementary strategies for ploidy manipulations in gynogenic onion (Allium cepa L.). Plant Sci. 173:21-35. [ Links ]
5. Aryavand, A.; Ehdaie, B.; Tran, B.; Waine, J. 2003. Stomatal frecuency and size differentiate ploidy levels in Aegilops neglecta. Genet. Res. Crop. Evol. 50: 175-182. [ Links ]
6. Beck, S.; Dunlop, W.; Fossey, A. 2003. Stomatal lenght and frecuency as a measure of ploidy level in black wattle, Acacia mearnsii (de Wild). Bot. J. Linn. Soc. 141:177-181. [ Links ]
7. Beck, S.; Visser, G.; Dunlop, R. 2005. A comparison of direct (flow cytometry) and indirect (stomatal guard cell lengths and chloroplast numbers) techniques as a measure of ploidy in black wattle, Acacia mearnsii (de Wild). South Afr. J. Bo. 71: 354-358. [ Links ]
8. Bohanec, B. 2002. Doubled-haploid onions. In: Rabinowitch, H.; Currah, L. (ed.). Allium Crop Science: Recent Advances. CABI Pub Wallingford Oxon New York. p. 145-157.
9. Bohanec, B. 2003. Ploidy determination using flow cytometry. In: Maluszynski, M.; Kasha, K.; Forster, B.; Szarejko, I. (ed.). Doubled haploid production in crop plants. Kluwer Academic Press, Dordrecht Boston London. p. 397- 403.
10. Bohanec, B.; Jackse, M.; Ihan, A.; Javornik, B. 1995. Studies of gynogenesis in onion (Allium cepa L.): induction procedures and genetic analysis of regenerants. Plant Sci. 104: 215-224. [ Links ]
11. Campion, B.; Alloni, C. 1990. Induction of haploid plants in onion (Allium cepa L.) by in vitro culture of unpollinated ovules. Plant Cell Tissue Organ Cult. 20: 1-6. [ Links ]
12. Campion, B.; Azzimonti, M.; Vicini, E.; Schiavi, M.; Falavigna, A. 1992. Advances in haploid plant induction in onion (Allium cepa L.) through in vitro gynogenesis. Plant Sci. 86: 97-104. [ Links ]
13. Campion, B.; Schiavi, M. 1994. Production of doubled haploid lines of onion (Allium cepa L.): Progress report and problems. In: Proceedings of the International Colloquium IPBA. December 5-7. Rogla Slovenia, p. 25-33.
14. Campion, B.; Azzimonti, M.; Perri, E.; Schiavi, M.; Vicini, E. 1995. Spontaneus and induced chromosome doubling in gynogenic lines of onion (Allium cepa L.). Plant Breed. 114:243-246. [ Links ]
15. D'Ambrogio de Argüeso, A. 1986. Manual de técnicas en histología vegetal. Buenos Aires. Editorial Hemisferio Sur S. A. 54 p.
16. Dhooghe, E.; Grunewald, W.; Leus, L.; Van Labeke, M. 2009. In vitro polyploidisation of Helleborus species. Euphytica 1: 89-95. [ Links ]
17. Forster, B.; Heberle-Bors, E.; Ken Kasha, J.; Touraev, A. 2007. The resurgence of haploids in higher Plants. Trends in Plant Sci. 12:368-375. [ Links ]
18. Foschi, M. L.; Martínez, L. E.; Ponce, M. T.; Galmarini, C. R. 2009. Doubled haploids, a biotechnologic strategy for onion (Allium cepa L.) genetic breeding. Avances en Horticultura, Review 66: 40-48. [ Links ]
19. Galmarini, C. R. 2000. Onions cultivars released by La Consulta experimental station, INTA, Argentina HortScience 35: 1360-1362.
20. Geoffriau, E.; Kahane, R.; Bellamy, C.; Rancillac, M. 1997. Ploidy stability and in vitro chromosome doubling in gynogenic clones of onion (Allium cepa L.). Plant Sci. 122:201-208. [ Links ]
21. Grzebelus, E.; Adamus, A. 2004. Effect of anti-mitotic agents on development and genome doubling of gynogenic onion (Allium cepa L.) embrions. Plant Sci. 69: 1-6. [ Links ]
22. Hansen, A.; Gertz, L.; Joersbo, M.; Andersen, S. 2000. Chromosome doubling in vitro amiprophos methyl in Beta vulgaris ovule culture. Acta Agr. Scand. 50: 89-95.
23. Inceer, H.; Hayirlioglu-Ayaz, S. 2010. Chromosome numbers in Tripleurospermum Sch. Bip. (Asteraceae) and closely related genera: relationships between ploidy level and stomatal length. Plant Systematics Evol. 285: 149-157.
24. Jakse, M.; Bohanec, B. 2000. Studies of alternative approaches for genome doubling in onion. Biotechnological Approaches for Utilization of Gametic Cells COST 824: Final Meeting. Bled, Slovenia. p. 101-104.
25. Jakse, M.; Bohanec, B. 2003. Haploid induction in onion via gynogenesis. In: Maluszynski, M.; Kasha, K.; Forster, B.; Szarejko, I. (ed.). Doubled haploid production in crop plants. Kluwer Academic Press, Dordrecht Boston London. p. 281- 285.
26. Jakse, M.; Havey, M.; Bohanec, B. 2003. Chromosome doubling procedures of onion (Allium cepa L.) gynogenic embryos. Plant cell Rep. 21: 905-910.
27. Kahane, R.; Rancillac, M.; Teyssendier de la Serve, B. 1992. Long- term multiplication of onion (Allium cepa L.) by cyclic shoot regeneration in vitro. Plant Cell Tissue Organ Cult. 28: 281-288. [ Links ]
28. Khazaei, H.; Monneveux, P.; Hongbo, S. ; Mohammady, S. 2010. Variation for stomatal characteristics and water use efficiency among diploid, tetraploid and hexaploid Iranian wheat landraces. Genet. Res. Crop Evol. 57: 307-314. [ Links ]
29. Lim, W.; Earle, E. 2009. Enhanced recovery of doubled haploid lines from parthenogenetic plants of melon (Cucumis melo L.). Plant Cell Tissue Organ Cult. 98: 351-356.
30. Martínez, L. E. 2003. In vitro gynogenesis induction and doubled haploid production in onion (Allium cepa L.). In: Maluszynski, M.; Kasha, K.; Forster, B.; Szarejko, I. (ed.). Doubled haploid production in crop plants. Kluwer Academic Press, Dordrecht Boston London. p. 276-279.
31. Murashige, T.; Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Plant Physiol. 15: 473-497.
32. Nowak, E. 2000. Gynogenic onions plants-studies on regeneration and diploidization. Biotechnological Approaches for Utilization of Gametic Cells COST 824: Final Meeting, Bled Slovenia, p. 95-99.
33. Otto, F. 1990. DAPI staining of fixed cells for high-resolution flow cytometry of nuclear DNA In: Darzynkiewickz, Z.; Crissman, H. A. (ed.). Methods in cell biology. Academic Press San Diego. p.105-110.
34. Ponce, M. T. 2007. Ginogénesis en cebolla. Avances en Horticultura, 5: 1-1.
35. Ponce, M. T.; Martínez, L. E.; Galmarini, C. R. 2006. Influence of CCC, putrescine and gellamgum concentration on gynogenic embryo induction in Allium cepa. Biol. Plant. 50:425-428.
36. Ponce, M. T.; Guiñazú, M. E.; Cirrincione, M.; Videla, M. E.; Arancibia, C. 2011. Micropropagación de la enredadera nativa Mutisia subspinosa Cav. Rev. FCA UNCUYO. 43(2): 235-243.
37. Rayburn, A.; Crawford, J.; Charlotte, M.; Rayburn, C. M.; Juvik, J. A. 2009. Genome size of three Miscanthus species. Plant Mol. Biol. Rep. 27: 184-188.
38. Rodríguez Portela de Carvalho, J. F.; Carvalho, C.; Otoni, W. C. 2005. In vitro induction of polyploidy in annatto (Bixa orellana). Plant Cell Tissue Organ Cult. 80:69-75. [ Links ]
39. Yang, X. M.; Cao, Z.; An, L.; Wang, Y.; Fang, X. 2006. In vitro tetraploid induction via colchicine treatment from diploid somatic embryos in grapevine (Vitis vinifera L.). Euphytica 152: 217-224. [ Links ]
40. Zhang, Z.; Dai, H.; Xiao, M.; Liu, X. 2008. In vitro induction of tetraploids in Phlox subulata L. Euphytica 159:59-65. [ Links ]
Acknowledgments
To SECTYP (Secretaría de Ciencia, Técnica y Posgrado de la Universidad Nacional de Cuyo), and INTA (Instituto Nacional de Tecnología Agropecuaria) for supporting this work.

