










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo
On-line version ISSN 1853-8665
Rev. Fac. Cienc. Agrar., Univ. Nac. Cuyo vol.47 no.1 Mendoza June 2015
ARTICULO ORIGINAL
Gene action associated with heterosis expression in scarlet eggplant (Solanum gilo Raddi.)
Acción gen asociado con la expresión de heterosis en Jiló (Solanum gilo Raddi.)
Thiago Matos Andrade 1 , André Lasmar 2 , Wilson Roberto Maluf 2 , Luiz Antonio Augusto Gomes 2 , Ranoel José de Sousa Gonçalves 3 , Arie Fitzgerald Blank 1
1 Department of Agronomic Engineering, Federal University of Sergipe, Av. Marechal Rondon s/n, São Cristóvão, Sergipe, Brazil. tmaagro@yahoo.com.br
2 Department of Agriculture, Federal University of Lavras - Depto Agricultura, Post Office Box 3037, CEP. 37200-000, Lavras, Minas Gerais, Brazil.
3 Academic Unit of Technology, Federal University of Campina Grande (UFCG), CEP 58540-000, Sumé, Paraíba, Brazil.
Originales: Recepción:07/02/2014 - Aceptación:27/11/2014
Abstract
The objectives of this work were to assess heterosis and its components in F 1 hybrids of a diallel cross among lines of scarlet eggplants (Solanum gilo Raddi, syn. S. aeth opicum L.), and to study the modes of gene action associated with its expression. Six lines (Morro Redondo, Branco Comprido, BGH-1544, Tinguá, Irajá, Comprido Verde Claro) and 15 hybrids resulting from a diallel cross among them (reciprocals excluded) were tested in a randomized complete block design with four replications. Traits under study were total yield, mean fruit mass and fruit length/diameter ratio. There was no evidence of epistatic gene action in the expression of any of the traits under study. Heterosis for yield could be explained by overdominance, but higher yields were only weakly associated with larger frequencies of dominant alleles an indication that yields may be markedly influenced by the action of relatively few loci with overdominant gene action. Heterosis for mean fruit mass was associated with incompletely dominant gene action, and larger fruit mass was associated with larger proportions of dominant alleles. For fruit length/diameter ration (L.D -1 ), heterosis was due to incompletely dominant gene action, and dominant alleles were predominantly associated with lower L.D-1 values.
Keywords: Solanum gilo Raddi; Dominance; Epistasis; Overdominance
Resumen
Los objetivos de este trabajo fueron evaluar la heterosis y sus componentes en los híbridos F1 de una dialélico entre líneas de jiló (Solanum gilo Raddi, syn. S. aethiopicum L.), y el estudio de los modos de acción de los genes asociados con su expresión. Seis líneas (Morro Redondo, Branco Comprido, BGH-1544, Tinguá, Irajá, Comprido Verde Claro) y 15 híbridos resultantes de un cruce dialélico entre ellos (recíprocos excluídos) se pusieron a prueba en un diseño de bloques completamente aleatorio con cuatro repeticiones. Los rasgos estudiados fueron producción total, peso promedio del fruto y la relación longitud/diámetro. No hubo evidencia de la acción de genes epistatic en la expresión de cualquiera de los rasgos en estudio. Para la producción total, la heterosis se explica por la acción de genes overdominance, ya que había una asociación débil, no significativa, entre los alelos dominantes y los valores de producción de frutas altas totales. La heterosis de masa media de la fruta se asoció con la acción del gen dominante incompleto, y la masa da fruta más grande se asoció con una mayor proporción de alelos dominantes. Para la relación longitud/diámetro (LD -1 ), la heterosis fue debida a la acción del gen dominante incompleto, y alelos dominantes se asociaron principalmente con menores valores LD-1 .
Palabras clave: Solanum gilo Raddi; Dominación; Epistasis; Overdominance
INTRODUCTION
The scarlet eggplant (Solanum gilo Raddi, syn. S. aethiopicum L. gilo group) is a little known vegetable crop akin to the widely known Asian eggplant (S. melongena) (7). Its characteristically bitter tasting fruit is consumed immature, as is that of eggplant, but it is usually much smaller than that of the latter. It is a species of African origin, very popular in states of Southeastern and Central Brazil, where it was introduced in colonial times by African slaves. In recent years, it became known in places where the population of Brazilian ethnic residents is significant, such as the state of Massachusetts in the USA, where a local production is now available (Mangan et al. 2010).
Cultivars grown in Brazil comprise basically two horticultural types: one with elongated light-green immature fruit (which is preferred in the Brazilian states of Minas Gerais, Rio de Janeiro, and Goiás), and another with roundish dark-green immature fruit (preferred in the state of São Paulo) (4). In both cases, mature fruit color is red. Other immature fruit colors, including white and green-netted, are also available, but have minor economic importance.
The use of F 1 hybrids is widespread for eggplants S. melongena, due to reported high degrees of heterosis (6, 13). For S. gilo, however, all cultivars grown in Brazil are open pollinated, in spite of indications that heterosis in scarlet eggplant is as conspicuous as it is for its S. melongena counterpart (2, 3, 9, 12). Because autogamy is the predominant breeding system of scarlet eggplant (2), it is assumed that the majority of S. gilo cultivars are either inbred lines or mixture of closely related inbred lines. Only a few reports are available to support the use of commercial F 1 hybrids. Campos (1973) studied the performance of eight lines and their respective F 1 hybrids in a diallel cross, and reported a high degree of heterosis for yield-related traits. Carvalho & Ribeiro (2002) reported that both general (GCA) and specific (SCA) combining abilities were significant for fruit shape, fruit number, fruit yield, fruit length, fruit diameter and mean fruit mass, among other traits an indication of dominat and/or epistatic effects involved.
Monteiro (2009) reported high degrees of heterosis expression in S. gilo hybrids, whether heterosis be measured relatively to midparental values, or relatively to the superior parental line, or even to a standard open-pollinated cultivar - a situation that should make the deployment of commercial F 1 hybrids very convenient. The study made no attempt, however, to study the components of heterosis, or the types of gene action involved in its expression.
Diallel analyses in S. gilo should provide a quick way to assess the potential for deployment of F 1 hybrids in the species. The diallel analysis proposed by Jinks & Hayman, (1953); Miranda et al. (1982), provides for a detailed study of the types of gene action involved in the trait expression. Alternatively, the diallel analysis proposed by Gardner & Eberhart (1966) provides for a detailed measure of the varietal effects and of heterosis and its components in the hybrids constituent of the diallel cross.
The objectives of this work were to assess heterosis and its components in F 1 hybrids of a diallel cross among lines of scarlet eggplant (Solanum gilo Raddi), and to study the modes of gene action associated with its expression.
MATERIALS AND METHODS
Six S. gilo lines (1=Morro Redondo; 2=Branco Comprido; 3=BGH-1544; 4=Tinguá; 5=Irajá; 6=Comprido Verde Claro) were crossed via manual emasculation and pollinations, in order to obtain all possible 15 hybrid combinations (no distinction was made between reciprocal hybrids). Morro Redondo has roundish dark green fruit, and is considered a standard cultivar in the state of São Paulo. Tinguá, Irajá and Comprido Verde Claro bear elongated light green fruit, and are the major cultivars in the Brazilian states of Minas Gerais, Rio de Janeiro and Goiás. BGH-1544 is an accession from the germplasm bank of the Universidade Federal de Viçosa, Viçosa-MG, Brazil, and also bears elongated light green fruit. Branco Comprido is an accession with elongated white skinned fruit. The six lines and their 15 hybrids were tested in a randomized complete block trial with four replications, which was carried out at the Vegetable Research Station of Horti Agro Sementes Ltda, Ijaci-MG-Brasil (21°14'16" S, 45°08'00'' W, altitude 920 m), from June 2008 through January 2009. Seeds were sown on June 17, 2008 in 128-cell speedling trays filled (35 mL.cell -1 ) with a commercial substrate (Geneplus®). Seedlings were transplanted on August 27, 2008, with 7 plants per plot, spaced 1.20 m between rows x 0.70 m between plants within a row. Standard cultural practices were deployed (4), and the trial was drip-irrigated.
Fruit harvests started 70 days after transplanting date, and were effected over a period of 60 days, totaling 10 harvests. The last harvest was on January 12, 2009.
The following traits were evaluated: total yield mean fruit mass and fruit length/diameter (L.D -1 ) ratio. Yield was measured as the sum of the recorded yields in all 10 harvest dates, and was expressed as metric tons per hectare (t.ha - 1 ). Mean fruit mass (g.fruit -1 ) and fruit L.D -1 ratio were measured by sampling 35 fruits per plot during the second harvest date.
Data were subject to analyses of variance, and means were separated by the Tukey test at a = 0.05. Diallel analyses were performed according to the procedures indicated by Gardner & Eberhart (1966) and by Jinks & Hayman (1953).
Results and discussion
Significant differences (a = 0.05) were detected among treatments for all traits under study for both the varietal and heterosis effects (table 1).
Table 1. Analyses of variance for total yield, mean fruit mass, and fruit length/diameter (L.D -1 ) ratio in a diallel cross among Solanum gilo lines.
Tabla 1. Análisis de varianza de la producción total, peso promedio del fruto y la relación longitud/diámetro en dialélico cruz de Solanum gilo Raddi.

Varietal effects and mean heterosis comprise the major portion of the variation (table 1, and table 2, page 23).
Table 2.Estimates of mean components according to the analysis of the diallel cross among Solanum gilo lines.
Tabla 2.Las estimaciones de los componentes de medias de acuerdo con el análisis de la cruz dialélico entre líneas Solanum gilo.
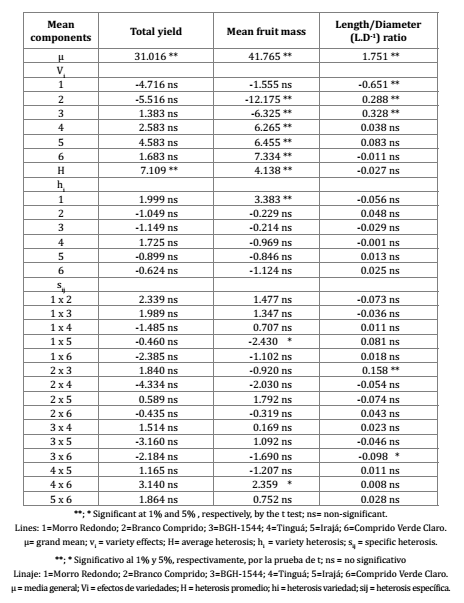
In the model proposed by Jinks & Hayman (1953), an estimate of the regression coefficient ("ß") of Wr (covariances) on Vr (variance among the parents) that differs from 1 indicates the presence of epistatic gene action; otherwise, if ß does not differ from 1, absence of epistasis is assumed. Accordingly, there was no evidence for epistasis in the expression of any of the three traits under study (table 3, page 24).
Table 3.Regressions (ß) of Wr on Vr, and correlations between (Wr+Vr) and Yr for total yield, mean fruit mass, and fruit length/diameter ratio, according to the diallel analysis.
Tabla 3.Regresiones (ß) de Wr sobre Vr, y correlaciones entre (Wr + Vr) y Yr de producción total, peso promedio del fruto y la relación longitud/diámetro, según el análisis dialélico.

Total fruit yield
The genotypes differed from each other in yield (table 1, page 22 and table 4, page 25).
Table 4.Means of the Solanum gilo genotypes, and percent heterosis in the hybrids, for total yield, mean fruit mass, and fruit length/diameter ratio.
Tabla 4. Medios de los genotipos de Solanum gilo, y el porcentaje de heterosis en los híbridos para la producción total, peso promedio del fruto y la relación longitud/diámetro.
Heterosis for yield was always positive, with values varying from 5.7 to 40.4% relatively to the midparental means. Hybrids with a moderate degree of heterosis, associated with high yielding parents, showed the best mean yields, as was the case of hybrids 3x4, 4x5, 4x6 and 5x6, with increases of 18.1, 26.4, 34.3 and 21.9% relative to midparent, respectively (table 4, page 25).
Yield was influenced largely by the varietal (v i ) and average heterosis (H) components (table 1, page 22 and table 2, page 23). Average heterosis (H) was significant and positive, indicating a general yield superiority of the hybrids when compared to midparental means (table 2, page 23). The varietal (h i ) and specific (s ij ) heterosis components were not significant.
The results therefore indicate that hybrid yields can be predicted by the sum of their midparental means with a value common to all hybrids - the average heterosis component (H).
In the Jinks-Hayman analysis, the regression coefficient of Wr on Vr was estimated as ß = 0.606 (table 3, page 24), a value that differed from zero (a =0.05) but not from 1, therefore the additivedominant model was adequate to explain yield expression, with no evidence for epistatic gene interactions. Overdominant gene action was detected, because of the intercept of the Wr-Vr regression line on the vertical axis is a negative value (figure 1, page 26).

Figure 1.Regression of Wr on Vr, and limiting parabola, for total yield in Solanum gilo.
Figura 1.Regresión de Wr en Vr y parábola limitante, para la producción total en Solanum gilo.
The value (-0.273) of the regression of (Wr+Vr) on Yr (= yield of the rth parent) (table 3, page 24) indicated a weak non-significant association between alleles with dominance and higher total fruit yields in S. gilo.
The highest yielding parent (parent 5) lay close to the lower portion of the regression line (figure 1), indicating that it is the parental line with higher proportion of dominant alleles; however, parents 4 and 6 are also high yielding, but were those with highest proportion of recessive genes, a result that reinforces the weak association of high yield with higher frequencies of dominant alleles. This weak association may be an indication that yields are markedly influenced by the action of relatively few loci with overdominant gene action.
Mean fruit mass
The parental lines differed among each other in mean fruit mass (table 1, page 22 and table 4, page 25). Line 2 had the lowest fruit mass (< 30 g. fruit -1 ), contrasting with lines 4, 5 and 6 (> 48 g fruit -1 ). Three hybrids (1x4, 4x6 and 5x6) had mean fruit mass of > 51 g.fruit -1 (table 4, page 25). Midparent heterosis was positive for all hybrids (table 4, page 25), with highest values, +25% and +23%, for hybrids 1x2 and 1x3, respectively. For the remaining hybrids, heterosis values varied from +2% to +16%.
Lower performance parents can generate hybrids with high degrees of heterosis for this trait, as was the case with 1x2 and 1x3. Hybrid 4x6, though not highly heterotic (+9%), was nonetheless the hybrid with highest fruit mass (table 4, page 25).
Mean fruit mass was largely influenced by the varietal (v i ) and average heterosis (H) components (table 1, page 22 and table 2, page 23), and in a few cases only, also by varietal (h i ) and specific (s ij ) heterosis components (table 1, page 22 and table 2, page 23).
Fruit mass in the hybrids would therefore be a value close, but not necessarily identical, to the sum of of the midparental mean and the average heterosis component (H).
The existence of significant varietal heterosis and specific heterosis effects cannot be disregarded in a few cases (table 2, page 23), but even in these cases they were lower in magnitude than the average heterosis effect (H).
The regression of Wr on Vr (ß = 0.714) differed from zero (a =0.05) but not from unity, indicating absence of epistatic gene action and adequacy of the additivedominant model. The intercept of the straight line (figure 2) is a positive value, indicating incomplete dominance of the genes affecting the trait.

Figure 2.Regression of Wr on Vr, and limiting parabola, for mean fruit mass in Solanum gilo.
Figura 2.Regresión de Wr en Vr y parábola limitante, para el peso promedio del fruto en Solanum gilo.
The correlation between (Wr + Vr) and Yr (r = -0.564) (table 3, page 24) is a significant value (a =0.05), indicating that dominant genes act predominantly in the direction of increased fruit mass. The parental line 2, which presented the lowest fruit mass, lay in the upper region of the regression line (figure 2, page 27), an indication of presence of a higher proportion of recessive alleles; on the other hand, parents 1, 4, 5 and 6, with heavier fruit, lay in intermediary position or in the lower region of the regression line (figure 2, page 27), reinforcing the association between dominant alleles and higher fruit mass.
Fruit length/diameter ratio (L.D -1 )
There were significant differences in L.D -1 among the genotypes (table 1, page 22 and table 4, page 25). L.D -1 varies according to fruit shape: round or roundish fruit have L.D -1 close to 1, while elongated fruit had L.D -1>1 and usually close to 2. The sole parent with roundish fruit (line 1) had L.D -1 = 1.1 (table 4, page 25), whereas lines 2 and 3, with the most elongated fruit, had L.D-1>2. None of the hybrids had round or roundish fruit: hybrids with line 1 as a parent had L.D-1 that varied from 1.37 to 1.46.
The remaining hybrids had L.D -1 values between 1.77 and 1.98 (table 4, page 25).
Heterosis values were negative for the majority of the hybrids (table 4, page 25), even though their magnitudes were low. In the six cases (1x5, 2x3, 2x5, 2x6, 4x6 e 5x6) in which it was not positive, it was a value close to zero. L.D -1 ratios were largely determined by varietal (vi ) and average heterosis (H) effects, and in a few cases only, also by varietal heterosis (h i ) and specific heterosis (s ij ) effects (table 1, page 22 and table 2, page 23).
Results therefore indicate that hybrid means for L.D -1 would be close to midparent, but the low-magnitude negative average heterosis (H) effect points out a tendency for L.D -1 ratios slightly lower than those of midparental means (table 1, page 22 and table 2, page 23).
There were, however, hybrids whose average heterosis and specific heterosis effects were significant, and larger in magnitude than the average heterosis (table 2, page 23); these hybrids may therefore diverge from the general rule of L.D -1 means lower than midparental means.
The regression of Wr on Vr (ß = 0.830) (table 3, page 24) differed from zero (a =0.05), but not from 1.
Therefore there was no evidence for epistatic gene action, and the additivedominant model was adequate to explain L.D -1 expression.
The correlation between (Wr + Vr) and Yr (r = -0.848) (table 3, page 24) indicated that dominant alleles act predominantly in the direction of lower L.D -1 values, i.e., towards rounder or less alongated fruit.
The line 1, with the lowest L.D -1 ratio, lay in the lower portion of the regression line (figure 3, page 29), indicating a higher frequency of dominant alleles, whereas lines 2 and 3 presented higher L.D -1 (>2.0), lay in the upper portion of the regression line (figure 3, page 29), and possess higher frequencies of recessive alleles.

Figure 3.Regression of Wr on Vr, and limiting parabola, for fruit length/diameter ratio in Solanum gilo.
Figura 3.Regresión de Wr en Vr y parábola limitante, para la relación longitud/ diámetro del fruto en Solanum gilo.
This reinforces the conclusion that dominant alleles are primarily responsible to determine round or roundish. Gene action is likely incomplete dominance, because the intercept of the regression line is a positive value (figure 3, page 29).
Only a few reports are available to S. gilo.
Campos (1973) studied the performance of eight lines and their respective F 1 hybrids in a diallel cross, and reported a high degree of heterosis for yield-related traits. Carvalho & Ribeiro (2002) reported that both general (GCA) and specific (SCA) combining abilities were significant for fruit yield, fruit diameter and mean fruit mass among other traits an indication of dominat and/or epistatic effects involved. For the characteristics studied in the present work, we observed the effect of dominacia and / or overdominance, showing what was reported by Carvalho & Ribeiro (2002).
Monteiro (2009) reported high degrees of heterosis expression in S. gilo hybrids, whether heterosis be measured relatively to midparental values, or relatively to the superior parental line, or even to a standard open-pollinated cultivar - a situation that should make the deployment of commercial F1 hybrids very convenient.
The diallel analysis proposed by Jinks & Hayman, (1953); Miranda et al., (1982) and Gardner & Eberhart (1966) allowed a detailed study of the types of gene action involved in the trait expression as well as a detailed measure of the varietal effects and of heterosis and its components in the hybrids in the diallel cross.
CONCLUSIONS
Heterosis was significant for yield, fruit mass or L.D -1 ratio in scarlet eggplant, Epistatic gene action was not important in the expression of heterosis for yield, fruit mass or L.D -1 ratio. Dominant effects were important in the expression of all three traits under study. Heterosis for yield was of higher magnitude than heterosis for mean fruit mass or for fruit length/ diameter ratio.
Heterosis for yield was largely due to overdominant gene action. There was only a weak non significant association between the frequency of alleles with dominance and larger yield values, an indication that yields may be markedly influenced by the action of relatively few loci with overminant gene action.
Heterosis for mean fruit mass and L.D-1 ratio was associated with incompletely dominant gene action. Larger fruit mass and lower L.D -1 ratios were associated with higher frequency of alleles with dominance for these traits.
1. Campos, J. P. 1973. Aspectos teóricos e aplicados da heterose em jiló (Solanum gilo Raddi). Piracicaba: ESALQ. 88 p. (Tese doutorado). [ Links ]
2. Campos, J. P.; Blumenschein, A.; Casali, V. W. D; Rodrigues, J. J. V. 1979. Avaliação de oito cultivares de jiló (Solanum gilo Raddi) e suas progênies híbridas F1. Revista de Olericultura. 17: 304-213. [ Links ]
3. Carvalho, A. C. P. P.; Ribeiro, R. L. D. 2002. Análise da capacidade combinatória em cruzamentos dialélicos de três cultivares de jiloeiro. Horticultura Brasileira. 20: 48-51. [ Links ]
4. Filgueira, F. A. R. 2003. Novo Manual de Olericultura - "Agrotecnologia moderna na produção e comercialização de hortaliças", Editora UFV, Viçosa, Brazil. 402 p. [ Links ]
5. Gardner, C. O.; Eberhart, S. A. 1966. Analysis and interpretation of variety cross diallel and related populations. Biometrics. 22: 439-452. [ Links ]
6. Ikuta, H. 1969. Melhoramento e genética da berinjela, In: KERR, W. E. (ed,), Melhoramento e genética.São Paulo. Melhoramentos. 9: 161-168. [ Links ]
7. Isshiki, S.; Taura, T. 2003. Fertility restoration of hybrids between Solanum melongena L, and S, aethiopicum L. Gilo Group by chromosome doubling and cytoplasmic effect on pollen fertility. Euphytica. 134: 195-201. [ Links ]
8. Jinks, J. L.; Hayman B. I. 1953. The analysis of diallel crosses. Maize Genetics Corporation News Letter. 27: 48-54. [ Links ]
9. Lester, R. N.; Thitai, G. N. W. 1989. Inheritance in Solanum aethiopicum, the scarlet eggplant. Euphytica. 40(1/2): 67-74. [ Links ]
10. Mangan, F.; Moreira, M., Martuscelli, T. Produtividade e comercialização de sementes à população de falantes de português em Massachusets. University of Massachusets. Amherst. Available In: http://www,umassvegetable,org/growers_services/pdf_files/ portuguese,pdf e www,umassvegetable,org/pdf_files/portuguese_crops_english,pdf, (Acessed Jan 03, 2010). [ Links ]
11. Miranda, J. E. C.; Maluf, W. R.; Campos, J. P. 1982. Análise genética de um cruzamento dialélico de cultivares de tomate - II Características vegetativas. Pesquisa Agropecuária Brasileira. 17: 767-773. [ Links ]
12. Monteiro, A. B. 2009. Obtenção de híbridos e análise da heterose em jiló (Solanum gilo Raddi), UFLA, Lavras, MG, Brazil. 44. (M,S, Dissertation, in Portuguese). [ Links ]
13. Sousa, J. A.; Maluf, W. R.; Gomes, L. A. A. 1997. Produtividade e qualidade de frutos de cultivares de polinização aberta e híbridos F1 de berinjela (Solanum melongena L,). Ciência e Agrotecnologia. 21(3): 334-342. [ Links ]
ACKNOWLEDGEMENTS
The authors wish to thank the following Brazilian institutions that provided support for this research project: FAPEMIG- Fundação de Amparo à Pesquisa do Estado de Minas Gerais, CNPqConselho Nacional de Desenvolvimento Científico e Tecnológico, FINEP/ MCT- Ministério de Ciência e Tecnologia/ FNDCT, CAPES/MEC-Brazilian Ministry of Education, UFLA- Universidade Federal de Lavras, HortiAgro Sementes Ltda,, FUNDECC- Fundação para o Desenvolvimento Científico e Tecnológico, FAEPE-Fundação de Apoio ao Ensino, Pesquisa e Extensão.

