










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo
Print version ISSN 1853-8665On-line version ISSN 1853-8665
Rev. Fac. Cienc. Agrar., Univ. Nac. Cuyo vol.50 no.2 Mendoza Dec. 2018
ORIGINAL ARTICLE
Planting density and its effect on plant height and rosette quality in ornamental cabbages for cutting
Efecto de la densidad de plantación sobre altura de planta y calidad de roseta en repollos ornamentales de corte
Yoo Ree Kim1, Libertad Mascarini1, Gabriel Antonio Lorenzo1, Mariel Norma González1
1 Universidad de Buenos Aires. Facultad de Agronomía. Departamento Producción Vegetal. Cátedra de Floricultura. Avda. San Martín 4453. (1417) Buenos Aires. Argentina. lmascari@agro.uba.ar
Originales: Recepción: 17/02/2017 - Aceptación: 22/08/2017
ABSTRACT
Brassica oleracea L. var. acephala is an ornamental cabbage cultivated as foliage for cutting due to the coloration of the rosette formed by its apical leaves. The plant height and the diameter of the rosettes may be modified through the planting density; however, there is not enough information about this. The aim of this study was to assess the influence of the planting density of four cultivars of B. oleracea L. var. acephala of the series F1 Crane: Bicolor, Pink, Red and Rose, on the plant height, rosette diameter, number of leaves, leaf area, root volume, and aerial and root fresh and dry weight. The experimental design was completely randomized, with six repetitions. The combination of density (56 plants.m-2 and 28 plants.m-2) and cultivar allowed obtaining eight treatments. The results showed that the treatments with high planting density improved commercial quality indices: achieving greater stem height and more compact and smaller diameter rosettes, leading to lower values of the rest of the vegetative variables evaluated. Also, as productive alternative, it would allow a more efficient use of the soil and reduce crop management activities such as manual leaves removal.
Keywords: Brassica oleracea L.; Foliage for cutting; Market quality
RESUMEN
Brassica oleracea L. var. acephala se cultiva como follaje para corte debido a la coloración de la roseta de hojas apicales de este repollo ornamental. Es posible modificar la altura de las plantas y el diámetro de las rosetas, para mejorar la calidad comercial, a través de la densidad de plantación, sin embargo, no hay suficiente información al respecto. Se evaluó la influencia de la densidad de plantación en cuatro cultivares de la serie F1 Crane: Bicolor, Pink, Red y Rose, sobre altura del tallo, diámetro de roseta, número de hojas, área foliar, volumen radical y peso fresco y seco, particionados en vástago y raíz. El diseño experimental fue completamente aleatorizado (DCA), con seis repeticiones. Por combinación de factores densidad (56 pl.m-2 y 28 pl.m-2) y cultivar se obtuvieron 8 tratamientos. Una alta densidad de plantación aumentó la calidad comercial de las plantas, las que tuvieron rosetas apicales más compactas y de menor diámetro y mayor altura del tallo, reduciendo el resto de las variables vegetativas. Asimismo, permitiría un uso más eficiente de la tierra disminuyendo tareas de manejo del cultivo tales como el deshoje de las varas.
Palabras clave: Brassica oleracea L.; Follaje para corte; Calidad comercial
INTRODUCTION
Brassica oleracea L. var. acephala is a biennial plant usually grown as an annual for ornamental purposes. Ornamental cabbages for cutting are used as substitutes for flowers during autumn and winter due to the decorative aspect of their central leaves, that display different colors, which are enhanced by the degradation of chlorophylls induced by low temperatures (13). Unlike that observed in other varieties, the central leaves of B. oleracea L. var. acephala develop a rosette-like shape, without forming a 'head' at the top of the plant (11). At the time of harvest, the lower leaves are removed, leaving the rosette on the top of the plant to give it a flower-like appearance.
The commercial production of ornamental cabbages usually takes place by the end of the summer, when warm temperatures promote the growth of plants. Growth stops when night temperatures are below 15°C, time at which the central and upper leaves express their characteristic coloration. An important management tool for the commercial production of ornamental cabbages is the choice of the number of plants per unit area because several factors such as the absorption of light, the consumption of water and nutrients, and the possible incidence of diseases depend on this variable.
According to different researchers, in order to satisfy the needs of the market, the size and composition of edible broccoli may be modified by different fertilizer (26) but ornamental cabbages for cutting may be modified through the management of the planting density. However, there is still not enough information on this issue. Thus, the aim of the present study was to assess the differences in plant height and rosette diameter, which are ornamental quality indexes, as well as in other vegetative variables in four cultivars of the series F1 Crane of B. oleracea L. var. acephala under two planting densities. A higher density of plants leads to cabbages of greater height and smaller rosette diameter, and reduces the individual growth of cabbages, affecting other components such as leaf area, number of leaves, root volume, and aerial and root fresh and dry weight.
MATERIALS AND METHODS
The experiment was carried out in the experimental field of the Cátedra de Floricultura of the Faculty of Agriculture of Buenos Aires University (34°40' S, 58°30' W). Four cultivars of B. oleracea L. var. acephala of the series F1 Crane (American Takii Inc.) were used: Bicolor, Red, Rose and Pink. Cultivation began on February 3 rd, 2012, starting from seedlings from the propagation company, on black plastic plug trays (150 cells of 28 cm3 each), with a substrate mix of 50:50 peat and perlite. The experiment was carried out in a greenhouse under controlled conditions for 14 days, time at which plants were transplanted to the field and sown in 0.8 m x 6.3 m flowerbeds raised 0.2 m above the ground.
For each cultivar, two treatments with different planting densities were carried out: high density (56 plants m-2) and low density (28 plants m-2), with a plantating diagram of 12.5 x 15.0 cm and 25 x 30 cm respectively. It was used a tutoring mesh (grid of 15 cm x 12.5 cm) that was raised as plants grew. Drip irrigation was managed according to the needs of the crop, and the necessary phytosanitary controls were carried out. Harvest took place when the upper leaves formed a colored center, characteristic of each cultivar. Data of the mean daily temperature during the experimental period were obtained from the Central Observatory of Buenos Aires (34°35'24"S 58°29'0" W).
The crop variables evaluated were: i) rosette diameter, determined at harvest as the average value of two perpendicular measures; ii) plant height, recorded on a weekly basis throughout the study with a metric tape from the soil surface up to the apex of the stem; iii) leaf area per plant, determined at harvest time and estimated by separating and scanning the leaves of each plant; the images were analyzed using ImageJ software; iv) number of leaves per plant, counted twice a week throughout the study, taking as a criterion the fully expanded leaf; v) root volume per plant, determined at harvest time by dipping the root (separated from the stem) in a test tube with a known volume of water (100 mL) and subsequently measuring the volume of the displaced water; vi) rate of leaf appearance, calculated from the cumulative number of new leaves per plant as a function of the elapsed time; vii) aerial fresh weight (AFW), considering the stem and the "rosette" of leaves, composed of two cycles of green leaves (eight leaves) taken from the colored center; viii) root fresh weight (RFW), determined by cutting the roots taking as a point of reference the first node located above their insertion; and ix) aerial and root dry weight (ADW and RDW respectively), determined by separating the stems and the leaves from the root and placing them in a stove at 70°C until consistency of weight. A TH-200 digital scale (accuracy 0.1 g) was used for all weight measurements. The measurements of all the studied variables were completed on April 23 rd, 2012.
The design was fully randomized with a 2 x 4 (density and cultivar) factorial arrangement, with the plant as experimental unit and six repetitions per treatment. The statistical analysis was performed using ANOVA and Tukey's Test (p = 0.05), using the Infostat software InfoStat / Professional V1.1 (8).
RESULTS
The evolution of the mean temperature during the experimental period presented a slightly descending trend, according to the change of season (summer-autumn). Coloration in the central leaves began to appear once mean temperatures showed a significant decrease (figure 1).
Figure 1. Evolution of the mean temperature (°C) during the crop cycle (summer-autumn 2012).
Figura 1. Evolución de la temperatura media durante el ciclo del cultivo (verano-otoño 2012).
This change was verified simultaneously in all the cultivars and showed to be gradually intensified in the subsequent days.
Regarding the plant height, the curves of the eight treatments showed a similar pattern of response, but with different magnitudes, which began to be obvious several weeks after planting.
The cv. Bicolor showed the greatest increases in plant height, mainly under the condition of high density, and, in the case of low density, in a shorter interval (with significant differences (p≤0.05) respect to the cv. Pink and Rose). At the end of the experiment, the plants cultivated at high density showed significant differences with respect to those planted at low density in all cultivars, except in Pink (figure 2, page 53).

Figure 2. Mean final height (cm) in ornamental cabbage plants (Brassica oleracea L. var. acephala) cv. Rose (Rs), Red (Rd), Pink (Pk) and Bicolor (Bc) at high planting density (HD) (56 plants/m2) and low planting density (LD) (28 plants/m2). The final height is the mean value ± standard error.
Figura 2. Altura final promedio (cm) (valor medio ± error estándar) en plantas de repollo ornamental (Brassica oleracea L. var. acephala) de los cv. Rose (Rs), Red (Rd), Pink (Pk) y Bicolor (Bc) en los tratamientos con plantación a alta densidad (AD) 56 plantas/m2 y baja densidad (BD) 28 plantas/m2.
All together, the plants grown at high density presented a final mean height that was 18.4% higher than that of plants grown at low density, with significant differences between them.
A significant density x cultivar interaction for plant height was found only for the first fifteen days from planting, whereas in the successive periods, the effects of both variables were independent (table 1, page 54).
Table 1. Mean final height (cm) of the plant as a function of the planting density and cultivar in B. oleracea L. var. acephala.
Tabla 1. Valores medios de altura final (cm) de la planta en función de los factores densidad y cultivar en repollos de B. oleracea L. var. acephala.
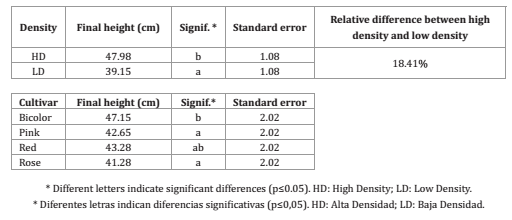
Rosette diameter was significantly affected by planting density and cultivar, but not by the interaction between these factors. This variable was smaller in plants sown at high density than in those sown at low density, with significant differences in all cultivars (figure 3, page 54).

Figure 3. Comparison of rosette diameters (cm) in plants of ornamental cabbage (Brassica oleracea L. var. acephala) cv Rose (Rs), Red (Rd), Pink (Pk) and Bicolor (Bc) at high planting density (HD) (56 plants/m2) and low planting density (LD) (28 plants/m2). Rosette diameter is the mean value ± standard error.
Figura 3. Comparación de diámetros de roseta (cm) (media ± error estándar de la media) en plantas de repollo ornamental (Brassica oleracea L. var. acephala) de los cv Rose (Rs), Red (Rd), Pink (Pk) y Bicolor (Bc) en los tratamientos con plantación a alta densidad (AD) 56 plantas/m2 y baja densidad (BD) 28 plantas/m2.
The mean rosette diameter of all the plants grown at high density was 32.7% smaller than that of those grown at low density, with significant differences (p≤0.05).
On the other hand, without considering the effect of density, it was found differences between cultivars, where the greatest mean rosette diameter was that of Bicolor, which was significantly different from that of Pink (p≤0.05). Both the planting density and the cultivar showed a significant effect on this variable, whereas the interaction of both factors did not.
The leaf area per plant presented significant differences between densities as well as between cultivars, but no density x cultivar interactions were found.
The mean leaf area developed by each plant showed different responses between treatments. In all cases, a lower planting density led to a greater leaf area (figure 4).

Figure 4. Comparison of the mean leaf area (m2/plant) of ornamental cabbage plants (Brassica oleracea L. var. acephala) (mean ± standard error of the mean) cv. Rose (Rs), Red (Rd), Pink (Pk) and Bicolor (Bc) at high planting density (HD) (56 plants/m2) and low planting density (LD) (28 plants/m2).
Figura 4. Comparación de medias de área foliar (m2/planta) en plantas de repollo ornamental (Brassica oleracea L. var. acephala) (media ± error estándar de la media) de los cv. Rose (Rs), Red (Rd), Pink (Pk) y Bicolor (Bc) en los tratamientos con plantación a alta densidad (AD) 56 plantas/m2 y baja densidad (BD) 28 plantas/m2.
The mean of the four cultivars showed that the plants grown at low density showed a significant increase in leaf area of 33.2% (p≤0.05). Regardless of planting density, this variable showed differences between cultivars.
The highest leaf area values were achieved by Red and Bicolor, which, on average, outperformed Pink and Rose with significant differences (p≤0.05).
The rate of leaf appearance showed a progressive increase as a function of temperature. This relationship was tighter in the treatments at high density (figure 5, page 56).
Figure 5. Rate of leaf appearance (leaves/day) in ornamental cabbage plants (Brassica oleraceaL. var.acephala) as a function of the mean daily temperature for the cultivars (A) Rose (Rs), (B) Red (Rd), (C) Pink (Pk) and (D) Bicolor (Bc) at high planting density (HD) (56 plants/m2 ) and low planting density (LD) (28 plants/m2 ).
Figura 5.Tasa de aparición foliar (hojas/día) en plantas de repollo ornamental (Brassica oleraceaL. var.acephala) en función de la temperatura media diaria para los cv (A) Rose (Rs), (B) Red (Rd), (C) Pink (Pk) y (D) Bicolor (Bc) en los tratamientos con plantación a alta densidad (AD) 56 plantas/m2 y baja densidad (BD) 28 plantas/m2.
Regarding the root volume per plant, it was found significant differences (p≤0.05) due to the density and the cultivar, but not to the density x cultivar interaction (table 2, page 57).
Table 2. Summary of the results of the ANOVAs and interactions for B. oleracea L. var. acephala for each of the assessed variables.
Tabla 2: Resumen de los resultados de los ANOVAs y las interacciones para B. oleracea L. var. acephala en cada una de las variables evaluadas.

The lower planting density determined a greater root volume (mL/plant) in all the cultivars, with significant differences in Bicolor (figure 6, page 58).

Figure 6. Comparison of means (mean ± standard error of the mean) of the root volume (mL/plant) in ornamental cabbage plants (Brassica oleracea L. var. acephala) cv. Rose (Rs), Red (Rd), Pink (Pk) and Bicolor (Bc) at high planting density (HD) (56 plants/m2) and low planting density (LD) (28 plants/m2).
Figura 6. Comparación de medias (media ± error estándar de la media) del volumen radical (ml/planta) en plantas de repollo ornamental (Brassica oleracea L. var. acephala) de los cv. Rose (Rs), Red (Rd), Pink (Pk) y Bicolor (Bc) en los tratamientos con plantación a alta densidad (AD) 56 plantas/m2 y baja densidad (BD) (28 plantas/m2).
Also, the mean of the four cultivars showed that the condition of lower planting density generated a significant increase in this variable with a difference of 39.8%.
Both, AFW and RFW were affected by the density and the cultivar, but not by the interaction between both factors (table 2, page 57). The treatments at low density led, on average, to 34.5% higher AFW, with significant differences (p≤0.05) regarding the treatments at high density.
The same trend was observed for RFW, where the lower density in each cultivar led to an increase in RFW, with significant differences in Bicolor (figure 7A, page 58).

Figure 7. Comparison of means of aerial and root fresh (A) and dry (B) weight (mean ± standard error of the mean) in ornamental cabbage plants (Brassica oleracea L. var. acephala) cv Rose (Rs), Red (Rd), Pink (Pk) and Bicolor (Bc) at high planting density (HD) (56 plants/m2) and low planting density (LD) (28 plants/m2).
Figura 7. Comparación de medias de peso fresco (A) y seco (B) aéreo y radical (media ± error estándar de la media) en plantas de repollo ornamental (Brassica oleracea L. var. acephala) de los cv Rose (Rs), Red (Rd), Pink (Pk) y Bicolor (Bc) en los tratamientos con plantación a alta densidad (AD) 56 plantas/m2 y baja densidad (BD) (28 plantas/m2).
On average, the plants grown at low density increased their RFW by 36%, with significant differences (p≤0.05). Regarding ADW and RDW, there were significant differences due to the planting density and cultivar but not to the density x cultivar interaction (table 2).
The mean of the resulting ADW of the treatments at high density was 33.4% higher, with significant differences (p≤0.05). The average increase in RDW in the treatments at low density was 37.8%, also with significant differences (p≤0.05) regarding those at high density (figure 7B, page 59).

Figure 7. cont. Comparison of means of aerial and root fresh (A) and dry (B) weight (mean ± standard error of the mean) in ornamental cabbage plants (Brassica oleracea L. var. acephala) cv Rose (Rs), Red (Rd), Pink (Pk) and Bicolor (Bc) at high planting density (HD) (56 plants/m2) and low planting density (LD) (28 plants/m2).
Figura 7. cont. Comparación de medias de peso fresco (A) y seco (B) aéreo y radical (media ± error estándar de la media) en plantas de repollo ornamental (Brassica oleracea L. var. acephala) de los cv Rose (Rs), Red (Rd), Pink (Pk) y Bicolor (Bc) en los tratamientos con plantación a alta densidad (AD) 56 plantas/m2 y baja densidad (BD) (28 plantas/m2).
DISCUSSION
The growth and productivity of a crop are the result of the interaction between its genotype and the environment that surrounds it (21).
However, eventhough the genotype is relatively constant as compared with the environment, its expression is largely influenced by environmental changes and any variable affecting the environment will be reflected in the growth and productivity of the crop (18). Also, as stated by Casal (2013), the management of crops involves decisions on several factors such as the sowing density, defoliation, fertilization, architecture, and choice of cultivar, all of which affect the leaf area and the position of the foliage in relation to the angle of the incoming radiation and therefore the degree of shading.
In this experiment, the planting density notoriously influenced all the parameters evaluated, regardless of the cultivar. A greater planting density led to an increase in plant height probably due to the changes caused in the light environment, i.e. due to the decrease in the Red:Far-Red ratio caused by this treatment, generating shade avoidance as a response. This response, the best-known of those mediated by photosensors, leads to a change in the architecture of plants by the elongation of internodes (1).
According to Casal (2013), in dense canopies, upper leaves intercept radiation that would otherwise reach the lower leaves, and plants are mutually shaded. Shade implies the selective attenuation of the wave length of the incident radiation.
Green leaves absorb primarily in the range of UV radiation (280-320 nm for UVB and 320-400 nm for UVA) and in the range of the photosynthetically active radiation (400-700 nm), particularly in the blue and red regions of the spectrum; and transmit and reflect more strongly in the far red (700-800 nm) and infrared regions. According to Smith (1995), the Red:Far-Red ratio provides a highly reliable indication of the degree of shading because it is relatively little affected by other environmental conditions.
The changes in the Red:Far-Red ratio caused by the reflected light constitute an early signal of alert of the presence of neighboring plants, even before the mutual shading of the leaves begins.
In the present study, although the differences in plant height were not expressed clearly from the start of the experiment (may be because the plants had not reached a size large enough to generate competition for the resources of the environment), the stimulus caused by the proximity of neighboring plants could have been early detected by the plants cultivated at high density. Thus, the stem elongation could be attributed to the increased levels of gibberellins (in some cases through inhibition of their inactivation), caused by a low Red:Far-Red ratio (4).
Other researchers also point out that, in these cases, the responses of elongation seem to be based on the combined action of several plant hormones: gibberellins, auxins, brassinosteroids and ethylene (12, 14). Such elongation includes various physiological components and processes that control the extensibility of the cell wall through expansins and xyloglucan endotransglucosylase/hydrolases and by acidification of the cell wall (22, 23). On the other hand, the reduction of the irradiance and the blue/green ratio in dense canopies would provide signals that do not share all the features of the Red:Far-Red ratio but that are important for the responses of the plants to the shade.
The light spectrum under a dense canopy shows a relative increase in green light (500-580 nm) and a decrease in blue light (400-500 nm).
The green light reflected or transmitted through the surrounding plants can be another signal to detect neighboring plants, because the addition of green light induces the elongation of the hypocotyl in seedlings and the hypoplasia and elongation of the petiole in adult Arabidopsis plants (10, 27).
When the canopy closes and the competition between plants is established, additional signals, such as the decline in the photosynthetically active radiation and UV-B light, are generated. These signals can inform plants of the progressive intensity of competition. Similarly to the red light, the UV-B light is absorbed by chlorophyll and used in photosynthesis. Thus, it has been suggested that a relative decrease in UV-B in the light spectrum would also be a signal of detection of neighboring plants and shade.
In the present study, the increase in the height of the plants grown at high density was not accompanied by an increase in the aerial or root biomass. This greater plant height at high density could not be attributed to a differential stem:root partition because no significant differences in fresh or dry weight were found for such relationship between treatments with high and low densities. Numerous studies have shown that the responses of shade avoidance are often accompanied by a reduction in the investment of carbon in organs such as roots and leaves (2, 19).
According to Casal (2013), one of the responses of plants against the limited generation of photoassimilates under shading is the reduction of potential yield.
In the present study, the higher density of plants also reduced the rosette diameter in the same way in all the cultivars, although the rosette diameters accomplished were within the commercial range (7.5 cm-18 cm).
This results coincide with those reported by Carter (2003), who stated that a smaller spacing between plants promotes the formation of longer stems and reduces the rosette diameter of ornamental cabbages. This same trend has been found in other varieties of the genus Brassica. Studies in B. oleracea L. var. capitata have shown that the diameter of the "heads" decreases as the spacing between plants decreases (25).
In B. oleracea L. var italica, it has been found that a high planting density reduces the size and the weight of the floret (6) as well as the diameter of the stem and the yield per plant (7).
In the present study, the cumulative number of new leaves and the leaf area per plant decreased as density increased. This trend was recorded in the four cultivars. Similar results have been previously found by Criollo and García (2009) in radish plants (Raphanus sativus) cultivated in pot and in greenhouse, in which treatments with lower density allowed achieving a greater leaf area. As stated by these authors, the increase in population density caused a decrease in size, vigor and weight of the plants.
In this study, the rate of leaf appearance was lower in the treatments at high density than in those at low density, in coincidence with that reported by Jolliffe and Gaye (1995), who claimed that growth is faster and the expansion of leaves is greater when there are no competing plants nearby. However, a much more marked adjustment in the rate of leaf appearance as a function of the environmental temperature was found in the treatments at high density than in those at low density when the temperatures were between 17 and 27°C (temperatures that promote the active growth of B. oleracea acephala).
Temperature is one of the most important physical factors of the environment that influence the processes of growth and development of plants (17). This variable has a widely-accepted role as a main factor in the control of the phyllochron, since it affects different biochemical processes that regulate the meristematic activity, closely associated with the development of the leaf and stem (9, 16). Thus, the value of the adjustments obtained in the treatments at high density would allow hypothesizing that, once the stimulus that induces shade avoidance is captured (due to the modification in the Red:Far-Red ratio of dense canopies), plants could present a greater sensitivity to other environmental factors (in this case temperature) than the plants grown at low density. Also, it should be noted that, as time passed by, plants grown at high density lost the leaves in the lower strata of the crop. It is likely that the increase in density, due to the increase in the shading, generated their senescence and abscission.
Casal (2013) argues that, in the vertical avoidance of shading, the average position of foliage is high due to the increase in the height of the stem and/or a decrease in the foliage of the lower strata. In this way, on average, the foliage is less shaded because the upper layers have more light and the increase in leaf senescence reduces the foliage volume at the base of the canopy.
However, although it is important that plants retain a sufficient number of leaves along their cultivation cycle to ensure the growth through the contribution of photoassimilates, the abscission of leaves in the lower strata in the treatments at high density could represent a comparative advantage respect to the crop grown at low density, because one of the technical recommendations for crop management consists in removing the lower leaves ("defoliation"), after which plants reach an average height of 25 cm. Unlike that observed in other crops, in ornamental cabbages, the decrease in foliar area per plant (with the consequent reduction in the rosette diameter) and the natural abscission of the leaves of the lower strata of the crop would be effective to improve the visual appearance of the harvested plants. Besides their coloration, one of the main attributes of ornamental cabbages is the compactness of the apical rosette whose appearance resembles that of roses. This is fundamental because, unlike other foliage for cutting that act as accompaniment products or complements in bouquets or flower arrangements, ornamental cabbages for cutting are novel products, being protagonists as "cut flowers" within the bouquet.
The higher values of root volume per plant and leaf area per plant observed in all low density treatments could be attributed to the greater space available per plant, which allowed an increase in the exploration of the soil, thus giving plants a greater capacity of absorption of water and nutrients. This was reflected in the responses of AFW, RFW, ADW and RDW, which showed a clear tendency to increase with lower planting density (figure 7, page 58-59). These results agree with those of Pérez et al. (2005), who reported that high planting densities reduce the growth of vegetative variables such as leaf area, number of leaves and dry matter, being these effects more notorious at the end of the crop.
The differences found between cultivars, for each of the parameters evaluated, are probably due to genetic features, being an indicator of the potential of growth of each plant. In this sense, the cultivars Bicolor and Red showed better performance in most of the variables evaluated, whereas, under the same conditions, Rose reached the lowest values.
CONCLUSIONS
In the cultivars evaluated, the density of plants may be increased as an alternative that would not only allow a more efficient use of theby the crop, but also improve quality indices, achieving greater plant height and more compact apical rosettes, while reducing crop management activities, such as manual leaves removal, which is made naturally. However, further studies with other cultivars should be carried out to test the effect of different densities under the growth conditions of each cultivar, in order to find the optimal planting density in each case.
1. Ballaré, C. L.; Sánchez, R. A.; Scopel, A. L.; Casal, J. J.; Ghersa. C. M. 1987. Early detection of neighbour plants by phytochrome perception of spectral changes in reflected sunlight. Plant, Cell and Environment. 10: 551-557. [ Links ]
2. Carabelli, M.; Possenti, M.; Sessa, G.; Ciolfi, A.; Sassi, M.; 2007. Canopy shade causes a rapid and transient arrest in leaf development through auxin-induced cytokinin oxidase activity. Genes Dev. 21: 1863-68. [ Links ]
3. Carter, K. 2003. Ornamental Kale. Center for landscape and urban Horticulture. Cooperative Extension/Botany Plant Sciences Dept., University California Riverside. [ Links ]
4. Casal, J. J. 2013. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 64: 403-427. [ Links ]
5. Criollo, H.; García, J. 2009. Efecto de la densidad de siembra sobre el crecimiento de plantas de rábano (Raphanus sativus L.) bajo invernadero. Revista Colombiana de Ciencias Hortícolas. 3(2): 210-222. [ Links ]
6. Chung, B. 1982. Effects of plant density on the maturity and once over harvest yield of broccoli. J. Hort. Sci. 57: 365-372. [ Links ]
7. Damato, G. 2000. Late sowing dates and high plant density in six cultivars of broccoli for processing. Acta Hort. 533: 267-274. [ Links ]
8. Di Rienzo, J. A.; Casanoves, F.; Balzarini, M. G.; Gonzalez, L.; Tablada, M.; Robledo, C. W. 2008. InfoStat. Grupo InfoStat. FCA. Universidad Nacional de Córdoba. Argentina. [ Links ]
9. Ferraris, R.; Chapman, L. S. 1991. The dynamics of bud development in ratooning stubble of sugarcane varieties. Proc. Aust. Soc. Sugarcane Technol. 13: 164-171. [ Links ]
10. Folta, K. M. 2004. Green light stimulates early stem elongation, antagonizing light-mediated growth inhibition. Plant Physiology. 135: 1407-1416. [ Links ]
11. Gilman, E. F. 1999. Brassica oleracea flowering kale, ornamental cabbage. University of Florida. IFAS Extension. FPS71. [ Links ]
12. Gommers, C. M. M.; Visser, E. J. W.; St Onge, K. R.; Voesenek, L. A. C. J.; Pierik, R. 2013. Shade tolerance: when growing tall is not an option. Trends in Plant Science. 18: 65-71. [ Links ]
13. Greer, L.; Gibson, J. L.; Dole, J. M.; Whipker, B. E. 2003. Cut Kale-A blooming success. NCSU Floriculture Research and Extension. [ Links ]
14. Jaillais, Y.; Chory, J. 2010. Unraveling the paradoxes of plant hormone signaling integration. Nature Structural and Molecular Biology. 17: 642-645. [ Links ]
15. Jolliffe, P. A.; Gaye, M. M. 1995. Dynamic of growth and yield components of bell peppers (Capsicum annum L) to row covers and population density. Scientia Hort. 62: 153-164. [ Links ]
16. Kingston, G. 2000. Climate and the management of sugarcane. En: Hogarth, D. M. and P. G. Allsopp (eds.), Manual of Canegrowing. BSES. Brisbane. Australia. p. 7-25. [ Links ]
17. Machado-Neto, N. B.; Prioli, M. R.; Gatti, A. B.; Cardoso, V. J. M. 2006. Temperature effects on seed germination in races of common beans (Phaseolus vulgaris L.). Acta Sci. Agron. 28: 155-164. [ Links ]
18. Marín, D. 1989. Análisis de crecimiento en Canavalia ensiformis (L.) DC. bajo condiciones de campo. Rev. Fac. Agron. Univ. Del Zulia. Venezuela. 15: 1-16. [ Links ]
19. Morelli, G.; Ruberti, I. 2000. Shade avoidance responses. Driving auxin along lateral routes. Plant Physiology. 122: 621-626. [ Links ]
20. Perez, M.; Carew, L.; Battey, N. 2005. Efecto de la densidad de siembra sobre el crecimiento vegetativo y reproductivo de la fresa cv Elsanta. Bioagro. 17(1): 11-15. [ Links ]
21. Rincón, A.; Ligarreto, G.; Sanjuanelo, D. 2007. Crecimiento del maíz y los pastos (Brachiaria sp.) establecido en monocultivo y asociados en suelos ácidos del piedemonte llanero colombiano. Agron. Colomb. 25(2): 264-272. [ Links ]
22. Sasidharan, R.; Chinnappa, C. C.; Voesenek, L.; Pierik, R. 2008. The regulation of cell wall extensibility during shade avoidance: a study using two contrasting ecotypes of Stellaria longipes. Plant Physiology. 148: 1557-1569. [ Links ]
23. Sasidharan, R.; Chinnappa, C. C.; Staal, M.; Elzenga, J. T. M.; Yokoyama, R.; Nishitani, K.; Voesenek, L. A. C. J.; Pierik, R. 2010. Light quality-mediated petiole elongation in Arabidopsis during shade avoidance involves cell wall modification by xyloglucan endotransglucosylase/hydrolases. Plant Physiology. 154: 978-990. [ Links ]
24. Smith, H. 1995. Physiological and ecological function within the phytochrome family. Annu. Rev. Plant Physiol. Plant Mol. Biol. 46: 289-315. [ Links ]
25. Stepanovic, M. V.; Bjelicy, V. V.; Dragicevic, V. D. 2000. Effect of crop density on morphological characteristics and yield of cabbage. Acta Hort. 533: 205-207. [ Links ]
26. Torres Nava, D.; Sandoval Castro, E.; Peña-Cabriales, J. J.; Vera-Núñez, J. A. 2017. Aporte de nitrógeno proveniente de pollinaza al cultivo de brócoli (Brassica oleracea L.). Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 49(2): 105-116. [ Links ]
27. Zheng, X.; Wu, S.; Zhai, H. 2013. Arabidopsis phytochrome B promotes SPA1 nuclear accumulation to repress photomorphogenesis under far-red light. The Plant Cell. 25: 115-133. [ Links ]

