










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo
versión impresa ISSN 1853-8665versión On-line ISSN 1853-8665
Rev. Fac. Cienc. Agrar., Univ. Nac. Cuyo vol.50 no.2 Mendoza dic. 2018
ORIGINAL ARTICLE
Biomass function for Acacia caven (Mol.) Mol. distributed in the dry land areas of south central Chile
Función de biomasa para Acacia caven (Mol.) Mol. distribuida en áreas secas del centro sur de Chile
Alejandro Lucero Ignamarca 1, 2, Fernando Muñoz Sáez 2, Jorge Cancino Cancino 2, Alvaro Sotomayor Garreton 1, Francis Dube 2, Arnoldo Villarroel Muñoz 1, Katia Sáez Carrillo 3
1 Instituto Forestal, Sede Biobío. Camino a Coronel Km 7.5, San Pedro de La Paz, Concepción. Código Postal 4030000 Concepción. Casilla 109-C. Concepción. alucero@infor.cl
2 Universidad de Concepción. Facultad de Ciencias Forestales. Victoria # 631. Barrio Universitario. Concepción. C. P. 4070386. Concepción. Casilla 160-C Correo 3. Concepción.
3 Universidad de Concepción. Facultad de Ciencias Físicas y Matemáticas. Departamento de Estadísticas. Av. Esteban Iturra s/n. Concepción. C. P. 4030000. Concepción. Casilla 160-C Correo 3. Concepción.
Originales: Recepción: 24/10/2017 - Aceptación: 25/04/2018
ABSTRACT
Acacia caven is a key native species in the dry land region of south-central Chile. It is a species of high social and productive interest to landowners. Therefore, this study proposes a biomass function for A. caven, which involves incorporating predictor variables that are easily obtainable in the field and less complex than those used in already existing functions for the species. Due to the multi-purpose nature of the species and its potential for silvopastoral systems, these functions typify important planning tools to improve the management of this plant resource. To generate the biomass function, the methodology of destructive analysis of components was used on a total of 71 trees. These trees were selected, cut and weighed in the field to generate and test different models. Statistical analysis models were used and root collar diameter, diameter at breast height and total height served as predictor variables, resulting in a good adjustment of the models (R2 adjusted the 0.97 for stem-branches, 0.90 for stems and 0.96 for total biomass), with high correlations between estimated and real values. These functions may be used safely in plant formations of A. caven located in the same distribution area and within the range of the used variables. In the future, however, they require validation with new measurements in other sectors than the area considered to increase geographical representativeness.
Keywords: Espino; Steppe; Semi-arid regions of Chile; Silvopastoral systems
RESUMEN
Acacia caven, es una de las principales especies nativas presentes en el secano interior de la zona central de Chile. Es una especie que tiene un alto interés social y productivo para los propietarios de este recurso. En este sentido, el objetivo de este estudio es proponer una función de biomasa validada para A. caven, y que incorpore variables predictoras de fácil obtención en terreno y de menor complejidad que las consideradas en funciones ya conocidas para la especie. Dadas las características de especie multipropósito y su potencial para su uso en sistemas silvopastorales, estas funciones son importantes herramientas de planificación para mejorar la gestión y manejo de este recurso vegetacional. Para la generación de la función de biomasa, se utilizó una metodología de análisis destructivo por componentes de 71 árboles, los cuales fueron seleccionados, volteados y pesados en terreno, para posteriormente generar y probar diferentes modelos. A través de análisis estadísticos se seleccionaron modelos que utilizan como variables predictoras el diámetro a la altura del cuello, diámetro a la altura del pecho y altura total. Los resultados muestran un buen ajuste de los modelos (R2 ajust. de 0,97 para fuste-ramas, 0,90 para ramillas y 0,96 para biomasa total), lo que indica un alto nivel de correlación entre valores estimados y valores reales. Estas funciones se pueden utilizar con seguridad en formaciones vegetacionales de A. caven que se ubiquen en la misma zona distribucional del recurso y dentro de los rangos de las variables utilizadas, siendo conveniente en el futuro validarlas con nuevas mediciones de sectores distintos al área considerada para estos análisis, aumentando su representatividad geográfica.
Palabras claves: Espino; Estepa; Zona semiárida de Chile; Sistemas silvopastorales
INTRODUCTION
The steppe or savanna formations, where Acacia caven (Mol.) Mol. ("espino") represents the dominant and co-dominant species, cover approximately 957.000 hectares in Chile (14) especially between the Coquimbo Region in the north and the Biobío Region in the south. Espino can be found in both pure formations and in association with tree species such as quillay (Quillaja saponaria), litre (Lithraea caustica), maitén (Maitenus boaria), huingán (Schinus polygamus) and others (32).
A. caven provides multiple benefits and so counts as multi-purpose species. It serves as firewood, for charcoal production, or atmospheric nitrogen sequestration (being a leguminous species). Its ecological use in erosion control, increase of organic matter and retention of soil humidity, nutrient recycling and the creation of a favorable micro-climate for the growth of vegetation under its influence, raise its importance further (13, 20, 23).
From a productive and ecological viewpoint, A. caven is among the main species present in the dry land area in south-central Chile and in countries such as Argentina, Bolivia, Uruguay, Paraguay and Brazil (13).
However, mainly as a result of human pressure, it is being degraded, inducing loss of economic value of soil and vegetation (7, 13). Despite such degeneration of A. caven formations, their restoration has been envisaged (31) by applying silvopastoral systems as a sustainable production alternative (16, 21, 22, 24).
In line with their appearance, Espinal formations are complex with a woody stratum of varying cover and height, dominated almost exclusively by A. caven (25). Currently existing formations correspond to an environmental unit of mainly secondary origin (9, 13, 31); composition and structure developed as consequence of human intervention. Land preparation for agricultural use altered this resource decisively predominantly for the production of wheat and cattle, albeit later being abandoned due to market fluctuations and low productivity. Espinal cover of the resulting formations is less dense, formed by smaller individuals mainly from stump sprouts; on rare occasions, where vegetation remained free of incisive alteration, denser formations can be found (7).
In order to allow for a sustainable use of A. caven and considering that it mainly provides firewood and charcoal, it is necessary to count on predictor functions that make it possible to estimate the biomass this resource potentially generates. Estimations, however, remain challenging due to the high variability of the species.
Functions make it possible to estimate biomass through predictor variables which can easily be measured in the field, such as the diameter at breast height (DBH) and total height (H), or crown diameter and length, basal diameter and length of branches, producing generic (for various species) or specific (for one only) functions (3, 11, 27).
Various studies have produced biomass functions for A. caven in the north-central part of its natural distribution in Chile (1, 10, 26, 30). These functions, nevertheless, lose precision when applied to other regions with different conditions than those of the study area, and different ranges and parameters than the original resource (1, 6, 17, 30). Predictor variables used in these functions, and for other tree species, are mainly DBH, root collar diameter (RCD), number of commercial branches, crown diameter, trunk height, commercial height, medium crown diameter, height of maximum foliage, number of stems (1, 2 , 4, 10, 26, 28, 30).
This study aims to develop a biomass function for A. caven present in the southern part of its geographical distribution area. The function is validated for the dry inland area of the Maule Region (Chile), considering predictor variables that are easily obtainable in the field, with less complexity than those considered in already existing functions, making them a key tool for improving the sustainable management of this native plant species.
MATERIAL AND METHOD
Location and description of the Study Area
The study area covers a surface of 2.4 hectares of adult A. caven, with the coordinates latitude 35°53'34" and longitude 72°08'44", within farm "San Adolfo", km 48 of "Los Conquistadores" Road, Cauquenes community, Maule Region.
Methodology for inventory and biomass measurements
The predictor function was generated using the method of direct biomass determination or destructive analysis (3). This required a previous enumeration in the study area, during which each tree got classified as mono- or pluri-fustal (2 or more stems) according to growth habit, RCD and DBH variables and H of each shank or stem. These variables were measured on a total of 3,367 single- and multi-stem A. caven trees distributed across the study area.
In order to select the sample trees for the destructive analysis, the observed frequency (OF) of the resource was determined first by using the root collar diameter variable, as it represents the most common variable in native forests with characteristics like that of A. caven and is also recommended by Gajardo & Verdugo (10).
Furthermore, OF was determined differentially according to root collar diameter due to the existence of single- and multi-stem individuals, and to increase representativeness of the diameter classes to be sampled (figure 1 and figure 2).
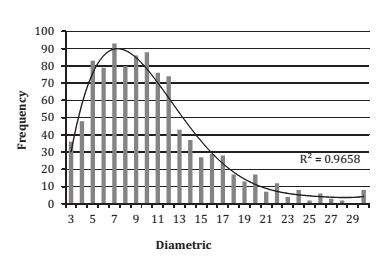
Figure 1. Observed frequency according to root collar diameter (RCD) A. caven single-stem.
Figura 1. Frecuencia observada según diámetro a la altura del cuello (DAC) de árboles monofustales de A. caven
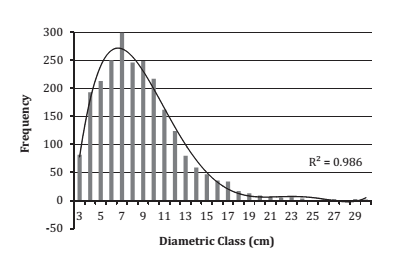
Figure 2. Observed frequency according to root collar diameter (RCD) A. caven multi-stem.
Figura 2. Frecuencia observada según diámetro a la altura del cuello (DAC) de árboles plurifustales de A. caven.
After determining the OF according to growth habits, a minimum of 20 trees was selected (35) which were distributed proportionally to diameters, with the restriction that at least one individual for each diameter class (if existing) was sampled.
In total, 71 trees (single- and multistem) were cut for the destructive analysis. The material extracted from these trees was classified into stem-branches -including the woody material with minimum diameter of 3 cm-and twigswoody material with diameter below 3 cm (including leaves and fruits) (10). Both categories were weighed in the field using a metallic support device designed specifically for this purpose and scales with a maximum capacity of 200 kg ± 0.1 kg, thus obtaining the green weight of each component for each tree.
To determine the relation between dry weight and green weight and thus estimate the dry weight of the components for the generation of the biomass function, three random samples of each component were taken from five randomly selected trees, both single- and multi-stem, of the 71 cut trees.
These samples were first weighed in the field (green weight) and then put into bags and labeled for dispatch to the laboratory of the Forestry Institute in Concepción.
The dry weight was obtained after drying the material in an Yih Der oven, DK-600DT model (Taiwan), with forced ventilation at 103°C ± 2°C, until obtaining steady weight between measurements every 2 or 4 hours (15). With the dry, green and total weight data the dry weight ratio (r sv ) per component according to growth habit (single- or multi-stem) and in total was determined, using equation 1 (3, 18). The extant literature for A. caven reports dry weight ratio values (r sv ) for stem-branches of 0.62 and for twigs 0.68 (30), so it was expected similar values for this study

where:
r sv = dry weight ratio
w s_ = dry weight of sample unit i
w v_i= green weight of sample unit i
n = size of selected sample (n≥1)
The information obtained in the field and described above was then systematized and processed for generating the biomass function (BF).
On specific occasions when the stem below 1.3 m split into two or more sub-stems thus generating more than one DBH, the medium DBH (DBH m ) and equivalent DBH (DBH e ) (3) were calculated according to equations 2 and 3 respectively, and these variables were included as part of the predictor variables to determine the biomass function. Importantly, in those trees with only one DBH (one stem) both, DBH m and DBH e equal DBH.

where:
DBH m = medium diameter at breast height in cm.
DBH i = diameter at breast height of sub-stem i in cm
n = number of sub-stems

where:
DBH e = equivalent diameter at breast height in cm
DBH i= diameter at breast height of sub-stem i in cm
n = number of sub-stems with DBH
Adjustment and selection of biomass function (BF)
One of the objectives when selecting the models was to acquire simpler functions than the existing ones for the species (northern part of its natural distribution area). Therefore, analyzed functions contemplated only variables that are easily measurable in the field. In the process of selecting the predictor functions, DBH e and DBH m were considered and analyzed independently, as was assumed that one of these variables related better to the DBH variable when contemplating more than one value, because no tested model has considered these variables in conjunction. Indeed, existing models contemplate these variables only independently in relation to other variables such as H, RCD, or crown starting height and radius (upper and lower crown diameter).
When analyzing the relation between DBH and H to evaluate the coherence of the in-field data, generally the expected correlation of higher H with higher DBH was maintained. In accordance with the information from the field, the expected relation of higher diameter class related to higher biomass was confirmed. This initial analysis gave information on the reliability of the in-field data, validating it for subsequent use for determining the biomass functions.
For the generation of the dry biomass function per components and in total, simple and multiple regression models were adapted, using 12 models as a guideline (6, 11, 29, 35) for the variable type used and for each component (stem-branches, twigs & total). Moreover, 12 additional trial models were obtained through the Step-Wise method. In total 147 models were tested, considering the modification of variables, namely the use of variables as measured in the field, their squared and cubed value, transformed into natural logarithm and the combinations between them.
Selection of the final models rested on their compliance with the distributional and qualification assumptions of each model in relation to the adjusted statistic, the Mean Squared Error (MSE), the Prediction Error Sum of Squares (PRESS) and the Furnival Index (FI). The latter enables comparing models using transformed variables with models of non-transformed variables (19, 33, 35). Also, only models using variables with p- value<0.05 were considered. For all analyses SPSS 21.0 software (New York, USA) was used.
RESULTS
Table 1 (page 193), shows the maximum, minimum and standard deviation range of the state variables of the 71 trees cut (36 single-stem and 35 multi-stem) for the destructive analysis to be carried out according to the selection methodology described in section 2.
Table 1. Range of state variables for the trees used in the destructive analysis and calculation of Rsv . Minimum, maximum and standard deviation values are given according to growth habit.
Tabla 1.Rango de variables de estado de los árboles utilizados para análisis destructivo y cálculo de R sv . Se muestran los valores mínimos, máximos y desviación estándar por hábito de crecimiento.
In addition, the rank of the state variables of the subsample of randomly selected trees is included to calculate r sv . Table 2 (page 194), shows r sv
values for each component and in total, subsequently used to determine the dry weight of each of the 71 trees evaluated in the field to specify dry biomass models.
Table 2. Dry weight / green weight ratio per growth habit, per component and total (value ± standard deviation).
Tabla 2. Razón peso seco / peso verde por hábito de crecimiento, por componentes y total (valor ± desviación estándar)
For the selection of the best models for the components among already proven models, a total of 18 models complying with the distributional assumptions were selected. These models were then qualified according to the statistics calculated for each of them in order to be able to compare them, with value 1 given to the best model according to the mentioned criteria. (For example, value 1 was assigned to the model with the best value for adjusted R2 etc.)
Eventually, the best model selected turned out the one with the lowest total accumulated qualification (table 3, page 195).
Table 3. Qualification and pre-selection of models listed according to lowest total qualification conforming to four statistics
Tabla 3. Calificación de modelos preseleccionados ordenados de acuerdo con la menor calificación total según cuatro estadígrafos.
Table 4 (page 194), shows the model selected for each component and total.
Table 4. Selection of models according to statistics.
Tabla 4. Selección de modelos de acuerdo con estadígrafos
Given this particular situation, the models selected were linearized applying the natural logarithm function, correction factor Fc was applied according to equation 4 below (2, 8, 11, 17, 36):
Fc = e MSE (4)
where:
Fc = correction factor
MSE = mean squared error
e= base natural logarithm
Subsequently, these models were submitted to a comparative analysis charting the dry weight values of observed (real) biomass in relation to the dry weight of estimated biomass (figure 3), to determine whether the selected models under- or overestimate tree biomass (8, 35).

Figure 3. Comparative analysis of model precision for components of A. caven. (a) stem-branches, (b) twigs, (c) total weight.
Figura 3. Análisis comparativo de la precisión de los modelos seleccionados por componente de A. caven. (a) fusteramas, (b) ramillas y (c) peso total.
If the models measure adequately, the charted dots should run along or near line y = x.
DISCUSSION
The enumeration carried out at the beginning helped proving the high variability of this species. Although in general the expected growth tendencies were confirmed, e.g., a higher RCD equals more biomass, this did not always hold, because mechanical damage (broken branches, rot, signs of decay, etc.) of some individuals affected their normal production and consequent biomass.
The use of the observed frequency criterion according to the RCD variable, following Gajardo & Verdugo (1979), for the selection of trees to be cut guaranteed high reliability of the model selection during the sampling process.
Regarding the easy measurement of the variables used in the models, it should be noted that e.g., Gonzalez-Benecke et al. (2014) indicate that in order to obtain estimations that are more precise and representative models, other variables, like age, wood density and/or productivity, should be included.
However, these variables complicate the actual applicability of the models in the field, and make in-field measurements difficult, considering in particular the great diversity of the species. Thus it was preferred sticking to variables which can be applied easily.
Combining the information obtained in the field with database analysis facilitated the generation of more than 147 preliminary models, based on the most commonly reported models found in the literature (6, 11, 29, 35). It also reinforced this confidence in the methodology proposed by Segura & Andrade (2008) utilizing the models selected for each component, and determine which one best adapts to the production of aerial biomass of A. caven in this particular region.
Out of these models, 18 with values of adjusted R2 between 87% and 97% (p<0.05) were pre-selected, resulting in good to very good data adjustment (table 3, page 195). Also, the selected functions consider the use of easy-to-use regression variables such as RCD, DBH e and H which can be obtained effortlessly in the field (table 4, page 194) (8). They also strengthen the models' reliability and usability, making it superior to other models, which consider variables such as RCD, medium DBH of sprouts, height of maximum foliage, upper and lower crown diameter and total height (13).
The proposed models (table 4, page 194) do not differentiate between singleor multi-stem individuals, although initially the sampling methodology had considered this differentiation.
The models obtained consider each sapling as an individual, no matter if several are part of one stump (multi-stem). This kind of model makes it necessary to measure the total height of each existing sapling, which may imply more measuring work, but also leads to higher representativeness than the in-field estimation of medium height of multi-stem individuals.
Moreover, with the existing technology of dasometric instruments, and as conducted in this study, the variable can be measured quickly, allowing the use of H as estimator for the two growth habit categories.
Also in table 4 (page 194), the lower values of adjusted R2 correspond to the models related to the twigs component with values between 0.87 and 0.90, which was expected considering the difficulty of finding a model for the crown biomass with state variables such as RCD and DBH e , and the high variability of the resource in the field.
The higher adjusted R2 values are found in the stem-branches and total components, which indicates that biomass estimations of these components relate better to the state variables of the trees, confirming Segura & Andrade's (2008) finding in relation to biomass model determination. They point out that a high correlation of the models for stem and branches with variables such as DBH is expected, but not so with leaves and twigs.
Regarding precision of the models it turns out that the models for stembranches and total components are more precise (figure 3a and figure 3c, page 196), although some over- and under estimations occur as biomass production increases. This has also been discussed by Segura & Andrade (2008).
In the case of the twigs component (figure 3b, page 196), the model is acceptable for lower values, but tends to underestimate biomass for higher values. It was believed that this tendency may prove significant in relation to the obtained frequency for individuals with RCD longer than 20 cm (on average one individual per diameter class), although this almost never occurs.
This makes it more difficult to elaborate a model for these diameter classes compared to individuals with a lower RCD, which were well represented in the in-field sampling process, again confirmed via the three models' perfectly acceptable precision level, both for lower and medium biomass production. Additionally, the studentized residue dispersion graphs (figure 4) show that the dot clouds have a random behaviour around 0, which indicates that the data fit properly to the models.
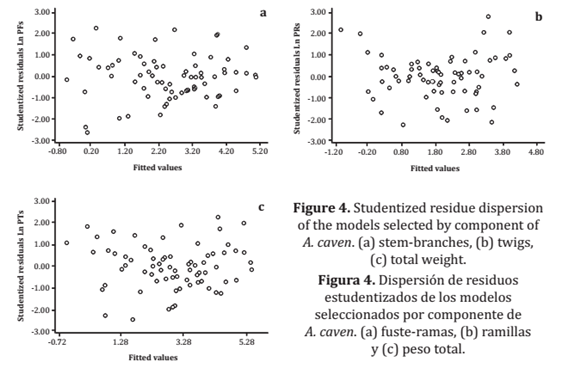
Figure 4. Studentized residue dispersion of the models selected by component of A. caven. (a) stem-branches, (b) twigs, (c) total weight.
Figura 4. Dispersión de residuos estudentizados de los modelos seleccionados por componente de A. caven. (a) fuste-ramas, (b) ramillas y (c) peso total.
It should be noted that the selected models by design do not comply with additivity property, although for Dieguez et al. (2003) this counts as one of the most important properties models should comply with. Yet, since the selected models are log-linearized, Cancino's (2006) argument holds, stating that the summed-up biomass of the estimated components for this type of model does not equal total estimated biomass, so they cannot be harmonized, and the additivity concept is not admissible in these particular situations.
In relation to the better adjusted variables (RCD, DBH e & H), it was rightly expected that RCD adjusts well, as this variable has been representative for characterizing these plant formations (10). DBH e variable adjusted better than DBH m in all selected models, pointing to higher representativeness than real DBH, as it produces better ponderation of the different DBH measured in the field than their simple measurement (Eq. 1 and 2, page 191). In the case of H it was also to be expected to rely on this variable in the stem-branches and total models, as this variable normally associates directly to biomass production-higher H equaling more biomass production.
A literature review on the determination of biomass in Peumus boldus (6, 11, 12) reveals that the state variables that best explain biomass production of the different species analyzed here can be multiple, but they also affirm that easily measurable variables such as RCD, DBH and H are more characteristic and representative.
In the selected models the coefficient of determination adjusted R2 is high, showing values of 0.97 for stem-branches, 0.90 for twigs and leaves, and 0.96 for total biomass, indicating a high level of correlation between estimated and real values. These values are similar to the R2 values presented by Aguirre & Infante (1988), Prado et al. (1988) and INFOR (2012) for the stem-branches (between 0.93 and 0.97), twigs (0.92) and total (between 0.96 and 0.98) components.
CONCLUSIONS
The initial characterization of the resource (enumeration) produced good information regarding distribution according to diameter classes, which substantially improved the sampling of the trees to be cut and their representativeness in what followed. Although methodology of biomass analysis with the destructive sampling process involves high costs, it still serves well to obtain this type of functions, which becomes clear considering the good adjustment level obtained in the selected models.
These functions, with the used methodology and adjustments, can reliably be used in formations of A. caven in the distribution area of the dry inland region between the Cauquenes (VII Region) and Chillán (VIII Region) communities within the range of the state variables used in this study (table 1, page 193). However, it is recommended to validate and adjust them with new measurements of trees from other regions, too, to get a higher level of geographical representativeness and reliability.
These new biomass functions with a high correlation level are a new tool for the forestry and agricultural sectors helping to better plan and manage this important resource in the dry land regions of southcentral Chile.
The potential of A. caven formations for the application of silvopastoral systems is well known, partly because it is a multi-purpose species that facilitates interactions between trees, grassland and livestock. In this context the new functions generated and proposed in this study allow for a more adequate and reliable assessment of the tree component before commencing silvopastoral management, and may also support new research to optimize these systems and the interrelations between their components.
1. Aguirre, S.; Infante, P. 1988. Funciones de biomasa para boldo (Peumus boldus) y espino (Acacia caven) de la Zona Central de Chile. Revista Ciencia e Investigación Forestal. Instituto Forestal. Chile. Vol. 2 N°3. 6 p. [ Links ]
2. Blujdea, V.; Pilli, R.; Dutca, I.; Ciuvat, L.; Abrudan, I. 2012. Allometric biomass equations for young broadleaved trees in plantations in Romania. Forest Ecology and Management. 264: 172-184. [ Links ]
3. Cancino, J. 2006. Dendrometría Básica. Departamento Manejo de Bosques y Medio Ambiente. Facultad de Ciencias Forestales. Universidad de Concepción. Chile. Editorial Talleres de Dirección de Docencia. 171 p [ Links ]
4. Del Fierro, P. 2001. Efecto de tratamientos de corte sobre el rebrote de Acacia caven (Mol.) Mol en Auco, IV Región. Memoria para optar a título profesional de Ingeniero Forestal. Departamento de Silvicultura. Escuela de Ciencias Forestales. Facultad de Ciencias Forestales. Universidad de Chile. 83 p. [ Links ]
5. Diéguez, U.; Barrio, M.; Castedo, F.; Ruíz, A.; Álvarez, M.; Álvarez, J.; Rojo, A. 2003. Dendrometría. Coedición Fundación Conde del Valle de Salazar. Ediciones Mundi-Prensa. 340 p. [ Links ]
6. Djomo, A.; Ibrahima, A.; Saborowski, J.; Gravenhorst, G. 2010. Allometric equations for biomass estimations in Cameroon and pan moist tropical equations including biomass data from Africa. Forest Ecology and Management. 260: 1873-1885. [ Links ]
7. Donoso, C. 1982. Reseña ecológica de los bosques mediterráneos de Chile. Bosque. 4(2): 117-146. [ Links ]
8. Fonseca, W.; Alice, F.; Rey, J. M. 2009. Modelos para estimar la biomasa de especies nativas en plantaciones y bosques secundarios en la zona Caribe de Costa Rica. Bosque. 30(1): 36-47. [ Links ]
9. Gajardo, R. 1983. Sistema básico de clasificación de la vegetación nativa chilena. Facultad de Ciencias Agrarias, Veterinarias y Forestales. Universidad de Chile. Santiago. Chile. 93 p. [ Links ]
10. Gajardo, M.; Verdugo, R. 1979. Rendimiento en hojas de boldo, (Peumus boldus Mol), corteza de quillay (Quillaja saponaria Mol), y carbón de espino, (Acacia caven Mol), en la V Región. Memoria para optar al título de Ingeniero Forestal. Facultad de Ciencias Forestales. Universidad de Chile. Santiago. Chile. 99 p. [ Links ]
11. Gonzalez-Benecke, C.; Gezan, S.; Albaugh, J.; Lee Allen, H.; Burkhart, H.; Fox, T.; Jokela, E.; Maier, C.; Martin, T.; Rubilar, R.; Samuelson, L. 2014. Local and general above-stump biomass function for loblolly pine and slash pine trees. Forest Ecology and Management. 334: 254-276. [ Links ]
12. INFOR. 2011. Boldo. Rescate de un patrimonio forestal chileno. Manejo sustentable y valorización de sus productos. Benedetti, S.; Barros, S. (eds.). Santiago. Chile. 235 p. [ Links ]
13. INFOR. 2012. Monografía del Espino. Acacia caven (Mol.) Mol. Programa de investigación de productos forestales no madereros. Benedetti S (ed.). Santiago. Chile. 71 p. [ Links ]
14. INFOR. 2015. Estimated surface according to the GIS Department of INFOR Biobío, based on updated information from the cadastre of native plant resources of Chile from 2015 (Catastro de los recursos vegetacionales nativos de Chile) under administration of the National Forestry Corporation (Corporación Nacional Forestal/CONAF). Not published. [ Links ]
15. INN. 1984. Norma Chilena Oficial: NCh0176/1.Of84. Madera Parte 1: Determinación de la humedad. Santiago. Chile. 10 p. [ Links ]
16. Mahecha, L. 2002. El silvopastoreo: una alternativa de producción que disminuye el impacto ambiental de la ganadería bovina. Revista Colombiana de Ciencias Pecuarias. Vol. 15 (2): 6 p. [ Links ]
17. Manuri, S.; Brack, C.; Puji Nugroho, N.; Hergoualc'h, K.; Novita, N.; Dotzauuer, H.; Verchot, L.; Septiadi, Ch.; Widyasari, E. 2014. Tree biomass equations for tropical peat swamp forest ecosystems in Indonesia. Forest Ecology and Management. 334: 241-253. [ Links ]
18. Muñoz, F.; Cancino, J.; Rodríguez, R.; Olave, R. 2018. Evaluación de crecimiento, rendimiento y calorimetría de biomasa de Miscanthus x giganteus (Poaceae) establecido en el centrosur de Chile. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 50(1): 47-60. [ Links ]
19. Murillo-Brito, Y.; Domínguez-Domínguez, M.; Martínez-Zurimendi, P.; Lagunes-Espinoza, L. del C.; Aldrete, A. 2017. Índice de sitio en plantaciones de Cedrela odorata en el trópico húmedo de México. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 49(1): 15-31. [ Links ]
20. Olivares, A. 2006. Relaciones entre el estrato arbóreo, el estrato herbáceo y la conducta animal en el matorral de Acacia caven (espinal). Science et changements planétaires/ Sécheresse. 17(1-2): 333-334. [ Links ]
21. Ovalle, C. 1986. Etude du système écologique sylvo-pastoral à Acacia caven (Mol.) Hook. et Arn: aplications à la gestion des ressources renouvelables dans l'aire climatique méditerranéenne humide et sub-humide du Chili. Centre Emberger (Francia). Groupe d'Ecologie des Ressources Renouvelables. Montpellier. Francia. 224 p. [ Links ]
22. Ovalle, C.; Avendaño, J. 1984a. Utilización silvopastoral del espinal. I. Influencia del espino (Acacia caven Mol.) sobre la productividad de la pradera natural. Agricultura Técnica (Chile). 44(4): 339-345. [ Links ]
23. Ovalle, C.; Avendaño, J. 1984b. Utilización silvopastoral del espinal. II. Influencia del espino (Acacia caven Mol.) sobre algunos elementos del medio. Agricultura Técnica (Chile). 44(4): 353-362. [ Links ]
24. Ovalle, C.; Aronson, J.; Del Pozo, A.; Avendaño, J. 1990. The espinal: agroforestry systems of the mediterranean-type climate region of Chile. Agroforestry Systems. 10(3): 213-239. [ Links ]
25. Ovalle, C.; Squella, F. 1988. Terrenos de pastoreo con praderas anuales en el área de influencia climática mediterránea. In I. Ruiz (ed.). Praderas para Chile. Instituto de Investigaciones Agropecuarias (INIA). Ministerio de Agricultura. Santiago. Chile. Capítulo 20: 369-409. [ Links ]
26. Oyarzún, M.; Palavicino, V. 1984. Evaluación de especies leñosas para ser usadas con fines energéticos en la Provincia de Choapa, IV Región. Tesis Ingeniero Forestal. Universidad de Chile. Facultad de Ciencias Agrarias y Forestales. Santiago. Chile. 159 p. [ Links ]
27. Pajtik, J.; Konôpka, B.; Lukac, M. 2008. Biomass function and expansion factors in young Norway spruce (Picea abies [L.] Karst) trees. Forest Ecology and Management. 256: 1096-1103. [ Links ]
28. Parra, G. 2001. Funciones de biomasa total y por componentes del "Espino" (Acacia caven Mol.) en Pencahue, VII Región. Tesis Ingeniero Forestal. Escuela de Ingeniería Forestal. Facultad de Ciencias Forestales. Universidad de Talca. 70 p. [ Links ]
29. Parresol, B. 2001. Additivity of nonlinear biomass equation. Can. J. For. Res. 31: 865-878. [ Links ]
30. Prado, J.; Infante, P.; Arriagada, M.; Aguirre, S. 1988. Funciones de Biomasa para Siete especies arbustivas de la IV Región. Proyecto "Investigación y desarrollo de áreas silvestres zonas áridas y semi áridas de Chile". CONAF/PNUD/FAO-CHI/83/017. Documento de Trabajo N°14. Santiago. Chile. 26 p. [ Links ]
31. Quintanilla, V. 1977. Diccionario de Biogeografía para América Latina. Ediciones Universitarias de Valparaíso. Universidad Católica de Valparaíso. Valparaíso. Chile. 249 p. [ Links ]
32. Rodríguez, R.; Matthei, O.; Quezada, M. 1983. Flora arbórea de Chile. Editorial de la Universidad de Concepción. Universidad de Concepción. 408 p. [ Links ]
33. Rodríguez-Toro, A.; Rubilar-Pons, R.; Muñoz-Sáez, F.; Cártes-Rodríguez, E.; Acuña-Carmona, E.; Cancino-Cancino, J. 2016. Modelo de ahusamiento para Eucalyptus nitens, en suelos de cenizas volcánicas de la región de La Araucanía (Chile). Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 48(1): 101-114. [ Links ]
34. Root-Bernstein, M.; Jaksic, F. 2013. The Chilean espinal: restoration for a sustainable silvopastoral system. Restoration Ecol. 21(4): 409-414. [ Links ]
35. Segura, M.; Andrade, H. 2008. ¿Cómo construir modelos alométricos de volumen, biomasa o carbono de especies leñosas perennes? Agroforestería en las Américas. 46: 89-96. [ Links ]
36. Sprugel, D. 1983. Correcting for bias in log-transformed allometric equations. Ecology. 64(1): 209-210. [ Links ]
ACKNOWLEDGEMENTS
The authors thank the Forestry Institute of Chile (INFOR) for the direct support in the realization and funding of this study. Mr. Marco Antonio Bejar Vega, owner of "San Adolfo" compound where this research took place. To the National Scientific and Technological Research Commission (CONICYT). The professional and technical staff of the Biobío office of the Forestry Institute of Chile, namely Mauricio Navarrete, Jorge Acevedo, Hernán Soto, Patricio Chung, Juan Carlos Muñoz, Alberto Avila, Juan Carlos Pinilla, Hans Grosse and Luis Vásquez, who contributed with their experience and knowledge to the implementation of this study in the different stages in the field and laboratory.

