










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo
versión impresa ISSN 1853-8665versión On-line ISSN 1853-8665
Rev. Fac. Cienc. Agrar., Univ. Nac. Cuyo vol.51 no.1 Mendoza jun. 2019
ORIGINAL ARTICLE
Effects of salt stress on germination, seedling growth, osmotic adjustment, and chlorophyll fluorescence in Prosopis alba G.
Efectos del estrés salino sobre la germinación, crecimiento de plántulas, ajuste osmótico y fluorescencia de la clorofila en Prosopis alba G.
Diego Ariel Meloni 1, Marta Rosalía Gulotta 1, Diolina Moura Silva 2, María Paz Arraiza 3
1 Universidad Nacional de Santiago del Estero. Av. Belgrano (S) 1912. 4200. Santiago del Estero. Argentina. dmeloniunse@gmail.com
2 Universidade Federal do Espírito Santo. Departamento de Ciencias Biológicas. Núcleo de Pesquisa em Fotossíntese. Vitória. ES. Brasil.
3 Universidad Politécnica de Madrid. Escuela Técnica Superior de Ingenieros de Montes, Forestal y Medio Natural. España.
Originales: Recepción: 14/03/2018 - Aceptación: 17/10/2018
ABSTRACT
Prosopis alba G. is a species of high forest importance in the phytogeographical region of Western Chaco. Although P. alba has been considered as salinity tolerant, its salinity thresholds for germination and seedling growth are unknown, as well as the physiological mechanisms involved in them. The aim of this study was to elucidate the mechanisms of tolerance of P. alba to salt stress. Seed germination, seedling growth, osmotic adjustment, and chlorophyll fluorescence were- analyzed. Germination was more tolerant to salinity than seedling growth, with thresholds of 600 mM and 500 mM, respectively. The species showed a high capability of osmotic adjustment, with values near to those observed in halophytes. The photochemical phase of photosynthesis was highly tolerant to saline stress, showing photoinhibition from 400 mM NaCl, as indicated by the fluorescence variables of chlorophyll. This behavior was associated to an increase in anthocyanin concentrations in the leaves.
Keywords: Germination; Seedling growth; Osmotic adjustment; Chlorophyll fluorescence
RESUMEN
Prosopis alba G es una especie de importancia forestal en la Región fitogeográfica del Chaco Occidental. Aunque P. alba ha sido mencionada como tolerante a la salinidad, no se conocen los umbrales de salinidad para la germinación y el crecimiento de plántulas, ni los procesos fisiológicos involucrados. El objetivo de este trabajo fue dilucidar los mecanismos de tolerancia de P. alba al estrés salino. Se estudió la germinación de semillas, el crecimiento de plántulas, ajuste osmótico y fluorescencia de la clorofila. La germinación fue más tolerante a la salinidad que el crecimiento de plántulas, con umbrales de 600 y 500 mM, respectivamente. La especie mostró una alta capacidad de ajuste osmótico, con valores cercanos a los observados en halófitas. La etapa fotoquímica de la fotosíntesis fue altamente tolerante al estrés salino, mostrando fotoinhibición a partir de 400 mM de NaCl, tal como lo indicaron las variables de fluorescencia de la clorofila. Este comportamiento estuvo asociado a un incremento en la concentración de antocianinas en hojas.
Palabras clave: Germinación; Crecimiento de plántulas; Ajuste osmótico; Fluorescencia de la clorofila
INTRODUCTION
Prosopis alba G. is a tree species of forest importance in the phytogeographical region of Western Chaco. It is used for furniture and carpentry, and its fruits are consumed by people and cattle. It inhabits the humid savanna and the most humid low lands with sandy soils, forming belts around saline depressions (7).
Germination and seedling establishment constitute the most critical periods among the physiological processes in the life cycle of the plants that grow in arid regions; therefore, the presence of adaptations at these ontogenetic stages can determine their natural distribution (10, 32). Salinity can inhibit germination and seedling growth by preventing water absorption due to either the low water potential of the soil solution or the toxic effect of ions (31).
Among the terrestrial nitrogen-fixing plants, the genus Prosopis presents high tolerance to salinity. P. juliflora (5), P. alpataco, and P. Argentina (33) have shown the capability to grow in salinities at about 500 mM NaCl. Velarde et al. (2003) compared the survival of 27 Prosopis families (seed from individual mother trees) as a function of salinity from 10 to 45 dSm-1. The P. pallida families had significantly greater mean survival than P. alba at seawater salinities of 45 dSm-1 (61.1% vs. 41.7%).
Osmotic adjustment (OA) has been considered an important physiological adaptation associated with tolerance to salt stress and has received much attention in recent years (11). It is the accumulation of osmotically active solutes (proline, glycine betaine, sugar alcohols, etc.) that reduce the water potential of tissues, maintaining a gradient with the soil solution to allow water absorption. This strategy enables the maintenance of cell turgor and the physiological processes that depend on it, such as growth (19). Stored solutes and its contribution to OA vary widely among species. For instance, in Prosopis strombulifera the degree of tolerance to salt stress correlates with the concentration of proline, polyols, and ions, which also contribute to OA (17).
Saline stress affects photosynthesis either directly or indirectly. In order to understand the physiological status in higher plants and determine photosynthetic damage affected by environmental stresses, chlorophyll fluorescence assay is a rapid and sensitive measure of photosynthetic competence (1).
The aim of this study was to contribute with information on the mechanisms of tolerance of P. alba to salt stress. The working hypotheses were that P. alba tolerates saline concentrations close to seawater (500 mM NaCl), performs osmotic adjustment, and exhibits a photochemical stage of stable photosynthesis at high saline concentrations.
MATERIALS AND METHODS
Plant Material
The fruits were harvested randomly within a P. alba forest developed outside temporary pools in the town of Maco (27° 51'20 "S, 64° 13'27" W), Santiago del Estero Province, Argentina. The seeds were manually extracted from the fruit and scarified by removing a small portion of the coat at the cotyledon end with nail clippers.
Germination
Four lots of twenty-five seeds were placed to germinate between paper towels moistened with 10 ml of NaCl solutions containing 0 (control), 100, 200, 300, 400, 500, 600, and 700 mM. Germination towels were rolled, covered with polythene bags to minimize water loss by evaporation, and placed vertically in a growth chamber at 25 °C with a 12- hour photoperiod (22). The seeds that emerged in the cotyledons were considered germinated. Both the percentage of germination and the mean germination time (MGT) were calculated according to Nichols and Heidecker (1996).
Growth
Seeds were placed to germinate under the conditions described in the previous item. Subsequently, 2-week-old seedlings were hydroponically cultivated in 5-liter containers with 25% Hoagland nutritive solution with the addition of 0 (control), 100, 200, 300, 400, 500, or 600 mM NaCl (20 seedlings per container).
The trial was conducted in a growth chamber (25°C, with a 12-hour photoperiod). Transpiration losses were offset by the addition of distilled water, keeping the level of solution in the containers constant.
After reaching the highest salinity, seedlings were kept under the same conditions for seven days. In that material, dry matter was determined, and the variables associated with water relations and chemical determinations were measured.
Water relations and quantification of inorganic and organic solutes
Water potential (ΨW), relative water content (RCW), and osmotic potential (ΨS), were determined using the methodology described by Silva et al. (2010). Pressure potential (ΨP) was calculated using the difference between ΨW and ΨS.
It was determined OA as the difference in osmotic potential at full turgor between control (ΨSC100) and salt stress (ΨSS100) conditions (3). It was quantified Na+ and K+ concentrations by flame spectrophotometry and determined Cl- concentration bytitration with AgNO3 (18).
Proline, soluble sugars, glycine betaine, and anthocyanin were extracted and spectrophotometrically quantified according to techniques described by Bates et al. (1973), Dubois et al. (1956), Grieve and Grattan (1983), and Krizek et al. (2016), respectively.
Each solute concentration was expressed as mmol of solute per kg of water in the tissue. Relative contribution (RC) of every solute to ΨS was estimated as a percentage of osmolarity and calculated by the following rate: RC=solute concentration (mmol kg-1 water)/solvent osmolarity (mmol kg-1 solvent), according to Silva et al. (2010).
Chlorophyll fluorescence parameters
Chlorophyll fluorescence parameters were assessed using a portable photosynthesis meter. Minimal fluorescence (Fo), was measured in 30-min dark-adapted leaves, whereas maximal fluorescence (Fm), was measured in the same leaves in full light-adapted conditions. Maximal variable fluorescence (Fv= Fm-F0) and the photochemical efficiency of PSII (Fv/ Fm) for dark-adapted leaves were also calculated from the measured parameters (21). In light-adapted leaves (for 15 min), steady state fluorescence yield (Fs´), maximal fluorescence (Fm´) after 0.8 s saturating white light pulse, and minimal fluorescence (F0´) were measured when actinic light was turned off; further calculation was made by using the equation F0´= F0/ (Fv/Fm+F0/Fm´) (28).
Non-photochemical quenching (NPQ) value due to the dissipation of absorbed light energy was determined at each saturating pulse in accordance with the equation NPQ= (Fm-Fm´)/Fm´. The coefficient for photochemical quenching, qP, which represents the fraction of open PSII reaction centre, was calculated as qP= (Fm´-Fs´)/(Fm´-F0´) (21).
Photochemical efficiency of photosystem II (ΦPSII) was calculated as followed (11): ΦPSII= (Fm´-Fs´)/Fm´.
Experimental design and statistical analysis
Each experiment was repeated at least twice. An experimental completely randomized design with four repetitions was used. For the germination tests, the experimental unit consisted of paper towels containing twenty-five seeds (four rolls of twenty-five seeds per treatment). As regards growth, water relations, and chlorophyll fluorescence tests, the experimental unit consisted of a 5-liter container accomodating twenty seedlings (four recipients per treatment). The plant physiological parameters were recorded from fully expanded second or third youngest leaves.
The germination and growth results did not meet the assumptions of ANOVA, which were analyzed with the nonparametric test Kruskal-Wallis at 0.05 confidence level. The results of water relations and chlorophyll fluorescence parameters were subjected to ANOVA, and means were compared using the Tukey test al 0.05 confidence level.
RESULTS
P. alba showed great tolerance to salinity during the germination stage (table 1, page 73).
Table 1. Percentage of germination, mean germination time (MGT), root and shoot dry weight, and root/shoot ratio in seeds and seedlings of P. alba incubated in NaCl solutions1.
Tabla 1. Porcentaje de germinación, tiempo medio de germinación (MGT), peso seco de raíces y parte aérea, y relación raíces/parte aérea en semillas y plántulas de P. alba incubadas en soluciones de NaCl1.
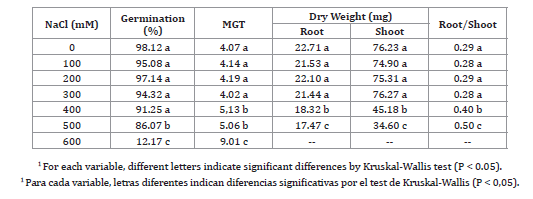
The percentage of germination was not affected by concentrations of 100, 200, 300, and 400 mM NaCl, compared with the control maintained in the order of 98%. At 500 mM NaCl, there was a small but significant decrease, reaching an average of 86%. At 600 mM, a significant inhibition of germination with an average of 12% was recorded.
At higher concentrations of NaCl, germination was completely inhibited and, though radicles emerged, cotyledons did not. Therefore, the threshold of the species in the germination stage was 600 mM NaCl.
The average germination time (TMG) was more sensitive to NaCl than the final percentage of germination, registering an increase from 400 mM NaCl, probably due to a slower imbibition of seeds as a result of the low water potential in the incubation solution.
The growth of hydroponically grown seedlings was more sensitive to salt stress than the germination process (table 1), and a different behavior was observed in shoots and roots, respectively. At 400 mM NaCl, aboveground biomass decreased 20% compared with the control, and 46% at 500 mM. On the other hand, at 400 mM NaCl, root biomass decreased 24 % compared with the control.
As a result, a significant increase was observed in the root/shoot ratio from 400 mM NaCl.
Seedlings grown at 600 mM NaCl showed chlorosis and necrosis, dying before concluding the test. Therefore, the threshold for seedling growth white carob is 500 mM NaCl.
The water potential of seedlings decreased gradually as the salt concentration in the nutrient solution increased, reaching values of -0.6 MPa in the control and -3 MPa in plants grown at 500 mM NaCl (table 2, page 74).
Table 2. Water potential (Ψw), osmotic potential (ΨS), pressure potential (ΨP), water relative content (WRC), and osmotic adjustment (OA) in seedlings of P. alba incubated in NaCl solutions1.
Tabla 2. Potencial hídrico (Ψw), potencial osmótico (ΨS), potencial de presión (ΨP), contenido relativo de agua (WRC) y ajuste osmótico (OA) en plántulas de P. alba incubadas en soluciones de NaCl1.

A similar trend was observed in the osmotic potential, with maximum values of -1.5 MPa in the control and -3.9 MPa at 500 mM NaCl. As a result, the seedlings were able to maintain cell turgor, with potential values of 0.9 MPa pressure.
The maintenance of cell turgor and WRC values (table 2, page 74) reinforces the hypothesis of the presence of OA mechanism as a response to salt stress. In agreement with these results, OA values calculated as the difference between Ψ SC 100 and Ψ SS 100 increased as the WNaCl concentration increased (table 2, page 74). It was observed OA high values of 1.42 MPa at the highest level of salinity.
The contribution of K+ to OA was declining as a result of salt stress, but it was always high, which shows a significant selectivity of this cation relative to sodium (table 3).
Table 3. Contribution of K+, Na+, Cl-, proline, total soluble sugars (TSS), and glycine betaine (GB) to osmolarity in seedlings of P. alba incubated in NaCl solutions1.
Tabla 3. Contribución de K+, Na+, Cl-, prolina, azúcares solubles totales (TSS) y glicina betaína (GB) a la osmolaridad, en plántulas de P. alba incubadas en soluciones de NaCl1.

As the contribution of K+ decreased, Na+, Cl- increased, but the contribution of the cation relative to the anion was always greater (table 3).
As regards organic solutes, the contribution of proline to osmolarity was very low, 0.94% in the control and 0.33% at the highest level of salinity (table 3).
The contribution of soluble sugars to osmolarity decreased as the NaCl concentration in the medium increased. In control, its contribution was 26.78% and, at 500 mM NaCl, it was 10.75%. The opposite was observed with glycine betaine (GB); its contribution tripled at 500 mM NaCl, compared with the control (table 3).
Fluorescence variables did not show significant differences in the range from 0 to 300 mM NaCl (table 4).
Table 4. Effects of saline stress on F v/Fm, NPQ, ΦPSII, and qP in P. alba1.
Tabla 4. Efectos del estrés salino sobre F v/Fm, NPQ, ΦPSII, and qP en P. alba1.

Higher concentrations produced a decrease in Fv/Fm, ΦPSII and qP values, whereas NPQ increased notoriously. The anthocyanin content in leaves was very sensitive to stress, increasing in all the saline concentrations (table 4).
DISCUSSION
P. alba was more tolerant to salinity in the germination stage tha in the seedling growth stage (table 1, page 73). The threshold for germination was similar to the one reported for Prosopis ruscifolia, a pioneer halophyte in the phytogeographic region of Western Chaco (22) and higher than Schinopsis lorentzii, a glycophyte part of the climax community together with P. alba. In S. lorenzii, the threshold was 300 mM, with an increase in the MGT from 100 mM NaCl (22).
Concentrations of 400 and 500 mM NaCl not only inhibited biomass production (table 1, page 73), but also increased the root/shoot ratio, resulting in more efficient water and nutrient uptake under saline stress (31).
In Western Chaco, the germination of P. alba occurs in summer, when precipitation reduces the salinity levels in the upper soil layer (23). In autumn, when precipitation ceases and the soil becomes dry, P. alba seedlings already possess a profound root system reaching the less saline soil layers, with more water available.
Salt stress can alter the physiology of plants either by an osmotic effect, reducing the water potential of the soil solution and thus limiting the water uptake by the roots, or by a specific toxic effect of ions (25).
Cellular adaptive mechanisms, such as OA, and compartmentalization of toxic ions are vital for plant tolerance to salinity (19). Under severe salt stress, P. alba was able to reduce its osmotic potential, and thus its water potential, keeping turgor constant (table 2, page 74).
Hence, the growth was not affected as a result of the water deficit caused by the low osmotic potential of the external solution.
The results of this work show that P. alba has an efficient adaptive mechanism that allows it to tolerate high levels of salinity through the maintenance of a good water status of the leaves and an effective osmotic adjustment. Table 3 (page 74) shows that he K+ involvement in osmolarity decreased with salt stress as the Na+ contribution increased, but it was always higher than the Cl- contribution.
In species that accumulate high concentrations of ions in cells, such as P. alba and halophytes, vacuoles play generally a central role in osmotic adjustment (22), allowing the compartmentalization of toxic ions such as Na+ and Cl-. Similar results were found by Villagra and Cavagnaro (2005) for P. alpataco, another salt-tolerant species.
In the cytoplasm, salinity can induce the accumulation of organic solutes compatible with cellular metabolism that, in addition to participating in the OA, act as protectors of macromolecules (22). Thus, the accumulation of ions in both vacuole and cytoplasm organic molecules allows a balance in the water relations within the cell itself.
From the organic solutes studied in this work, GB was the only one that increased its concentration as a response to increased salinity, reinforcing its contribution to osmolarity (table 3, page 74). If stored in the cytoplasm, it is normally around 10% of the cell volume.
These results differ from those observed in Prosopis strombulifera (17), which tolerates up to 700 mM NaCl; it had a significant increase in foliar concentrations of proline and soluble sugars, whereas the concentration of GB remained constant. Moreover, whereas Na+ played a key role in the OA in P. alba when compared with Cl-, in P. strombulifera the opposite was observed (17). These results suggest that Prosopis species have different levels of tolerance to salinity, and they have also developed differential physiological adaptations during the speciation process.
The Chlorophyll fluorescence analysis shows that in P. alba the photochemical stage of photosynthesis is highly tolerant to salinity, because it was only disturbed at concentrations higher than 300 mM NaCl (table 4, page 75). The Fv/Fm value is the ratio of variable fluorescence to maximal fluorescence and measures the maximum efficiency of PSII (26). This value can be used to estimate the potential efficiency of PSII by considering dark-adapted measurements (8). An increase in Fv/Fm reflects more light use efficiency in plants (16). In addition, an increase in Fv/Fm is also correlated with reduced energy loss as heat. These results are in agreement with Lauer and Ross (2016), who reported that an increase in salinity caused a reduction in Fv/Fm, indicating that there was more energy dissipated as heat (table 4, page 75).
The qP value approximates the proportion of open PSII reaction centers. In other words, qP represents the energy consumed in the photosynthesis. On the other hand, NPQ is the amount of dissipated excessive irradiation into heat. It represents the effective way in which photosynthetic organisms can accomplish the process of dissipating excessive irradiation into heat (29).
The Φ PSII value gives an estimation of the efficiency. It represents the photochemistry at different photon fluxes. There is an inverse relationship between qP and NPQ as well as between Φ PSII and NPQ (6). In this study, both ΦPSII and qP decreased with an increasing value of NPQ and were in line with the results reported by Hazrati et al. (2016). Under favourable conditions, almost all the absorbed light is consumed in photochemical reactions (15). There is a negative correlation between ΦPSII and NPQ (6). In addition, it has been reported that there is a linear relationship between Φ PSIIand CO2 absorption (20). In a study on Aloe vera, water deficit stress decreased Φ PSII and qP but increased NPQ (12). In this study, the NPQ value increased with increasing saline stress severity. The higher value of NPQ indicates the ability to mitigate the negative effects of environ mental stress at the chloroplast level, as these organelles can dissipate the excess of excitation energy (16). Similar results were found in P. alba plants exposed to NaCl stress for four days (24).
It has been shown that in photosynthetic tissues, anthocyanin acts as a “sunscreen”, protecting cells from highlight damage by absorbing ultraviolet light, thereby protecting the tissues from photoinhibition. It has also been reported that there is a positive correlation between water stress and anthocyanin biosynthesis (12). All treatments in P. alba increased the anthocyanin content in leaves, which is in agreement with the high stability of the photochemical stage of photosynthesis at high salt concentrations (less than 400 mM NaCl).
CONCLUSIONS
P. alba tolerates up to 600 mM NaCl during the germination stage and 500mM NaCl for seedling growth. Its high tolerance to salt stress is determined by its ability to adjust osmotically maintaining cell turgor through the accumulation of osmolytes, such as Na+ and GB.
The growth inhibition of the aerial part observed at the highest salt concentrations (400 and 500 mM NaCl) matches the photoinhibition of photosynthesis, as indicated by a reduced quantum yield of PSII, Φ PSII, and qP, as well as higher NPQ. The high concentration of anthocyanins constitutes a protection mechanism against adverse effects of saline stress.
1. Ashraf, M.; Harris, P. J. C. 2013. Photosynthesis under stressful environments: an overview. Photosynthetica 51: 163-190. [ Links ]
2. Bates, L. S.; Waldren, R. P.; Tear, I. D. 1973. Rapid determination of free proline for water stress studies. Plant Soil 39: 205-207. [ Links ]
3. Díaz- López, L.; Gimeno, V.; Lidón, V.; Simón, I.; Martínez, V. 2012. The tolerance of Jatropha curcas seedlings to NaCl: an ecophysiological analysis. Plant Physiol. Bioch. 54: 34-42. [ Links ]
4. Dubois, M.; Gilles, K. A.; Hamilton, J. K.; Rebers, P. A.; Smith, F. 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem. 28: 350-356. [ Links ]
5. El Keblawy, A.; Al-Rawai, A. 2005. Effect of salinity, temperature and light on germination of invasive Prosopis juliflora (Sw.) D. C. Journal of Arid Environments. 61: 555-565. [ Links ]
6. Fu, W.; Li, P.; Wu, Y. 2012. Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Sci. Hortic. 135: 45-51. [ Links ]
7. Giménez, A. M.; Moglia, J. G. 2003. Árboles del Chaco Argentino. Guía para el reconocimiento dendrológico. Facultad de Ciencias Forestales UNSE. Argentina. [ Links ]
8. Gorbe, E.; Calatayud, A. 2012. Applications of chlorophyll fluorescence imaging technique in horticultural research: a review. Sci. Hortic. 138: 24-35. [ Links ]
9. Grieve, C. M.; Grattan, S. R. 1983. Rapid assay for determination of water-soluble quaternary ammonium-compounds. Plant Soil. 70: 303-307. [ Links ]
10. Guida-Johnson, B.; Abraham, E. M.; Cony, M. A. 2017. Salinización del suelo en tierras secas irrigadas: perspectivas de restauración en Cuyo, Argentina. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 49(1): 205-215. [ Links ]
11. Guo, R.; Zhou, J.; Ren, G.; Han, W. 2013. Physiological responses of linseed seedlings to iso osmotic polyethylene glycol, salt, and alkali stresses. Agron. J. 105: 764-772. [ Links ]
12. Hazrati, S.; Tahmasebi-Sarvestani, Z.; Modarres-Sanavy, S. A. M.; Mokhtassi-Bidgoli, A.; Nicola, S. 2016. Effects of water stress and light intensity on chlorophyll fluorescence parameters and pigments of Aloe vera L. Plant Physiol. Biochem. 106: 141-148. [ Links ]
13. Jameel Al-Kahayri, M. 2002. Growth, proline accumulation, and ion content in sodium chloridestressed callus of date palm. In Vitro Cellular and Developmental Biology. 38: 79-82. [ Links ]
14. Krizek, D. T.; Kramer, G. F.; Upadhyaya, A.; Mirecki, R. M. 1993. UV-B response of cucumber seedlings grown under metal halide and high pressure sodium/deluxe lamps. Physiol. Plant. 88: 350-358. [ Links ]
15. Lauer, N.; Ross, C. 2016. Physiological and oxidative stress responses of baldcypress in response to elevated salinity: linking and identifying biomarkers of stress in keystone species. Acta Physiologiae Plantarum 38. doi:10.1007/s11738-016-2281-9. [ Links ]
16. Li, Q.; Deng, M.; Xiong, Y.; Coombes, A.; Zhao, W. 2014. Morphological and photosynthetic response to high and low irradiance of Aeschynanthus longicaulis. Sci. World. J. 14: 1-8. [ Links ]
17. Llanes, A.; Bertazza, G.; Palacio, G.; Luna, V. 2013. Different sodium salts cause different solute accumulation in the halophyte Prosopis strombulifera. Plant Biol. 15: 118-125. [ Links ]
18. Malavolta, E.; Vitti, G. C.; Oliveira, S. A. 1989. Avaliação do estado nutricional das plantas: princípios e aplicações. Associação Brasileira para a Pesquisa da Potassa e do Fosfato, Piracicaba, Brasil. [ Links ]
19. Mancarella, S.; Van Oostern, M. J.; Sanoubar, R.; Stanghellini, C.; Kondo, S.; Gianquinto, G.; Maggio, A. 2016. Leaf sodium accumulation facilitates salt stress adaptation and preserves photosystem functionality in salt stressed Ocimum basilicum. Environ. Exp. Bot. 130: 162-173. [ Links ]
20. Martins, S. C. V.; Galmes, J.; Molins, A.; Damatta, F. M. 2013. Improving the estimation of mesophyll conductance to CO2: on the role of electron transport rate correction and respiration. J. Exp. Bot. 64: 3285-3298. [ Links ]
21. Maxwell, K.; Johnson, G. N. 2000. Chlorophyll fluorescence practical guide. J. Exp. Bot. 51: 659-668. [ Links ]
22. Meloni, D. 2017. Fisiología Vegetal. Respuestas de especies leñosas al estrés salino. Editorial Universidad Nacional de Santiago del Estero. 176 p. [ Links ]
23. Meloni, D. A.; Gulotta, M. R.; Martínez, C. A. 2008. Salinity tolerance in Schinopsis quebracho colorado: seed germination, growth and metabolic responses. J. Arid Environ. 72: 1785-1792. [ Links ]
24. Meloni, D. A.; Silva, D. M.; Ledesma, R.; Bolzón. G. 2017. Nutrición mineral y fotosíntesis en plántulas de algarrobo blanco, Prosopis alba (Fabaceae) bajo estrés salino. UNED Res. J. 9:297-304. [ Links ]
25. Munns, R.; Tester, M. 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 59: 651-681. [ Links ]
26. Murchie, E. H.; Lawson, T. 2013. Chlorophyll fluorescence analysis: a guide to good practice and understanding some new applications. J. Exp. Bot. 64: 3983-3998. [ Links ]
27. Nichols, M. A.; Heidecker, W. 1996. Two approaches to the study of germination data. Proceeding of International Seed Testing Association. 33: 351-340. [ Links ]
28. Oxborough, K.; Baker, N. 1997. An instrument capable of imaging chlorophyll a fluorescence from intact leaves at very low irradiance and at cellular and subcellular levels of organization. Plant, Cell Environ. 20: 1473-1483. [ Links ]
29. Pinnola, A.; Dall’Osto, L.; Gerotto, C.; Morosinotto, T.; Bassi, R.; Alboresi, A. 2013. Zeaxanthin binds to light-harvesting complex stress-related protein to enhance nonphotochemical quenching in Physcomitrella patens. Plant Cell. 25: 3519-3534.
30. Silva, E. N.; Silva, S. L.; Viégas, R. A. 2010. The role of organic and inorganic solutes in the osmotic adjustment of drought-stressed Jatropha curcas plants. Environ. Exp. Bot. 69: 279-295. [ Links ]
31. Silveira, J. A. G.; Araújo, S. A. M.; Lima, J. P. M.; Viégas, R. A. 2009. Roots and leaves display contrasting osmotic adjustment mechanims in responses to NaCl-salinity in Atriplex nummularia. Environ. Exp. Bot. 66:1-8. [ Links ]
32. Velarde, M.; Felker, P.; Degano, C. 2003. Evaluation of Argentine and Peruvian Prosopis germoplasm for growth and seawater salinities. J. Arid Environ. 55:512-531. [ Links ]
33. Villagra, P. 2009. Germination of Prosopis argentina and P. alpataco seeds under saline conditions. J. Arid Environ. 37: 261-267. [ Links ]
34. Villagra, P. E.; Cavagnaro, J. B. 2005. Effects of salinity on the establishment and the early growth of Prosopis argentina and Prosopis alpataco seedlings in two contrasting soils: implications for their ecological success. Austral Ecol. 30:325-335. [ Links ]

