










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo
Print version ISSN 1853-8665On-line version ISSN 1853-8665
Rev. Fac. Cienc. Agrar., Univ. Nac. Cuyo vol.51 no.2 Mendoza Dec. 2019
ORIGINAL ARTICLE
Morphological variability of native maize (Zea mays L.) of the west highland of Puebla and east highland of Tlaxcala, Mexico
Variabilidad morfológica del maíz nativo (Zea mays L.) del altiplano poniente de Puebla y altiplano oriente de Tlaxcala, México
Gregorio Alvarado-Beltrán 1, Higinio López-Sánchez 2*, Amalio Santacruz-Varela 1, Abel Muñoz-Orozco 1, Ernestina Valadez-Moctezuma 3, Ma. Alejandra Gutiérrez-Espinosa 1, Pedro A. López 2, Abel Gil-Muñoz 2, Juan de Dios Guerrero-Rodríguez 2, Oswaldo R. Taboada-Gaytán 2
1 Colegio de Postgraduados. Campus Montecillo. Programa de Recursos Genéticos y Productividad. km 36,5. Carretera México-Texcoco. Montecillo. Edo. de México. C.P. 56230.
2 Colegio de Postgraduados. Campus Puebla. km. 125,5. Carretera Federal México-Puebla. Santiago Momoxpan. San Pedro Cholula. Puebla C. P. 72760. * higiniols@colpos.mx
3 Universidad Autónoma Chapingo. Departamento de Fitotecnia. km 38,5. Carretera México-Texcoco. Chapingo. Edo. de México. C. P. 56230.
Originales: Recepción: 29/11/2018 - Aceptación: 06/03/2019
ABSTRACT
The objective of the present study was to analyze the morphological variability of native maize of the western highlands of Puebla and east of Tlaxcala, Mexico, in order to, in addition to defining it, relate it to races, commercial varieties and the altitude of the localities of collection. The genetic material evaluated were 134 populations collected in 34 localities, in addition to 10 witnesses. The experiments were established in three localities, using a 12 x 12 lattice. A total of 32 variables were evaluated, of which 27 presented highly significant differences, reflecting high variability at the level of morphological characters, many of them of agronomic interest. From the analysis of variance, 16 variables were selected for use in a cluster analysis using the Modified Localization Method, which concentrated the populations in six groups, most of them in group 1, with morphological characteristics of late-cycle varieties: tall plants, with more primary ramifications of the spike, ears of greater length and diameter and with greater length and thickness of grain. The conclusions indicate that the morphological variability of the populations is not associated with the altitude of the localities where they were collected and that these have a greater relationship with the Chalqueño race, little with the Cónico race, null with the Cónico Norteño and Palomero Toluqueño races and almost null with the commercial varieties.
Keywords: Zea mays L.; Creole maize; Plant genetic resources; Germplasm; In situ conservation
RESUMEN
El objetivo del presente estudio fue analizar la variabilidad morfológica de los maíces nativos del altiplano poniente de Puebla y oriente de Tlaxcala, México, para, además de definirla, relacionarla con razas, variedades comerciales y con la altitud de las localidades de colecta. El material genético evaluado fueron 134 materiales colectados en 34 localidades, además de 10 testigos. Los experimentos se establecieron en tres localidades, mediante un Látice12 x 12. Un total de 32 variables fueron evaluadas, de las cuales 27 presentaron diferencias altamente significativas, lo que refleja alta variabilidad a nivel de caracteres morfológicos, muchos de ellos de interés agronómico. Del análisis de varianza se seleccionaron 16 variables para utilizarse en un análisis de agrupamiento mediante el Método de Localización Modificado, el cual concentró a las poblaciones en seis grupos, la mayoría de ellas en el grupo 1, con características morfológicas de variedades de ciclo tardío: plantas altas, con más ramificaciones primarias de la espiga, mazorcas de mayor longitud y diámetro y con mayor longitud y grosor de grano. Las conclusiones indican que la variabilidad morfológica de las poblaciones no se asocia con la altitud de las localidades donde fueron colectadas y que estas tienen mayor relación con la raza Chalqueño, poca con la raza Cónico, nula con las razas Cónico Norteño y Palomero Toluqueño y casi nula con las variedades comerciales.
Palabras clave: Zea mays L.; Maíz criollo; Recursos fitogenéticos; Germoplasma; Conservación in situ
INTRODUCTION
The study of the genetic diversity of maize (Zea mays L. ssp. mays) in traditional agricultural production systems is important, since it is the primary source of food and nutrition for the rural population, based mainly on native varieties, both in Mexico and in other regions, such as southeastern Europe (41). Corn is not a typical result of natural selection, it was literally invented by the ethnic groups of Mexico (46). Over time, corn has evolved in situ, thanks to the joint intervention of men and women (26); the woman improves quality characters, while the man preserves the plant and increases its yield, especially in adverse conditions for the crop, such as drought.
Corn is the most important crop in Mexico, since, according to data from the Food and Agriculture Organization, in the period 2007-2017, an average of 23.4 million t was produced in an average annual area of 7.0 million hectares (8); In addition, it is the crop that involves a greater number of farmers (around 3.2 million out of a total of four million farmers in the country).
The state of Puebla participates with an approximate production of 10% of the national total, in an area that comprises 5% of the national total sown.
In the state, the Rural Development District of Libres is the one with the highest production, since in the 2007-2017 period it produced 28.9% of the state total, with average yields of 4.7 and 2.3 t ha-1 in irrigation and in rainfall conditions, respectively. However, the area sown in rainfall conditions in the same period represents 94% (approximately 102,788 ha), while in irrigation it is only 6% (approximately 6,221 ha) (40).
In the state of Puebla, research has been carried out that describes the morphological variability of maize populations: Mixteca Alta (2), part of the plateau (9, 17), Puebla Valley (16) and humid tropics (20), without an investigation to date indicating the morphological variability of the races planted in the Rural Development District of Libres, despite its importance in maize production.
The studies that could give an idea of the races are those of Wellhausen et al. (1951) and Cervantes and Mejía (1984), who mention that populations belonging to the Chalqueño and Cónico races are planted in the Valley of Puebla, adjacent to the District.
Despite the engrained tradition of the District of Libres as a producer of maize, a fluctuation in the area (ha) planted (123,298, 118,685, 127,841, 121,254, 118,910, 118,157, 116,678, 134,366, 122,413, 120,564 and 130,646 ha) has been observed (2007-2017 period), with a significant decrease in the years 2008, 2011, 2012 and 2013 (40), a situation that has serious implications from the point of view of the preservation of the native germplasm, in addition to the fact that the presence of generations with more field experience and knowledge about plant genetic resources is decreasing due to natural aging, without replacing new generations in agricultural activities, due to the marked process of emigration.
The inadequate preservation of germplasm can lead to genetic erosion, understood as the loss of a crop, of varieties within the crop or alleles within the variety (43). The importance of preventing such erosion lies in that the genetic diversity is a support for sustainable food production systems (6, 18) and for the generation of improved varieties that increase food production that will be demanded by the increasingly growing world population, estimated to exceed 9 billion in 2050 (14).
It is therefore urgent to document and evaluate the existing plant genetic resources (1, 3), in order to propose appropriate strategies for future use in the event of climate change and the incidence of biotic and abiotic adverse factors. In this context, the present study was conducted with the objective of analyzing the morphological variability of the native maize that originated in the corridor that includes the western highlands of Puebla (Libres and Mazapiltepec) and the eastern highlands of Tlaxcala (Huamantla), to know their main morphological characteristics and the relationship they still have with the maize races and commercial varieties grown in the region, as well as establishing some relationship of morphological variation with altitudinal patterns of the localities where the seed was collected.
MATERIALS AND METHODS
Description of the study area, the localities of collection and evaluation of localities
Three ecological niches were defined based on the criteria of Muñoz (1988): Libres (NE1), Mazapiltepec (NE2) and Huamantla (NE3), in the states of Puebla and Tlaxcala, respectively. In each ecological niche, a random stratified sampling was carried out (27) in order to select a representative number of municipalities and localities for the seed collection of maize populations, obtaining a total of 13 municipalities and 34 localities (table 1, page 220).
Table 1. List of localities of seed collection.
Tabla 1. Lista de las localidades de la colecta de semilla.
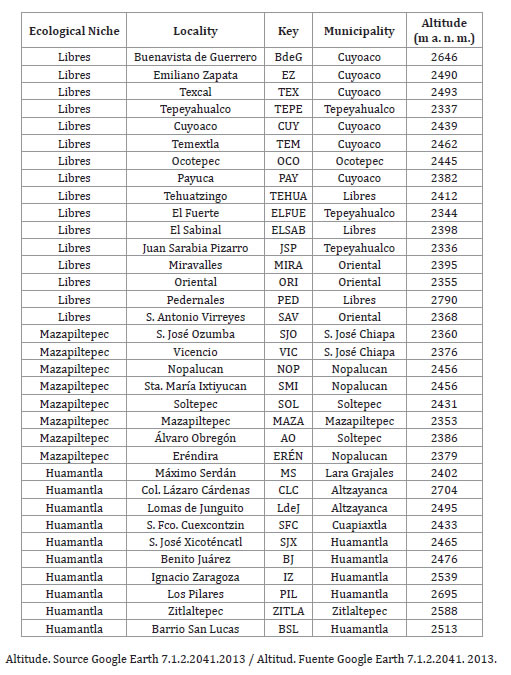
Information on the localities where the experimental material was evaluated is presented below. Buenavista de Guerrero: NE1, 19°38'07" N L, 97°30'32" W L, altitude 2646 m a. s. l., rain 516 mm, average maximum and minimum annual temperature 23 and 4°C, planting April 14th; Máximo Serdán: NE3, 9°16'36" N L, 97°48'19" W L, altitude 2402 m.a.s.l., rain 598 mm, average annual temperature 21 and 5°C, planting April 21st; Mazapiltepec: NE2, 19°07'14" N L, 97°39'56" W L, altitude 2359 m a. s. l., rain 665 ml, average annual temperature 22 and 6°C, planting May 4th. The planting in the 3 localities was in the 2007 spring summer cycle.
Genetic material
134 native populations collected in the study area were evaluated, where the racial witnesses Cónico (Criollo del Mezquital), Cónico Norteño (Zac 58), Chalqueño Crema (7CSM), Chalqueño Palomo (Col-6583), Chalqueño del Valle de Toluca (Méx-158) and Palomero Toluqueño (Méx-5) were added.
Racial witnesses were used for the purpose of relating populations to any of them. Additionally, four commercial witnesses (Sintético Serdán, 32D06, Halcón and Z60) were used to see the agronomic potential of corn populations.
Design and experimental unit
The 144 materials were evaluated in a simple 12x12 lattice design at each experimental site. The experimental unit consisted of two rows of 5 m long and 0.8 m wide, where three seeds were sown every 50 cm to later leave two plants, generating a population density of 51,754 plants per hectare.
Each experiment was surrounded by 2 rows of plants on the sides and 10 m of plants in rows at the front and at the end of the experiment. The experimental units were separated by one meter in between them.
Crop management
The fertilization was carried out in two applications: during the first weeding all the phosphorus and 1/3 of the nitrogen were applied and during the second weeding the rest of the nitrogen. In Buenavista de Guerrero the fertilization dose was 100-30-00, in Máximo Serdán 100-40-00, and for Mazapiltepec 110-50-00. Weeding was carried out periodically and herbicide was applied once (2,4-D amine) at a rate of 1 L ha-1.
Registered traits
The following traits were recorded: 1)- days to male flowering (DMF) and 2)- days to female flowering (DFF), counted from the day of planting until 50% of the plants presented dehiscent anthers and exposed stigmas, respectively, 3)- floral asynchrony (FAS), considered as the difference between DFF and DMF, recording these three traits at the experimental unit level; five random plants were subsequently selected to record the traits 4)- plant height (PH), from the stem base to the base of the spike and 5)- ear height (EH), from the base of the stem to the knot of insertion of the ear, both in cm, 6)- height index (HI), as the ratio EH/PH, 7)- number of leaves above the ear (LAE), 8)- number of primary ramifications of the spike (PRS), 9)- spike peduncle length (SPL) in cm, 10)- length of the branched spike section (LBSS) in cm, 11)- length of the spike's central branch (LPCB) in cm, 12)- total length of the spike (TLS) in cm, 13)- number of plants (NP), 14)- percentage of plants with more than one ear (PPM1E), 15)- percentage of sterile plants (PSP), 16)- plant-toharvest aspect (PHA) in visual scale 1-5, where 1 corresponds to plants with good appearance and 5 to plants with bad appearance, 17)- number of harvested ears (NHE), 18 )- ear rating (ER) in scale 1-5, where 1 corresponds to a good aspect and 5 the opposite case, 19)- field weight (FW) in g, 20)- weight of five ears in g, 21)- grain yield per hectare (GYPH) in t ha-1, using the formula:
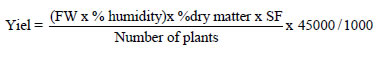
Additionally, five-plant ear characters were recorded per experimental unit: 22)- ear diameter (ED) in cm, 23)- ear length (EL) in cm, 24)- number of rows (NR), 25)- grains per row (GPR), 26)- ear length/diameter ratio (ELDR). From each ear 10 grains were taken to measure 27)- width (W), 28)- length (L) and 29)- grain thickness (GT), all of them in mm, and 30)- grain length/width ratio (GLWR); In addition, the five ears were shelled and 31) - grain weight (GW) and 32)- cob weight (CW) in g were taken to calculate 33) - the shelling factor (SF) using the ratio GW/(GW + CW).
Statistical analysis
In order to assess the stability of the expression of the traits in the three ecological niches, a combined analysis of variance between environments was performed using the PROC GLM procedure of the SAS package (Statistical Analysis System) version 9.1 (38); The linear model used was as follows:
Yijkl = µ + αi + γ j+δij + B(L)l(kj) + εijkl, i =1,2,..........,144, j =1,2,3, k=1,2, l=1,2,..........,12
where:
Yijkl= the observation of the i-th collection in the j-th environment of the k-th repetition and the l-th block
μ= the general mean and constitutes a constant common to all observations
α i= the effect of the I-th observation of the collection
γ j= the effect of the j-th environment
δ ij= the interaction effect of the i-th collection with the j-th environment
B(L) l(kj)= the effect of the l-th block nested in the k-th repetition of the j-th environment
ε ijkl= the experimental error associated with the experimental unit Yij(k).
Subsequently, in order to detect and eliminate multicollinearity, defined as the linear dependence between traits and which implies singularity (the matrix is not full range) in the Mahalanobis variance-covariance matrix, in the multivariate post-analysis, a Pearson's simple correlation matrix was elaborated, using the PROC CORR procedure of SAS, eliminating highly correlated traits (r ≥ 0.7); For example, between days of male and female flowering, which were highly correlated, only the one with the greatest biological significance was selected for later analysis. Traits were also debugged using the PROC STEPDISC procedure of SAS, applying the STEPWISE sequential method by which the discriminatory power of the traits involved in the analysis is analyzed.
STEPWISE is a multivariate technique that is applied when the traits have the property of having a normal multivariate distribution and have a common covariance matrix. This technique basically consists in the selection of a subset of informative traits or with greater discriminatory capacity to identify differences between the units of analysis.
However, this technique alone does not ensure a good selection of discriminant traits and should be complemented with other techniques and with the experience of the researcher to choose those traits that are as informative as possible of the existing variation (36, 37). Based on the F statistic, traits at 15% significance were selected.
The modified localization model or method (MLM) was used, which is a two-stage classification strategy (7, 10, 11, 12): in the initial stage, groups are defined using a hierarchical grouping method (UPGMA-unweighted pair group method with arithmetic mean), the location model is then applied to the groups formed, in which significant differences between and within groups will be verified using the Mahalanobis distance criterion, which estimates the distances and the parameters that define the groups in diversity studies (21).
The MLM has the advantage of combining all categorical traits into a single multinomial trait, which in turn can then be used in a single matrix, along with continuous traits; it is even possible to combine common grouping methods, such as Ward's method, with this technique (24).
RESULTS AND DISCUSSION
Variance analysis
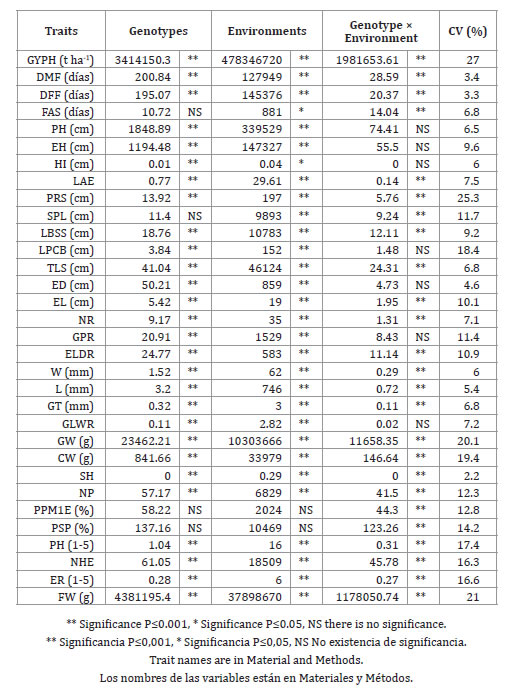
In the analysis of combined variance FAS, SPL, PPM1E and PSP did not show significant differences for genotypes, while the remaining 28 traits were significant (p ≤ 0.01), which is considered as an indicator of diversity (9, 17).
In most genetic diversity studies of native populations of maize races, high diversity is reported, mainly intrapopulation (19, 44, 45), although in some cases, such as in the Cacahuacintle race, it is minimal (13).
With respect to environments, PPM1E and PSP showed no statistical significance, FAS and HI were significant with p ≤ 0.05, while the remaining 28 traits were significant with p ≤ 0.01. As in this study, others have reported significant differences between environments in most of the traits evaluated in the native populations of maize races (2, 20).
Regarding the interaction genotypes × environments PH, EH, HI, LPCB, ED, GPR and GLWR did not show statistical significance (table 2, page 224), which implies that these traits respond in a similar way to changes in the environment between the various genotypes.
Table 2. Mean squares, level of significance and coefficients of variation of morphological data of maize landraces tested in three localities of the Libres-Mazapiltepec-Huamantla región.
Tabla 2. Cuadrados medios, niveles de significancia y coeficientes de variación para datos morfológicos de maíces nativos evaluados en tres ambientes de la región Libres-Mazapiltepec-Huamantla, 2007.
Trait selection
Multicollinearity is defined as the linear dependence between traits, its presence implies singularity (the matrix is not of full range) in the Mahalanobis variance-covariance matrix (exponential part of the multivariate normal distribution), which estimates the distances and parameters that form and define the groups in diversity studies.
One way to detect and eliminate multicollinearity is to perform a simple correlation analysis (ρ), followed by a method of trait selection.
According to Romero et al. (2002) and Mijangos-Cortés et al. (2007), of the pairs of traits that presented correlations greater than 0.75 in absolute value (r < -0.75 and r > 0.75), 9 traits with less agronomic importance were eliminated: DMF-DFF (0,98), PH-EH (0,94), DMF-TLS (-0,79), DFF-TLS (-0,78), DMF-SPL (-0,77), DFF-SPL (-0,78), SPL-TLS (0,92), LBSS-TLS (0,92) and EL-ELDR (0,79).
When applying the method of sequential selection of STEPWISE traits, the selected traits were HI, NR, ED, GLWR, DFF, PH, LAE, PRS, W, L, SH, LBSS, LPCB, EL, GT and PHA (table 3).
Table 3. Selected traits using the STEPWISE method.
Tabla 3. Variables seleccionadas utilizando el método STEPWISE.
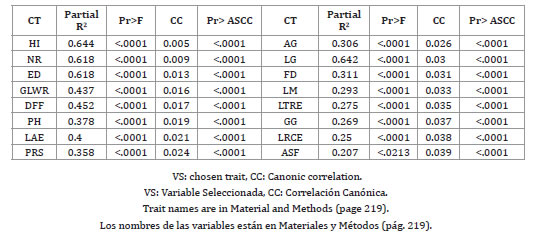
Most of the selected traits are consistent with previous diversity studies (15, 19, 30, 31, 35).
Grouping analysis
The grouping analysis, using the Modified Localization Method, in its first classification strategy defined four groups. When applying the second classification strategy, the optimal number of groups that were formed by estimating the likelihood profile and obtaining the probabilities of belonging a posteriori to each of the 144 materials evaluated with maximum restricted likelihood was determined (39).
The likelihood test ran 14 iterations before converging on the value -1098,4327, after which it assembled 6 groups (figure 1, page 226).
Figure 1. Groups and subgroups obtained by the Modified Localization Method.
Figura 1. Grupos y subgrupos ensamblados mediante el Método de Localización Modificado.
When calculating the a posteriori probabilities for each of the 144 evaluated materials, they fluctuated between 0,88 and 1,00, which represented a good classification of each of the 144 materials under study (information not shown). It was observed that there are significant differences between groups and within groups using the distance criterion of Mahalanobis (21), which allowed to form 6 groups.
Additionally, and considering the large number of populations and the variability that the materials presented within Group 1, it was considered convenient to form subgroups within this group using the Modified Localization Method (MLM), thus integrating Subgroups 1a, 1b and 1c of figure 1 (page 226).
In table 4 (page 228), the morphological characteristics of each of the groups can be observed, as follows: Group 1 was made up of 119 materials, of which 116 are native populations, two racial witnesses belonging to the Chalqueño breed (Chalqueño Crema and Chalqueño del Valle de Toluca) and an improved variety (Sintético Serdán).
Table 4. Morphological data by group of 134 landraces, four control cultivars and six control maize races grouped by the modified localization method.
Tabla 4. Datos morfológicos por grupo de 134 poblaciones nativas, 4 testigos comerciales y 6 testigos raciales ensamblados mediante el Método de Localización Modificado.
The materials of this group are late flowering, higher, greater number of PRS, occupies the third place in regards to NR, longer ears with larger diameter, longer and thicker grains, and it is the second group with wider grains. Within Group 1 is Subgroup 1a, consisting of 31 materials, of which 30 are native, plus the racial witness Chalqueño Crema. This subgroup presents the latest flowering varieties, the largest PH, and the highest number of PRS, longer ears with the largest diameter and less thick and less wide grains.
Subgroup 1b was integrated with 37 materials, 36 of which are native and one is the Chalqueño del Valle de Toluca racial witness. This subgroup is the earliest flowering of the three, the second with the highest PH, lowest number of PRS, highest NR, less wide grains and smaller thickness. Subgroup 1c was made up of 51 materials, 50 native and the witness Sintético Serdán; it is the second with the highest number of DFF and lowest PH, the second with the highest number of PRS and lowest NR, the second with L and with the highest W and with greater GT.
Group 2 was made up of 15 materials, 14 native and the Cónico racial witness. This group is the earliest flowering of all, the third with the highest PH, an average of 6,9 PRS, 15,2 rows per ear (NR). It ranks third place in terms of EL and ED, second in terms of W, third with respect to L, fourth in W, and third in GT. This group was characterized by grouping only yellow, blue and "arrocillo" grain materials, not including white color materials.
Group 3 was integrated with three commercial witnesses (Z-60, Halcón and 32D06) plus a native material, from Oriental, Puebla. These materials are late flowering, not very tall. They have a greater number of rows, longer ear length, and occupy the penultimate place in regards to PRS, second in ED, third in L, fourth in W, and last place in regards to GT.
Group 4 was made up of four native materials: 522 Cr and 524 Bl wide, from the localities of Los Pilares and Barrio San Lucas of the region of Huamantla, Tlaxcala and 25 Bl and 29 Bl wide, from Ocotepec and Payuca in the region of Libres, Puebla. These materials are of intermediate flowering and plant height. They are the ones with the lowest NR, and occupy second place in EL and fourth place in ED.
Group 5 was formed by the racial witness Cónico Norteño, which was the earliest flowering of all, the smallest in size, the penultimate in regards to NR, lower EL, the penultimate with respect to ED, lower L and second place with respect to W.
Group 6 was represented by a single material, the racial witness Palomero Toluqueño, which occupied the second place in EH, the penultimate in PH. It is the one who presented a higher NR, and occupied the penultimate place in EL, lower ED, narrower and thinner grains and had the penultimate place in L.
Based on the above, the characteristics of the majority (86.6%) of the populations are related to the Chalqueño race, few (10.4%) with the Cónico race, none with the Cónico Norteño and Palomero Toluqueño races. It is worth mentioning that the Chalqueño race, which is distributed mainly in the eastern part of the state of Mexico (15) is not considered as one of the six main races of maize in Mexico, contrary to Cónico and Cónico Norteño races that are considered the second and fourth most common, respectively (28). Only 0.7% share characteristics with commercial varieties and 2.98% do not resemble any races or commercial varieties. Therefore, the majority of the populations are of late flowering cycle, of higher plants, with more primary ramifications of the spike, ears of greater length and diameter, and with greater grain length and thickness. Even though there is a high morphological variability in the populations, it is important to highlight the low relationship with maize races, since only a relationship was found with 2, despite having conducted the study in 34 locations of three ecological niches located in the state of Puebla and Tlaxcala.
In another study carried out in several locations in the central region of Mexico, the presence of up to 8 races of maize at the local level is reported (32), which can be explained based on the fact that this is considered one of the six regions of Mexico with the highest diversity of races (34).
The greater relationship of the populations with the Chalqueño race indicates that the adaptation area has been expanded, because the in situ conservation of the genetic diversity of the crops is dynamic (31), since the extension is due to the fact that each race has a new area of potential distribution, where proper management can favor the conservation of its diversity (42).
Regarding the relationship of the groups with grain color (white, cream, blue, yellow and red) of the populations, there is no direct relationship between the two, since all were present in Subgroup 1a, and, with the exception of red, in Subgroup 1b and Subgroup 1c, which grouped 83% of the populations. In the other groups, for having fewer populations and for being integrated by the commercial witnesses, white color prevailed.
The grouping of genetic diversity through multivariate statistical methods and the modified localization method (MLM) have been used in various studies on genetic diversity of maize races. Just to mention an example, Ortiz et al. (2008) formed eight groups with populations of 8 maize races from the High Valleys of Peru.
Relationship between locations and groups
In figure 2 (page 230), it can be seen that most of the seed collection localities were integrated in Group 1, associated with native materials genetically close to the racial witnesses of the Chalqueño race.
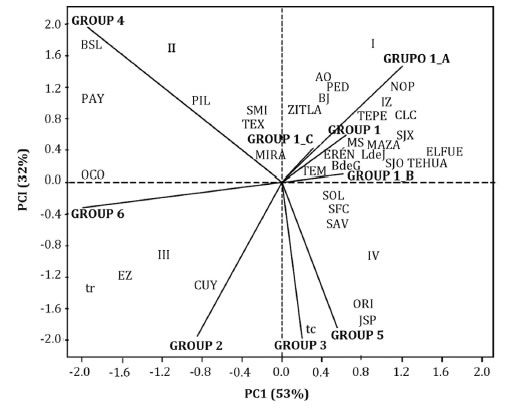
Figure 2. Relation among localities and the groups formed by the Modified Localization Method. The meaning of codes are in table 1 (page 220).
Figura 2. Relación entre las localidades y los grupos ensamblados mediante el Método de Localización Modificado. El significado de las claves está en la tabla 1 (pág. 220).
When performing the breakdown in subgroups, it is observed that Subgroup 1a was associated with the localities of Álvaro Obregón (AO, 2386 m a. s. l.), Nopalucan (NOP, 2456 m a. s. l.), Sta. María Ixtiyucan (SMI, 2456 m a. s. l.), Zitlaltepec (ZITLA, 2588 m a. s. l.), Pedernales (PED, 2790 m a. s. l.), Ignacio Zaragoza (IZ, 2539 m a. s. l.) and Benito Juárez (BJ, 2476 m a. s. l.), indicating that these localities have a closer relationship with native materials associated with the Chalqueño Crema subrace. On the other hand, Subgroup 1b was associated with the localities of Temextla (TEM, 2462 m a. s. l.), Soltepec (SOL, 2431 m a. s. l.), S. Fco. Cuexcontzin (SFC, 2433 m a. s. l.), and S. Antonio Virreyes (SAV, 2368 m a. s. l.), which indicates that these localities are mostly associated with native materials and the Chalqueño del Valle de Toluca witness; Subgroup 1c had a strong relationship with the localities of Buenavista de Guerrero (BdeG, 2619 m a. s. l.), Lomas de Junguito (LdeJ, 2495 m a. s. l.), Tehuatzingo (TEHUA, 2412 m a. s. l.), El Fuerte (ELFUE, 2344 m a. s. l.), Mazapiltepec (MAZA, 2413 m a. s. l.), Eréndira (ERÉN, 2379 m a. s. l.), S. José Xicoténcatl (SJX, 2465 m a. s. l.), Tepeyahualco (TEPE, 2337 m a. s. l.) and Col. Lázaro Cárdenas (CLC, 2704 m a. s. l.).
Group 2, which was strongly related to the Cónico racial witness, presented an association with the localities of Cuyoaco (CUY, 2439 m a. s. l.) and Emiliano Zapata (EZ, 2490 m a. s. l.).
On the other hand, Group 3 was strongly related to commercial witnesses and shared together with Group 5 the locality of Juan Sarabia Pizarro (JSP, 2336 m a. s. l.). Group 4 was associated with the localities of Barrio San Lucas (BSL, 2520 m a. s. l.), Payuca (PAY, 2382 m a. s. l.), Los Pilares (PIL, 2695 m a. s. l.), Texcal (TEX, 2493 m a. s. l.) and Miravalles (MIRA, 2395 m a. s. l.). It should be clarified that the native materials of this group had no relationship to any racial witness.
Group 5, which was related to the Cónico Norteño racial witness, but without native materials, was associated with the localities of Oriental (ORI, 2355 m a. s. l.) and part of Juan Sarabia Pizarro (JSP, 2336 m a. s. l.), while Group 6 was related to Ocotepec (OCO, 2445 m a. s. l.).
When relating the groups and subgroups to the altitude of the localities, it is observed that this ranges from 2386 to 2790 m in Subgroup 1a, from 2368 to 2462 m in Subgroup 1b, from 2337 to 2704 m in Subgroup 1c, from 2439 to 2490 m in Group 2, 2336 m in Group 3 and from 2382 to 2695 m in Group 4; in Group 5 and 6 there were no populations.
Therefore, the maximum altitudinal difference between the localities was 454 m, a difference that was not sufficient to associate the variation of the populations with an altitudinal pattern.
In other studies with a greater altitudinal difference, such as that of Diego-Flores et al. (2012) and that of Mercer et al. (2008), a relationship was found between the origin of the populations and the altitudinal pattern of evaluation.
Romero et al. (2002) mention that one of the effects of altitude on populations is associated with the temperature it generates and the effect on the regulation of genes responsible for flowering. The same authors, in a very large study on allelic diversity in flowering time in maize and its local adaptation, report 366 genes with significant association to altitude and 881 and 883 genes with significant association at days of female and male flowering, respectively.
CONCLUSIONS
There is a wide morphological variability among native maize grown in the study area, based on the statistical differences between vegetative, reproductive and yield traits, and in the 6 groups and the 3 subgroups of group 1 where the populations were located.
Most populations have vegetative, reproductive and yield characteristics typical of long or late flowering-cycle plants: tall plants, with more primary ramifications of the spike, ears of greater length and diameter, and with greater grain length and thickness, which is why they were mainly related to the Chalqueño race, to a lesser extent with the Cónico race, minimal or almost null with the commercial varieties and null with the Cónico Norteño and Palomero Toluqueño races.
The morphological variation of the populations is not associated with an altitudinal pattern, due to the little altitudinal difference between the localities.
1. Alcalá-Rico, J. S. G. J.; Espinoza-Velázquez, J.; López-Benítez, A.; Borrego-Escalante, F.; Rodriguez- Herrera, R.; Hernández-Martínez, R. 2019. Agronomic performance of maize (Zea mays L.) populations segregating the polyembryony mutant. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. [ Links ]
2. Ángeles-Gaspar, E.; Ortiz-Torres, E.; López, P. A.; López-Romero, G. 2010. Caracterización y rendimiento de poblaciones de maíz nativas de Molcaxac, Puebla. Revista Fitotecnia Mexicana. 33(4): 287-296. [ Links ]
3. Carloni, E.; López Colomba, E.; Ribotta, A.; Quiroga, M.; Tommasino, E.; Griffa, S.; Grunberg, K. 2018. Analysis of genetic variability in vitro regenerated buffelgrass plants through ISSR molecular markers. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 50(2): 1-13. [ Links ]
4. Cervantes, S. T.; Mejía, A. H. 1984. Maíces nativos del área del Plan Puebla: recolección de plasma germinal y evaluación del grupo tardío. Revista Chapingo. 9: 64-71. [ Links ]
5. Diego-Flores, P.; Chávez-Serbia, J. L.; Carrillo-Rodríguez, J. C.; Castillo-González, F. 2012. Variabilidad en poblaciones de maíz nativo de la Mixteca Baja Oaxaqueña, México. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 44(1): 157-171. [ Links ]
6. Espósito, M. A.; Gatti, I.; Bermejo, C. J.; Cointry, E. L. (en prensa). Evaluation of a lentil collection (Lens culinaris Medik) using morphological traits and digital phenotyping. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. [ Links ]
7. Everitt, B. S.; Dunn, G. 2001. Applied Multivariate Data Analysis. 2nd edition. Edward Arnold. London. UK. 342 p. [ Links ]
8. FAOSTAT. 2019. FAO Statistical Databases. Food and Agriculture Organization of the United Nations. Disponible en: http://faostat.fao.org/. Consultado Febrero de 2019. [ Links ]
9. Flores-Pérez, L.; López, P. A.; Gil-Muñoz, A.; Santacruz-Varela, A.; Chávez-Servia, J. L. 2015. Variación intra-racial de maíces nativos del altiplano de Puebla, México. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 47(1): 1-17. [ Links ]
10. Franco, J.; Crossa, J.; Villaseñor, S. J.; Taba, S.; Eberhart, A. 1998. Classifying genetic resources by categorical and continuous variables. Crop Science. 38: 1688-1696. [ Links ]
11. Franco, J.; Crossa, J.; Taba, S.; Shands, H. A. 2005. Sampling strategy for conserving genetic diversity when forming core subsets. Crop Science. 45(3): 1035-1044. [ Links ]
12. Goncalves, L. S. A.; Rodrigues, R.; Amaral, J. A. D.; Karasawa, M.; Sudré, C. P. 2009. Heirloom tomato gene bank: assessing genetic divergence based on morphological, agronomic and molecular data using a Ward-modified location model. Genetics and Molecular Research. 8(1): 364-374. [ Links ]
13. González, H. A.; Pérez, L. D. de J.; Domínguez, L. A.; Franco, M. O.; Balbuena, M. A.; Ramos, M. A.; Sahagún, C. J. 2008. Variabilidad genética, diversidad fenotípica e identificación de poblaciones sobresalientes de maíz Cacahuacintle. Ciencia Ergo Sum. 15(3): 297-305. [ Links ]
14. Govindaraj, M.; Vetriventhan, M.; Srinivasan. 2015. Importance of genetic diversity assessment in crop plants and its recent advances: an overview of its analytical perspectives. Genetics Research International. Article ID 431487. 14 p. Available in: http://dx.doi. org/10.1155/2015/431487. [ Links ]
15. Herrera-Cabrera, B. E.; Castillo-González, F.; Sánchez-González, J. J.; Hernández-Casillas, J. M.; Ortega-Pazkca, R. A.; Major-Godman, M. 2004. Diversidad del maíz Chalqueño. Agrociencia. 38(2): 191-206. [ Links ]
16. Hortelano, S. R. R.; Gil, M. A.; Santacruz, V. A.; Miranda, C. S.; Córdova, T. L. 2008. Diversidad morfológica de maíces nativos del Valle de Puebla. Agricultura Técnica Mexicana. 34: 189-200. [ Links ]
17. Hortelano, S. R. R.; Gil, M. A.; Santacruz, V. A.; López, S. H.; López, P. A.; Miranda, C. S. 2012. Diversidad fenotípica de maíces nativos del Altiplano Centro-Oriente del estado de Puebla, México. Revista Fitotecnia Mexicana. 35(2): 97-109. [ Links ]
18. Lobo, A.; M. Medina, C. C. I. 2009. Conservación de recursos genéticos de la agrobiodiversidad como apoyo al desarrollo de sistemas de producción sostenibles. Revista Corpoica-Ciencia y Tecnología Agropecuaria. 10(1): 33-42. [ Links ]
19. López, R. G.; Santacruz, V. A.; Muñoz, O. A.; Castillo, G. F.; Córdova, T. L.; Vaquera, H. H. 2010. Perfil isoenzimático de maíces nativos del Istmo de Tehuantepec, Oaxaca, México. I. Caracterización de grupos. Revista Fitotecnia Mexicana. 33(1): 1-10. [ Links ]
20. López-Morales, F.; Taboada-Gaytán, O. R.; Gil-Muñoz. A.; López, P. A.; Reyes-López, D. 2014. Morphological diversity of native maize in the humid tropics of Puebla, Mexico. Tropical and Subtropical Agroecosystems. 17: 19-31. [ Links ]
21. Mahalanobis, P. C. 1936. On the generalized distance in statistics. Proceedings of the National Institute of Sciences of India. 2: 49-55. [ Links ]
22. Mercer, K.; Martínez-Vásquez, A.; Perales, H. R. 2008. Asymmetrical local adaptation of maize landraces along an altitudinal gradient. Evolutionary Applications: 1(3): 489-500. doi:10.1111/j.1752-4571.2008.00038.x [ Links ]
23. Mijangos-Cortés, J. O.; Corona-Torres, T.; Espinosa-Victoria, D.; Muñoz-Orozco, A.; Romero-Peñaloza, J. ; Santacruz-Varela, A. 2007. Differentiation among maize (Zea mays L.) landraces from the Tarasca Mountain Chain, Michoacan, Mexico and the Chalqueño complex. Genetic Resources and Crop Evolution. 54: 309-325. [ Links ]
24. Mohammadi, S. A.; Prasanna, B. M. 2003. Analysis of genetic diversity in crop plants-salient statistical tools and considerations. Crop Science. 43: 1235-1248. [ Links ]
25. Muñoz, O. A. 1988. Resistencia a factores adversos y mejoramiento de los patrones etnofitogenéticos de la Mixteca. In: Memoria del seminario Cómo Aumentar la Producción Agropecuaria y Forestal en la Región Mixteca Oaxaqueña. Tomo II. Muñoz, O. A.; Dimas, Ch B. (comps). Colegio de Postgraduados. Tiltepec. Oax. 537-548. [ Links ]
26. Muñoz, O. A. 2005. Centli-Maíz. Prehistoria e Historia, diversidad, potencial, origen genético y geográfico. Colegio de Postgraduados. Montecillo. Edo. de México. 210 p. [ Links ]
27. Navarro, A. J. 2004. Introducción al diseño y análisis del muestreo de poblaciones finitas. In: Técnicas de muestreo para manejadores de recursos naturales. Bautista-Zúñiga, F.; Delfín-González, H.; Palacio-Prieto, J. L.; Delgado-Carranza, M. del C. (eds). UNAM, UAY, CONACYT, INE. México. p. 19-69. Available in: http://www.ine.gob.mx/ueajei / publicaciones/consultaPublicacion.html?id_pub=429. Consultado Enero 2012. [ Links ]
28. Orozco-Ramírez, Q.; Perales, H.; Hijmans, R. J. 2017. Geographical distribution and diversity of maize (Zea mays L. subsp. mays) races in Mexico. Genetic Resources and Crop Evolution 64: 855-865. DOI 10.1007/s10722-016-0405-0. [ Links ]
29. Ortiz, R.; Crossa, J.; Franco, J.; Sevilla R.; Burgueño J. 2008. Classification of Peruvian highland maize races using plant traits. Genetic Resources and Crop Evolution. 55: 151-162. [ Links ]
30. Perales, R. H.; Brush, S. B.; Qualset, C. Q. 2003a. Dynamic management of maize landraces in Central Mexico. Economic Botany. 57(1): 21-34. [ Links ]
31. Perales, R. H.; Brush, S. B.; Qualset, C. Q. 2003b. Landraces of maize in Central Mexico: an altitudinal transect. Economic Botany. 57(1): 7-20. [ Links ]
32. Perales, H. H.; Golicher, D. 2014. Mapping the diversity of maize races in Mexico. PLoS ONE 9(12): e114657. doi:10.1371/journal.pone.0114657. [ Links ]
33. Romero, P. J.; Castillo, G. F.; Ortega, P. R. 2002. Cruzas de poblaciones nativas de maíz de la raza Chalqueño. II. Grupos genéticos, divergencia y heterosis. Revista Fitotecnia Mexicana. 25: 107-115. [ Links ]
34. Romero, N. J. A.; Wilcox, M.; Burgueño, J.; Romay, C.; Swarts, K.; Trachsel, S.; Preciado, E.; Terrón, A.; Vallejo, D.; H.; Vidal, V.; Ortega, A.; Espinoza, B. A.; Gómez M., N. O.; Ortiz-Monasterio, I.; San Vicente, F.; Guadarrama E. A.; Atlin, G.; Wenzl, P.; Hearne, S.; Buckler, E. S. 2017. A study of allelic diversity underlying flowering-time adaptation in maize landraces. Nature Genetics. 49: 476-480. https://doi.org/10.1038/ng.3784 [ Links ]
35. Santacruz-Varela, A. 2001. Genetic diversity of North American popcorn and its relationship with Mexican and South American popcorns. Ph.D. Diss. Iowa State. University. Ames. IA. 326 p. [ Links ]
36. SAS Institute Inc. 2004. SAS/STAT® 9.1 User's Guide. Cary, NC: SAS Institute Inc. [ Links ]
37. SAS Institute Inc. 2008. SAS/STAT® 9.1 User's Guide. Cary, NC: SAS Institute Inc. [ Links ]
38. Searle, S. R. 1989. Variance components -some history and a summary account of estimation methods. Journal of Animal Breeding and Genetics. 106: 41-62. [ Links ]
39. SIAP. 2010. Anuario Estadístico de la Producción Agrícola. Estado de Puebla. Servicio de Información Agroalimentaria y Pesquera. SAGARPA. México, D. F. Available in: http:// www.siap.gob.mx/aagricola_siap/icultivo/index.jsp. Consultado Mayo 2010. [ Links ]
40. Simeonovska, E.; Gadžo, D.; Ovovi, Z.; Murariu, D.; Kondic, D.; Mandic, D.; Fetahu, D.; Šarevi, H.; Elezi, F.; Prodanovi, S.; Rozman, L.; Veverita, E.; Kolev, K.; Antonova, N.; Thörn, E. 2013. Collecting local landraces of maize and cereals in south Eastern Europe during 2009 and 2010. Romanian Agricultural Research. 30: 37-43.
41. Ureta, C.; Martínez-Meyer, E.; Perales, H. R.; Álvarez-Buylla, E. R. 2012. Projecting the effects climate change on the distribution of maize races and their wild relatives in Mexico. Global Change Biology. 18(3): 1073-1082. Available in: https://doi.org/10.1111/ j.1365-2486.2011.02607.x [ Links ]
42. Van de Wouw, M.; Kik, Ch.; van Hintum, T.; van Treuren, R.; Visser, B. 2009. Genetic erosion in crops: concept, research results and challenges. Plant Genetic Resources: characterization and utilization. 8(1): 1-15. [ Links ]
43. Vázquez, P. F. R. 2003. Envejecer entre los cultivos del campo mexicano. Memorias del 51 Congreso Internacional de Americanistas. 14 al 28 de julio de 2003. Santiago de Chile. 12 p. [ Links ]
44. Vega-Alvarez, I.; Santacruz-Varela, A.; Rocandio-Rodríguez, M.; Córdova-Téllez L.; López-Sánchez, H.; Muñoz-Orozco, A.; Hernández-Bautista, A. 2017. Genetic diversity and structure of native maize races from northwestern Mexico. Pesq. Agrop. Bras. 52(11): 1023-1032. DOI 10.1590/S0100-204X2017001100008. [ Links ]
45. Warman, A. 1988. La Historia de un bastardo: maíz y capitalismo. Fondo de Cultura Económica. México. D. F. 283 p. [ Links ]
46. Wellhausen, E. J.; Roberts, L. M.; Hernández, X. E.; Mangelsdorf, P. C. 1951. Razas de maíz en México: su origen, características y distribución. Folleto Técnico No. 5. Oficina de Estudios Especiales. Secretaría de Agricultura y Ganadería. México. D. F. 237 p. [ Links ]
ACKNOWLEDGEMENTS
To the Priority Research Line 6: Conservation and Improvement of Plant Genetic Resources of the Colegio de Postgraduados, for the funding provided for conducting the research.

