











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Canola is a source for the production of edible oil and biofuel. In Southern Brazil, grain production fields have shown increases in the cultivation of canola (Brassica napus var. oleifera (Mönch) Delile) in winter. The state of Rio Grande do Sul (RS) is the largest canola producer in the country, being responsible for about 85% of the total Brazilian production, with average grain yields of 1,422 kg ha-1 (8. This productivity is much lower than that obtained in countries that have grain yields of about 4,000 kg ha-1, such as Belgium, Germany, Denmark, and Chile 13. This difference may be associated with cultivation technologies such as better pest control (diseases, insects, and weeds), harvesting technology, superior genotypes, and climatic conditions that favor plant performance.
One of the factors that canola producers are facing is the interference caused by weeds, since there are few herbicides registered for this crop in Brazil. Among the main weed species infesting canola are turnip (Lolium multiflorum Lam.) and black oat (Avena strigosa Schreb.), which are very competitive for water, light, CO2, and nutrients. Furthermore, in many cases, these weeds are resistant to ALS, ACCase, or EPSPs, inhibiting herbicides 1,10,30 and making them harder to control 5.
Canola causes allelopathic suppression on germination and seedling development of both weeds and mulching species when cultivated under no-tillage; this agronomic trait allows for better competitive ability. Thus, it is an excellent alternative to manage weeds when cultivated in succession to summer crops, such as maize, soybean, and common bean 18,32. Furthermore, there is strong evidence of differences in competitive ability among both varieties of the same crop and among distinct genotypes of wild species 3,19.
Black oat is sown as mulching for the production of grains/seeds and for animal feed; however, in many cases, natural reseeding occurs, and that is when it becomes a weed in winter crops. The extract of A. strigosa may have an allelopathic effect on some crops and weeds, but the first studies were inconclusive 30. Black oats are known to perform better than barley (Hordeum vulgare L.) in competition 16, as evidenced by its ability to suppress other crops. The competition dynamics between black oat and canola is not yet well defined in subtropical production systems, such as those found in Southern Brazil and neighboring regions of Uruguay, Argentina, Paraguay, and Chile.
Weeds, if not properly controlled, reduce both grain yield and quality, in addition to hosting insects and diseases and hampering mechanized harvest 17. Experiments with replacement series are used to assess weed interference on crops by helping to understand how distinct plant species perform when grown in a community 26. The development of strategies to minimize the effects of weeds on crops is essential in the agriculture since there is an environmental need to reduce pesticide applications while maintaining high grain yields to guarantee the economic sustainability of farming.
Therefore, the objective of this study was to assess the competitive ability of canola hybrids in competition with black oat in a subtropical environment.
Material and methods
The experiments were conducted in a greenhouse in the agricultural year 2015/16. Experimental units consisted of 8 L plastic pots filled with Humic Oxisol 31, previously corrected and fertilized according to the technical recommendation for canola 36 and watered to maintain field capacity. A completely randomized design was used, as the greenhouse was considered sufficiently homogeneous, with four replications. The canola hybrids ‘Hyola 61,’ ‘Hyola 76,’ ‘Hyola 433,’ and ‘Hyola 571’ were tested individually for their competitive performance with black oat. There were no attacks, pests, or diseases on the plants throughout the study.
Preliminary experiments were carried out, both for canola hybrids and for black oat grown in monoculture to determine the minimum plant density in which the final dry mass becomes constant. For this, densities of 1, 2, 4, 8, 16, 24, 32, 40, 48, 56, and 64 plants pot-1 were used (equivalent to 24, 48, 96, 192, 384, 576, 768, 960, 1,152, 1,344, and 1,536 plants m-2, respectively). The constant final production was obtained with 20 plants pot-1, for all hybrids tested in competition with the weed, equivalent to 481 plants m-2 (data not shown). The constant final production obtained in the preliminary experiment was higher than what is usually found in field conditions; this may occur because the objective in the field is the production of grains and not competition or the accumulation of dry mass, as the case in the greenhouse.
In the second phase of experimentation, four experiments with replacement series were installed to assess the competitiveness of canola hybrids with black oat, in different combinations, with distinct plant proportions (20:0, 15:5, 10:10, 5:15, and 0:20 plants pot-1, or 100%:0%;75%:25%; 50%:50%; 25%:75%, and 0%:100%), maintaining the total population of plants at 20 plants pot-1.
Physiological variables were assessed using an infrared gas analyzer (IRGA), model LCA4 Pro SD (ADC Bioscience, UK), 55 days after emergence (DAE) to evaluate a completely developed leaf in the middle third of canola plants; for black oats, readings were performed on a fully developed leaf in the top third of the plant. The photosynthesis rate (A: μmol m-2 s-1), internal CO2 concentration (Ci: μmol mol-1), and transpiration rate (E: mol H2O m m-2 s-1) were evaluated.
After physiological assessment, leaf area (AF: m2 m-2) was assessed with a portable leaf area meter (model CI-203 BioScience). Plants were also collected, packed in paper bags, and dried in an air circulating oven at 65oC until a constant mass, for later weighing and determination of dry mass (DM: g m-2).
The expected values are indicated in the graphs as dotted straight lines, while the observed values are shown with solid regression lines. This analysis was based on the construction of a diagram based on the variations in relative (PR) and total (PRT) productivity, following this equation with the two tested species: PR(A)= PA AMIX/ AMON, where PA is the proportion of species A in the mixture; Amix is the productivity of species A in the mixture; and Amon is the productivity of species A in monoculture. When PRA was a straight line (observed = expected), the competitive ability of both species was equal. A PR below that expected indicated harm to the growth of the species, and a PR above that expected indicated a benefit to the growth of the species (4).
For the PRT values, the equation was PRT= PRA + PRB. When PRT = 100%, there was competition for the same resources; values of PRT > 100% indicated that competition was avoided, and values of PRT < 100% indicated mutual damage for growth 26. Differences between treatments for estimated and observed values of PR and PRT were compared by their 95% confidence intervals of the means (colored bands around the regressions) and were considered distinct when the estimated values were not in the range of the 95% confidence intervals for the observed means 7.
The relative competitiveness index (CR), relative clustering coefficient (K), and aggressiveness (A) were obtained, as well as their standard errors. CR was the comparative growth of a species in relation to the other under competition. K indicated the relative dominance of one species over another, and A determined which species was more aggressive in the competition. Species 1 was more competitive than species 2 when CR1 > 1, K1 > K2, and A > 0, and vice-versa 11. To calculate these indices, the 50%:50% species proportion was used, with the following equations: CR1 = PR1 / PR2; CR2 = PR2 / PR1; K1 = PR1 /(100 R1); K2 = PR2 /(100 R2); A1 = PR1 - PR2; and A2 = PR2 - PR1, as suggested by other studies. The significances of CR and A were measured by the one-way T-test and were considered significant when they differed (p ≤ 0.05) from 1 and 0, respectively. Differences between K1 and K2 were compared by the two-way T-test with Welch criteria and considered significant when p ≤ 0.05. All graphics and coefficients were obtained using the statistical software environment ‘R’ 27.
Results and discussion
The relative yield of the photosynthesis rate (A) did not differ consistently from the expected values in most situations (Figure 1).

Figure 1: Figura 1: Relative yield (%) for photosynthesis rate (A) of canola (●) hybrids (A: Hyola 61; B: Hyola 76; C: Hyola 433; D: Hyola 531) and black oat (▲) and for the whole plant community (■) as a function of plant proportion. Rendimiento relativo (%) para la tasa de fotosíntesis (A) de híbridos de canola (●) (A: Hyola 61; B: Hyola 76; C: Hyola 433; D: Hyola 531) y avena negra (▲) y para toda la comunidad de plantas (■) en proporción de plantas.
Only hybrids Hyola 61 and 76 presented lower photosynthetic rates when in smaller proportions in the mixture (less than 25% of the plants in the stand; Figures 1A; 1B, page 122). The photosynthetic behavior of Hyola 433 (Figure 1C, page 122) did not differ from the expected values (dotted lines), while Hyola 571 (Figure 1D, page 122) showed increased competition levels with black oats when their plant stands were equal. In this situation, the competition for the same environmental resources intensified.
There was no difference in the photosynthesis rate when canola, under any competition level, was compared to the respective control treatment free of competition, and it ranged between 15.68 and 21.04 μmol m-2 s-1 (Table 1).
Table 1: Tabla 1: Physiological responses of canola (Brassica napus) hybrids under competition with black oat (Avena strigosa), in terms of photosynthesis rate (A), internal CO2 concentration (Ci), and transpiration rate (E) in experiments conducted by the substitutive series method. Respuestas fisiológicas de híbridos de canola (Brassica napus) en competencia con avena negra (Avena strigosa), en términos de tasa de fotosíntesis (A), concentración interna de CO2 (Ci) y tasa de transpiración (E) en experimentos realizados por el método de serie sustitutiva.

* Means differ from the respective control treatment (T) according to Dunnett’s test (p ≤ 0.05).
* Las medias difieren del tratamiento control respectivo (T) según la prueba de Dunnett (p ≤ 0,05).
Similar results were also observed by Naderikharaji et al. (2008), when observing the effect of radiance and drought stress on the photosynthetic rate of four canola cultivars, respectively. The results showed that the values of net photosynthesis were similar even with the use of different varieties.
The relative yield based on the CO2 concentration in the leaf mesophyll (Ci) presented narrower confidence intervals (Figure 2, page 124) compared to the photosynthesis rate (Figure 1, page 122).

Figure 2: Figura 2: Relative yield (%) for the internal CO2 concentration (Ci) of canola (●) hybrids (A: Hyola 61; B: Hyola 76; C: Hyola 433; D: Hyola 531), black oat (▲) and for the whole plant community (■) as a function of plant proportion. Rendimiento relativo (%) para la concentración interna de CO2 (Ci) de híbridos de canola (●) (A: Hyola 61; B: Hyola 76; C: Hyola 433; D: Hyola 531); avena negra (▲) y para toda la comunidad de plantas (■) en función de la proporción de plantas.
There were no consistent changes in relative Ci yield among cultivars. Hyola 61 had a lower performance than the competitor when it represented 75% of the plant stand (Figure 2A, page 124), and Hyola 76 performed better than the competitor in the same situation (Figure 2B, page 124). Hyola 433 was harmed when the plant stand equaled the competitor (Figure 2C, page 124), while Hyola 571 was harmed when it represented only 25% of the stand (Figure 2D, page 124).
The internal CO2 concentration of the leaf (Ci) depends greatly on A 35; the higher the A, the lower the Ci is expected to be, as the CO2 is consumed at higher rates by photosynthesis. The photosynthesis rate was barely changed by competition levels (Figure 1, page 122); thus, it was expected that Ci would not be changed (Figure 2). The narrow confidence interval may indicate that Ci is a reliable marker for inferences about relative physiological yield 28. The CO2 concentration in canola leaves ranged between 212.19 and 277.00 μmol mol-1 (Table 1, page 123).
Values of 390 ppm (390 μmol mol-1) can be considered as usual CO2 concentrations in free air in the Southern hemisphere 30; thus, between 113 and 178 ppm CO2 were consistently consumed by the photosynthetic metabolism of canola (Table 1, page 123). In this study, competition did not affect the physiological metabolism of the plant. One possible explanation for this lack of change in the studied proportions can be attributed to the rapid carboxylation and subsequent transformation of CO2 into glucose, which can be evidenced by the increase in photosynthetic rate. Similar behavior was observed in competitive studies of soybeans with barnyard grass 6.
Competition also did not affect the relative plant yields based on transpiration rates (E); as for most situations, no effects of competition were reported (Figure 3, page 125).

Figure 3: Figura 3: Relative yield (%) for transpiration rate (E) of canola (●) hybrids (A: Hyola 61; B: Hyola 76; C: Hyola 433; D : Hyola 571); black oat (▲) and for the whole plant community (■) as a function of plant proportion. Rendimiento relativo (%) de la tasa de transpiración (E) de híbridos de canola (●) (A: Hyola 61; B: Hyola 76; C: Hyola 433; D: Hyola 571); avena negra (▲) y para toda la comunidad de plantas (■) en función de la proporción de plantas.
There was, however, a consistent decrease in the physiology of the competitor species (black oat) when it represented 75% of the plant stand (Figures 3A; 3B; 3D, page 125).
From the point of view of the competitor, this is an indication that a few plants of canola may trigger competition sensing in black oats; in other words, black oat is capable of noticing the competition 16, but the data in this work suggest that it is not capable of overcoming canola easily in competition. Canola, however, may be harmed in the competition process if some environmental resource or a mismanaged cultural practice promotes advantages to the competitor 25. Absolute transpiration rates averaged 4.54-6.69 mol H2O m-2 s-1 (Table 1, page 123).
In contrast to the physiological parameters (Figure 1, page 122, Figure 2, page 124 and Figure 3), the relative yield of canola based on leaf area (Figure 4, page 126) was affected by competition for all genotypes.
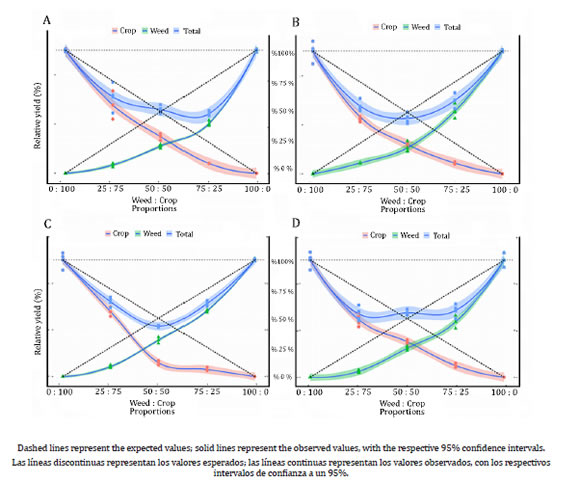
Figure 4: Figura 4: Relative yield (%) for leaf area (AF) of canola (●) hybrids (A: Hyola 61; B: Hyola 76; C: Hyola 433; D: Hyola 531); black oat (▲) and for the whole plant community (■) as a function of plant proportion. Rendimiento relativo (%) para el área foliar (AF) de híbridos de canola (●) (A: Hyola 61; B: Hyola 76; C: Hyola 433; D: Hyola 531); avena negra (▲) y para toda la comunidad de plantas (■) en función de la proporción de plantas.
The competitor (black oat) was most seriously affected by the crop (canola) when the latter represented between 50 and 75% of the plant stand. Canola was more sensitive to competition when it represented 25-50% of plant stand (Figure 4, page 126). Both species were more harmed by compatible combinations. Decreases in leaf area yield ranged between 22 and 40% for the competitor in the more harmful plant combinations. Similarly, the most damaging competition levels for canola were at complementary (opposite) densities (Figure 4, page 126).
The community behavior for leaf area showed strong competition for the same resources since both competitors were seriously harmed in their interaction (Figure 4).
The community lost between 44 and 57% of the total leaf area canopy when plants were grown together, compared to the monoculture stands. Competition for essential resources is also an important aspect of integrated weed management. The canola crop represents a tool capable of reducing the presence of some spontaneous species and, through crop rotation, delays the emergence of resistant weeds due to allelopathic effects 22. Winter cereals, when competing for resources, invest their photoassimilates in the formation of stalks (height) as a strategy to capture more luminosity; consequently, there is a decrease in the development of leaf area and dry mass 29.
Leaf area (Table 2, page 127) was presented in terms of m2 of leaf area per m2 of soil to correspond to leaf area index (LAI), a growth analysis parameter 24.
Table 2: Tabla 2: Biometrical responses of canola (Brassica napus) hybrids under competition with black oat (Avena strigosa), in terms of leaf area (AF) and dry matter (DM) in experiments conducted by the substitutive series method. Respuestas biométricas de los híbridos de canola (Brassica napus) en competencia con avena negra (Avena strigosa), en términos de área foliar (AF) y materia seca (DM) en experimentos realizados por el método de serie sustitutiva.

* Means differ from the respective control treatment (T) according to Dunnett’s test (p ≤ 0.05).
* Las medias difieren del tratamiento control respectivo (T) según la prueba de Dunnett (p ≤ 0,05).
The leaf area index differed for all treatments and genotypes compared to their respective control treatments. Hyola 61, 76, and 433 averaged LAI’s of 6.16, while Hyola 571 averaged an LAI of 2.31. Overall, leaf area decreases reached about 65% when all canola varieties were faced with the most intense competition levels with black oats (Table 2, page 127).
The mass accumulation of canola was consistent with the final grain production 20; thus, any weed species interference will result in loss of yield. With the development of more competitive hybrids, it has been possible to use more aggressive species, obtaining a higher final grain yield in the presence of weeds compared to less competitive hybrids 33. In field conditions, the use of more competitive hybrids can decrease up to 50% of the accumulation of biomass in weeds 21. In competitive studies with ryegrass (Lolium multiflorum), which is also a Poaceae, the behavior between canola varieties determined the survival strategy generated by the competition 34; for example, the hybrid Hyola 571 invests in taller plants, allowing greater capture of solar radiation 22. In this study, the difference in architecture between species showed greater differentials, where the canola was more efficient in capturing light when compared to some Poaceae species 33,36.
For dry mass (Figure 5, page 128), the relative yield presented similar behavior as that reported for leaf area, as these two variables were co-dependent.

Figure 5: Figura 5: Relative yield (%) for dry mass (DM) of canola (●) hybrids (A: Hyola 61; B: Hyola 76; C: Hyola 433; D: Hyola 531); black oat (▲) and for the whole plant community (■) as a function of plant proportion. Rendimiento relativo (%) para la masa seca (MS) de híbridos de canola (●) (A: Hyola 61; B: Hyola 76; C: Hyola 433; D: Hyola 531); avena negra (▲) y para el toda la comunidad de plantas (■) en función de la proporción de plantas.
In general terms, canola was harmed with any degree of competition with black oats, with more intense impacts being reported when it represented 25-50% of the plant stand (Figure 5 A-D, page 128).
For all genotypes, 50% of the environment (overall) relative yield was decreased by competition, proving that the interaction of canola and black oats is harmful for both species. Plant dry mass accumulation (Table 2) also followed that reported for leaf area, where all competition levels for all canola genotypes differed from their respective control treatment according to Dunnett’s test.
Studies on the capacity of dry matter accumulation are important tools to determine the impact of competition between crops and weeds. Photosynthesis is directly related to this production of dry mass, since the productive capacity depends on the photosynthetic efficiency of the crop, combined with the speed with which resources are captured 14.
While the stress levels imposed by competition were not enough to decrease photosynthesis rates (Figure 1, page 122), they did decrease leaf expansion (Figure 4, page 126). Physiologically, lower stress levels tended to decrease photosynthesis while maintaining the leaf expansion rate. In contrast, intense stress also decreased leaf area, as plants strive to surpass competition, in some cases by elongation of the culm or stem to the detriment of leaf expansion 9,14,21. Thus, more competitive canola genotypes were supposed to maintain photosynthesis rates even under competition, compared to less competitive ones 23. Furthermore, the relationship between net assimilation rate and leaf area expansion is not always proportional.
The canola genotypes also suffered a relative decrease compared to the black oat average in the studied proportions, suggesting that the crop had greater competition ability than the weed. Maximum dry mass reductions due to competition reached 60-75% for all varieties.
The relative competition index (CR) of canola (Table 3, page 129) indicated that the crop was more competitive than black oats when it differed from ‘1’ by the one-way t-test 15.
Table 3: Tabla 3: Competitive parameters of canola (Brassica napus) hybrids under competition with black oat (Avena strigosa), in terms of relative competitiveness (CR), clustering coefficient (K), and aggressiveness (A) in experiments conducted by the substitutive series method. Parámetros competitivos de híbridos de canola (Brassica napus) en competencia con avena negra (Avena strigosa), en términos de competitividad relativa (CR), coeficiente de agrupamiento (K) y agresividad (A) en experimentos realizados por el método de serie sustitutiva.

CR: * = differs from “1” by the one-way t-test, at 5% probability; Kx Ky: * = Kx differs from Ky by the two-way t-test, at 5% probability; A : * = differs from “0” by the one-way t-test, at 5% probability.
CR: * = difiere de “1” en la prueba t unidireccional, con una probabilidad del 5%; Kx Ky: * = Kx difiere de Ky en la prueba t de dos vías, con un 5% probabilidad; A: * = difiere de “0” en la prueba t unidireccional, con un 5% de probabilidad.
This was reported for Hyola 76 and 571 for all variables except Ci, and for the leaf area of Hyola 433 (Table 3, page 129). This endorses Hyola 76 and 571 as more competitive with black oat than Hyola 61 and 433. The joint study of the values of CR, K (dominance), and C (competitiveness) represents the reliability between the studied competitiveness 6.
The CR index relates the comparison between the growth of species, relating CR(canola) to CR(black oat) Therefore, canola was considered more competitive than black oats when CR > 1 11 and vice versa. In a study with canola and turnip, Galon et al. (2015) showed that CR = 1.59 (leaf area), demonstrating a greater ability of the hybrid Hyola 433 in relation to the others tested.
The clustering coefficient (K) reports, in a loose definition, the ability of different plants of a given species to aggregate efforts aiming to surpass the competition imposed by plants of a different genetic background, including genotype, ecotype, biotype, or species, as discussed by Concenço (2016). K is superior for the crop (Kcrop) when it differs from Kcompetitor18, as found for Hyola 61 in AF and MS, Hyola 76 and 571 in A, E, AF, and MS, and Hyola 433 in Ci and AF (Table 3). This corroborates the CR values, where overall Hyola 76 and 571 performed better. Similar results were also observed by Durigon et al. (2019) when working with the canola hybrid Hyola 571 and turnip (Raphanus sativus). The greater index of dominance of a species indicates greater efficiency in the apprehension of resources of the environment, thus, greater growth and development in conditions of competition 2.
The aggressiveness (A) determines how intense the attack of the crop is against the competitor, or how fast it responds to competition Durigon et al. (2019). Not considering Ci, where there was no difference for any genotype, Hyola 571 was more aggressive than black oat for all variables. Hyola 61 surpassed the competitor for AF and DM, and Hyola 76 performed better for A, E and DM. Hyola 433 performed better than the competitor only for AF (Table 3). The hybrid Hyola 571 was highlighted as the most aggressive canola genotype among those evaluated.
Conclusions
The results found evidence of intense competition between canola and black oat, independent of plant proportion, and the competitive ability of canola was distinct among hybrids. The hybrid Hyola 571 performed better than the others, followed by Hyola 76. Hyola 433 was the most susceptible competition with black oat. As the competition between canola and black oat is intense, weed control should be effective and respect the critical period of weed interference in canola. The choice for most competitive hybrids, such as Hyola 571, can help in the task of keeping high canola grain yield levels in areas infested with black oat.

